
Comparative Plastid Genome and Phylogenomic Analyses of Potamogeton Species

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and DNA Extraction
2.2. DNA Sequencing, Assembly, and Annotation
2.3. Comparative Genomics, Divergence Hotspot and Repeat Analysis
2.4. Phylogenetic and Substitution Rate Analysis
3. Results
3.1. Genome Features of Potamogetonaceae Species
3.2. Repeat and Simple Sequence Repeat (SSR) Analysis
3.3. Divergence Regions in the Potamogetonaceae Plastomes
3.4. Comparative Analyses of the Plastomes among Alismatales
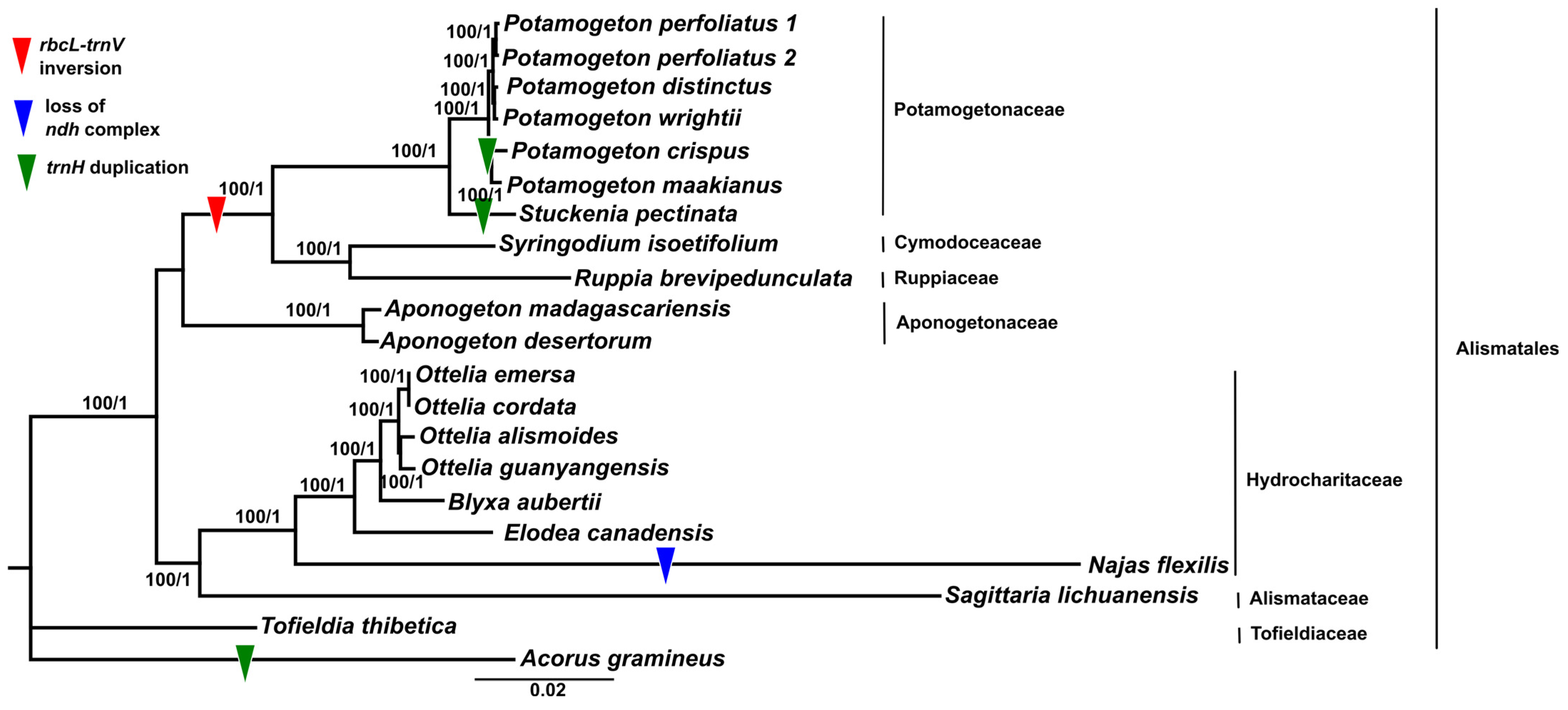
3.5. Phylogenetic Analysis
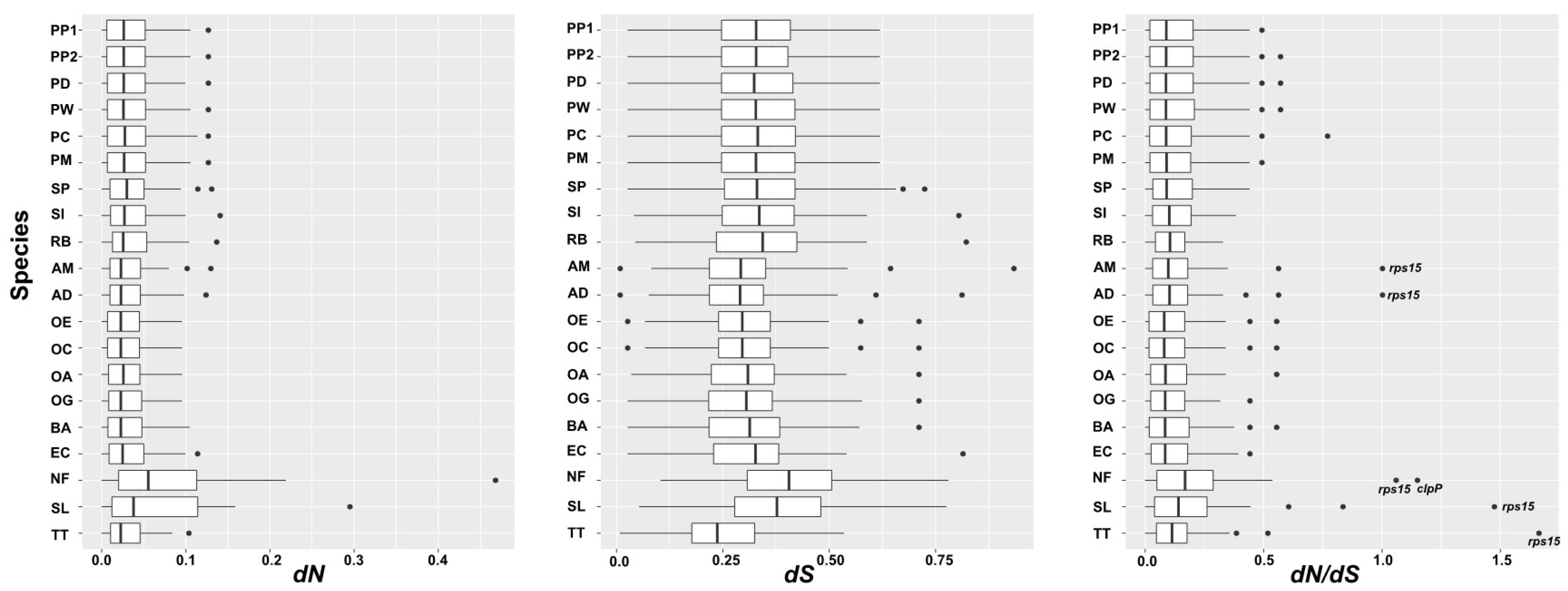
3.6. Nucleotide Substitution Rate Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Christenhusz, M.J.M.; Byng, J.W. The number of known plants species in the world and its annual increase. Phytotaxa 2016, 261, 201. [Google Scholar] [CrossRef]
- Hagström, J.O. Critical Researches on the Potamogeton; Kungliga Svenska Vetenskapsakademiens Handlinger; Almqvist & Wiksells: Stockholm, Sweden, 1916; Volume 55, pp. 1–281. [Google Scholar]
- Wiegleb, G. A redescription of Potamogeton distinctus including remarks on the taxonomy of the Potamogeton nodosus group. Plant Syst. Evol. 1990, 169, 245–259. [Google Scholar] [CrossRef]
- Wiegleb, G. The importance of stem anatomical characters for the systematics of the genus Potamogeton L. Flora 1990, 184, 197–208. [Google Scholar] [CrossRef]
- Xu, Z.; Chang, L. (Eds.) Potamogetonaceae. In Identification and Control of Common Weeds; Springer: Singapore, 2017; Volume 3, pp. 733–737. [Google Scholar]
- Les, D.H.; Haynes, R.R. Coleogeton (Potamogetonaceae), a new genus of Pondweeds. Novon 1996, 6, 389. [Google Scholar] [CrossRef]
- Holub, J. Stuckenia Börner 1912—The correct name for Coleogeton (Potamogetonaceae). Preslia 1997, 69, 361–366. [Google Scholar]
- Kaplan, Z. Phenotypic plasticity in Potamogeton (Potamogetonaceae). Folia Geobot. 2002, 37, 141–170. [Google Scholar] [CrossRef]
- Kaplan, Z.; Symoens, J.-J. Taxonomy, distribution and nomenclature of three confused broad-leaved Potamogeton species occurring in Africa and on surrounding islands. Bot. J. Linn. Soc. 2005, 148, 329–357. [Google Scholar] [CrossRef]
- Wafai, B.A.; Khuroo, A.A. Fruit morphology of the genus Potamogeton L. in Kashmir Himalaya and its utility in taxonomic delimitation. J. Asia-Pac. Biodivers. 2017, 10, 274–278. [Google Scholar]
- Wieglet, G.; Kaplan, Z. An account of the species of Potamogeton L. (Potamogetonaceae). Folia Geobot. 1998, 33, 241–316. [Google Scholar] [CrossRef]
- Kaplan, Z.; Fehrer, J. Erroneous identities of Potamogeton hybrids corrected by molecular analysis of plants from type clones. Taxon 2011, 60, 758–766. [Google Scholar] [CrossRef]
- Kaplan, Z.; Fehrer, J.; Hellquist, C. Potamogeton × jacobsii (Potamogetonaceae) from New South Wales, Australia—The first Potamogeton hybrid from the Southern Hemisphere. Telopea 2011, 13, 245–256. [Google Scholar] [CrossRef]
- Kaplan, Z.; Fehrer, J. Molecular identification of hybrids from a former hot spot of Potamogeton hybrid diversity. Aquat. Bot. 2013, 105, 34–40. [Google Scholar] [CrossRef]
- Kaplan, Z.; Fehrer, J.; Bambasová, V.; Hellquist, C.B. The endangered Florida pondweed (Potamogeton floridanus) is a hybrid: Why we need to understand biodiversity thoroughly. PLoS ONE 2018, 13, e0195241. [Google Scholar] [CrossRef] [PubMed]
- Lindqvist, C.; De Laet, J.; Haynes, R.R.; Aagesen, L.; Keener, B.R.; Albert, V.A. Molecular phylogenetics of an aquatic plant lineage, Potamogetonaceae. Cladistics 2006, 22, 568–588. [Google Scholar] [CrossRef] [PubMed]
- Iida, S.; Kosuge, K.; Kadono, Y. Molecular phylogeny of Japanese Potamogeton species in light of noncoding chloroplast sequences. Aquat. Bot. 2004, 80, 115–127. [Google Scholar] [CrossRef]
- Wang, Q.D.; Zhang, T.; Wang, J.B. Phylogenetic relationships and hybrid origin of Potamogeton species (Potamogetonaceae) distributed in China: Insights from the nuclear ribosomal internal transcribed spacer sequence (ITS). Plant Syst. Evol. 2007, 267, 65–78. [Google Scholar] [CrossRef]
- Du, Z.-Y.; Yang, C.-F.; Chen, J.-M.; Guo, Y.-H. Identification of hybrids in broad-leaved Potamogeton species (Potamogetonaceae) in China using nuclear and chloroplast DNA sequence data. Plant Syst. Evol. 2010, 287, 57–63. [Google Scholar] [CrossRef]
- Aykurt, C.; Fehrer, J.; Sarı Yol, D.; Kaplan, Z.; Bambasova, V.; Deniz, İ.G.; Aydemir, E.; İmir, N. Taxonomic treatment and phylogenetic analysis of the family Potamogetonaceae in Turkey. Taxon 2020, 69, 1172–1190. [Google Scholar] [CrossRef]
- Wicke, S.; Schneeweiss, G.M.; dePamphilis, C.W.; Müller, K.F.; Quandt, D. The evolution of the plastid chromosome in land plants: Gene content, gene order, gene function. Plant Mol. Biol. 2011, 76, 273–297. [Google Scholar] [CrossRef]
- Dobrogojski, J.; Adamiec, M.; Luciński, R. The chloroplast genome: A review. Acta Physiol Plant. 2020, 42, 98. [Google Scholar] [CrossRef]
- Du, Z.-Y.; Yang, C.-F.; Chen, J.-M.; Guo, Y.-H.; Kadiri, A.B. Using DNA-based techniques to identify hybrids among linear-leaved Potamogeton plants collected in China. J. Syst. Evol. 2010, 48, 265–270. [Google Scholar] [CrossRef]
- Li, S.; Duan, W.; Zhao, J.; Jing, Y.; Feng, M.; Kuang, B.; Wei, N.; Chen, B.; Yang, X. Comparative analysis of chloroplast genome in Saccharum spp. and related members of ‘Saccharum Complex’. Int. J. Mol. Sci. 2022, 23, 7661. [Google Scholar] [CrossRef] [PubMed]
- Jansen, R.K.; Ruhlman, T.A. Plastid Genomes of Seed Plants. In Genomics of Chloroplasts and Mitochondria; Bock, R., Knoop, V., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 103–126. [Google Scholar]
- Cai, Z.; Guisinger, M.; Kim, H.G.; Ruck, E.; Blazier, J.C.; McMurtry, V.; Kuehl, J.V.; Boore, J.; Jansen, R.K. Extensive reorganization of the plastid genome of Trifolium subterraneum (Fabaceae) is associated with numerous repeated sequences and novel DNA insertions. J. Mol. Evol. 2008, 67, 696–704. [Google Scholar] [CrossRef] [PubMed]
- Choi, I.-S.; Choi, B.-H. The distinct plastid genome structure of Maackia fauriei (Fabaceae: Papilionoideae) and its systematic implications for genistoids and tribe Sophoreae. PLoS ONE 2017, 12, e0173766. [Google Scholar] [CrossRef]
- Duan, L.; Li, S.J.; Su, C.; Sirichamorn, Y.; Han, L.N.; Ye, W.; Loc, P.K.; Wen, J.; Compton, J.A.; Schrire, B.; et al. Phylogenomic framework of the IRLC legumes (Leguminosae subfamily Papilionoideae) and intercontinental biogeography of tribe Wisterieae. Mol. Phylogenet. Evol. 2021, 163, 107235. [Google Scholar] [CrossRef]
- Wu, S.; Chen, J.; Li, Y.; Liu, A.; Li, A.; Yin, M.; Shrestha, N.; Liu, J.; Ren, G. Extensive genomic rearrangements mediated by repetitive sequences in plastomes of Medicago and its relatives. BMC Plant Biol. 2021, 21, 421. [Google Scholar] [CrossRef]
- Son, O.; Choi, K.S. Characterization of the chloroplast genome structure of Gueldenstaedtia verna (Papilionoideae) and comparative analyses among IRLC species. Forests 2022, 13, 1942. [Google Scholar] [CrossRef]
- Chumley, T.W.; Palmer, J.D.; Mower, J.P.; Fourcade, H.M.; Calie, P.J.; Boore, J.L.; Jansen, R.K. The complete chloroplast genome sequence of Pelargonium × hortorum: Organization and evolution of the largest and most highly rearranged chloroplast genome of land plants. Mol. Biol. Evol. 2006, 23, 2175–2190. [Google Scholar] [CrossRef]
- Blazier, C.J.; Guisinger, M.M.; Jansen, R.K. Recent loss of plastid-encoded ndh genes within Erodium (Geraniaceae). Plant Mol. Biol. 2011, 76, 263–272. [Google Scholar] [CrossRef]
- Weng, M.-L.; Blazier, J.C.; Govindu, M.; Jansen, R.K. Reconstruction of the ancestral plastid genome in Geraniaceae reveals a correlation between genome rearrangements, repeats, and nucleotide substitution rates. Mol. Biol. Evol. 2014, 31, 645–659. [Google Scholar] [CrossRef]
- Cosner, M.E.; Jansen, R.K.; Palmer, J.D.; Downie, S.R. The highly rearranged chloroplast genome of Trachelium caeruleum (Campanulaceae): Multiple inversions, inverted repeat expansion and contraction, transposition, insertions/deletions, and several repeat families. Curr. Genet. 1997, 31, 419–429. [Google Scholar] [CrossRef] [PubMed]
- Cosner, M.E.; Raubeson, L.A.; Jansen, R.K. Chloroplast DNA rearrangements in Campanulaceae: Phylogenetic utility of highly rearranged genomes. BMC Evol. Biol. 2004, 4, 27. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-A.; Cheon, K.-S. Complete chloroplast genome sequence of Adenophora racemosa (Campanulaceae): Comparative analysis with congeneric species. PLoS ONE 2021, 16, e0248788. [Google Scholar] [CrossRef]
- Wicke, S. Genomic Evolution in Orobanchaceae. In Parasitic Orobanchaceae; Joel, D.M., Gressel, J., Musselman, L.J., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 267–286. [Google Scholar]
- Cusimano, N.; Wicke, S. Massive intracellular gene transfer during plastid genome reduction in nongreen Orobanchaceae. New Phytol. 2016, 210, 680–693. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.S.; Park, S. Complete plastid and mitochondrial genomes of Aeginetia indica reveal intracellular gene transfer (IGT), horizontal gene transfer (HGT), and cytoplasmic male sterility (CMS). Int. J. Mol. Sci. 2021, 22, 6143. [Google Scholar] [CrossRef]
- Davis, C.C.; Xi, Z.; Mathews, S. Plastid phylogenomics and green plant phylogeny: Almost full circle but not quite there. BMC Biol. 2014, 12, 11. [Google Scholar] [CrossRef]
- Henriquez, C.L.; Arias, T.; Pires, J.C.; Croat, T.B.; Schaal, B.A. Phylogenomics of the plant family Araceae. Mol. Phylogenet. Evol. 2014, 75, 91–102. [Google Scholar] [CrossRef]
- Li, H.T.; Luo, Y.; Gan, L.; Ma, P.F.; Gao, L.M.; Yang, J.B.; Cai, J.; Gitzendanner, M.A.; Fritsch, P.W.; Zhang, T.; et al. Plastid phylogenomic insights into relationships of all flowering plant families. BMC Biol. 2021, 19, 232. [Google Scholar] [CrossRef]
- Wu, F.H.; Chan, M.T.; Liao, D.C.; Hsu, C.T.; Lee, Y.W.; Daniell, H.; Duvall, M.R.; Lin, C.S. Complete chloroplast genome of Oncidium Gower Ramsey and evaluation of molecular markers for identification and breeding in Oncidiinae. BMC Plant Biol. 2010, 10, 68. [Google Scholar] [CrossRef]
- Li, C.; Zheng, Y.; Huang, P. Molecular markers from the chloroplast genome of rose provide a complementary tool for variety discrimination and profiling. Sci. Rep. 2020, 10, 12188. [Google Scholar] [CrossRef]
- Li, B.; Lin, F.; Huang, P.; Guo, W.; Zheng, Y. Development of nuclear SSR and chloroplast genome markers in diverse Liriodendron chinense germplasm based on low-coverage whole genome sequencing. Biol. Res. 2020, 53, 21. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, E.N.; Ruhlman, T.A.; Weng, M.L.; Khiyami, M.A.; Sabir, J.S.; Hajarah, N.H.; Alharbi, N.S.; Rabah, S.O.; Jansen, R.K. Plastome-wide nucleotide substitution rates reveal accelerated rates in Papilionoideae and correlations with genome features across Legume subfamilies. J. Mol. Evol. 2017, 84, 187–203. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.S.; Ha, Y.-H.; Gil, H.-Y.; Choi, K.; Kim, D.-K.; Oh, S.-H. Two Korean endemic Clematis chloroplast genomes: Inversion, reposition, expansion of the inverted repeat region, phylogenetic analysis, and nucleotide substitution rates. Plants 2021, 10, 397. [Google Scholar] [CrossRef] [PubMed]
- Claude, S.-J.; Park, S.; Park, S. Gene loss, genome rearrangement, and accelerated substitution rates in plastid genome of Hypericum ascyron (Hypericaceae). BMC Plant Biol. 2022, 22, 135. [Google Scholar] [CrossRef]
- Luo, Y.; Ma, P.-F.; Li, H.-T.; Yang, J.-B.; Wang, H.; Li, D.-Z. Plastid phylogenomic analyses resolve Tofieldiaceae as the root of the early diverging monocot order Alismatales. Genome Biol Evol. 2016, 8, 932–945. [Google Scholar] [CrossRef]
- Jin, J.J.; Yu, W.B.; Yang, J.B.; Song, Y.; DePamphilis, C.W.; Yi, T.S.; Li, D.Z. GetOrganelle: A fast and versatile toolkit for accurate de novo assembly of organelle genomes. Genome Biol. 2020, 21, 241. [Google Scholar] [CrossRef]
- Tillich, M.; Lehwark, P.; Pellizzer, T.; Ulbricht-Jones, E.S.; Fischer, A.; Bock, R.; Greiner, S. GeSeq—Versatile and accurate annotation of organelle genomes. Nucleic Acids Res. 2017, 45, W6–W11. [Google Scholar] [CrossRef]
- Chan, P.P.; Lin, B.Y.; Mak, A.J.; Lowe, T.M. tRNAscan-SE 2.0: Improved detection and functional classification of transfer RNA genes. Nucleic Acids Res. 2021, 49, 9077–9096. [Google Scholar] [CrossRef]
- Greiner, S.; Lehwark, P.; Bock, R. OrganellarGenomeDRAW (OGDRAW) version 1.3.1: Expanded toolkit for the graphical visualization of organellar genomes. Nucleic Acids Res. 2019, 47, W59–W64. [Google Scholar] [CrossRef]
- Katoh, K. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Kurtz, S.; Choudhuri, J.V.; Ohlebusch, E.; Schleiermacher, C.; Stoye, J.; Giegerich, R. REPuter: The manifold applications of repeat analysis on a genomic scale. Nucleic Acids Res. 2001, 29, 4633–4642. [Google Scholar] [CrossRef]
- Beier, S.; Thiel, T.; Münch, T.; Scholz, U.; Mascher, M. MISA-web: A web server for microsatellite prediction. Bioinformatics 2017, 33, 2583–2585. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Yang, Z. PAML 4: Phylogenetic Analysis by Maximum Likelihood. Mol. Biol. Evol. 2007, 24, 1586–1591. [Google Scholar] [CrossRef]
- Tian, J.-J.; Zhang, X.; Liu, Z.-D.; Shen, S.-K. Characterization of the complete chloroplast genome sequence of submerged macrophyte Stuckenia pectinata (Potamogetonaceae) and its phylogenetic position. Mitochondrial DNA Part B 2020, 5, 327–328. [Google Scholar] [CrossRef]
- Rogalski, M.; Ruf, S.; Bock, R. Tobacco plastid ribosomal protein S18 is essential for cell survival. Nucleic Acids Res. 2006, 34, 4537–4545. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.-S.; Lin, C.-P.; Hsu, C.-Y.; Wang, R.-J.; Chaw, S.-M. Comparative chloroplast genomes of Pinaceae: Insights into the mechanism of diversified genomic organizations. Genome Biol. Evol. 2011, 3, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Ruhlman, T.A.; Zhang, J.; Blazier, J.C.; Sabir, J.S.M.; Jansen, R.K. Recombination-dependent replication and gene conversion homogenize repeat sequences and diversify plastid genome structure. Am. J. Bot. 2017, 104, 559–572. [Google Scholar] [CrossRef]
- Fullerton, S.M.; Bernardo Carvalho, A.; Clark, A.G. Local rates of recombination are positively correlated with GC content in the human genome. Mol. Biol. Evol. 2001, 18, 1139–1142. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.H.; Wicke, S.; Wang, H.; Jin, J.J.; Chen, S.Y.; Zhang, S.D.; Li, D.Z.; Yi, T.S. Plastid genome evolution in the early-diverging Legume subfamily Cercidoideae (Fabaceae). Front. Plant Sci. 2018, 9, 138. [Google Scholar] [CrossRef] [PubMed]
- Cauz-Santos, L.A.; da Costa, Z.P.; Callot, C.; Cauet, S.; Zucchi, M.I.; Bergès, H.; van den Berg, C.; Vieira, M.L.C. A repertory of rearrangements and the loss of an inverted repeat region in Passiflora chloroplast genomes. Genome Biol. Evol. 2020, 12, 1841–1857. [Google Scholar] [CrossRef]
- Chen, N.; Sha, L.N.; Wang, Y.L.; Yin, L.J.; Zhang, Y.; Wang, Y.; Wu, D.D.; Kang, H.Y.; Zhang, H.Q.; Zhou, Y.H.; et al. Variation in plastome sizes accompanied by evolutionary history in monogenomic Triticeae (Poaceae: Triticeae). Front. Plant Sci. 2021, 12, 741063. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.S.; Son, O.; Park, S. The chloroplast genome of Elaeagnus macrophylla and trnH duplication event in Elaeagnaceae. PLoS ONE 2015, 10, e0138727. [Google Scholar] [CrossRef]
- Wang, R.-J.; Cheng, C.-L.; Chang, C.-C.; Wu, C.-L.; Su, T.-M.; Chaw, S.-M. Dynamics and evolution of the inverted repeat-large single copy junctions in the chloroplast genomes of monocots. BMC Evol. Biol. 2008, 8, 36. [Google Scholar] [CrossRef] [PubMed]
- Ross, T.G.; Barrett, C.F.; Soto Gomez, M.; Lam, V.K.; Henriquez, C.L.; Les, D.H.; Davis, J.I.; Cuenca, A.; Petersen, G.; Seberg, O.; et al. Plastid phylogenomics and molecular evolution of Alismatales. Cladistics 2016, 32, 160–178. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.Y.; Lu, B.; Morales-Briones, D.F.; Moody, M.L.; Liu, F.; Hu, G.W.; Huang, C.H.; Chen, J.M.; Wang, Q.F. Phylogenomic analyses of Alismatales shed light into adaptations to aquatic environments. Mol. Biol. Evol. 2022, 39, msac079. [Google Scholar] [CrossRef]
- Haynes, R.R.; Les, D.H.; Král, M.; Kral, M. Two New Combinations in Stuckenia, the correct Name for Coleogeton (Potamogetonaceae). Novon 1998, 8, 241. [Google Scholar] [CrossRef]
- Ito, Y.; Robledo, G.L.; Iharlegui, L.; Tanaka, N. Phylogeny of Potamogeton (Potamogetonaceae) revisited: Implications for hybridization and Introgression in Argentina. Bull. Natl. Sci. Mus. 2016, 42, 131–141. [Google Scholar]
- Clegg, M.T.; Gaut, B.S.; Learn, G.H.; Morton, B.R. Rates and patterns of chloroplast DNA evolution. Proc. Natl. Acad. Sci. USA 1994, 91, 6795–6801. [Google Scholar] [CrossRef] [PubMed]
- Lawrie, D.S.; Messer, P.W.; Hershberg, R.; Petrov, D.A. Strong purifying selection at synonymous sites in D. melanogaster. PLoS Genet. 2013, 9, e1003527. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.-Y.; Yang, J.-X.; Bai, M.-Z.; Zhang, G.-Q.; Liu, Z.-J. The chloroplast genome evolution of Venus slipper (Paphiopedilum): IR expansion, SSC contraction, and highly rearranged SSC regions. BMC Plant Biol. 2021, 21, 248. [Google Scholar] [CrossRef] [PubMed]








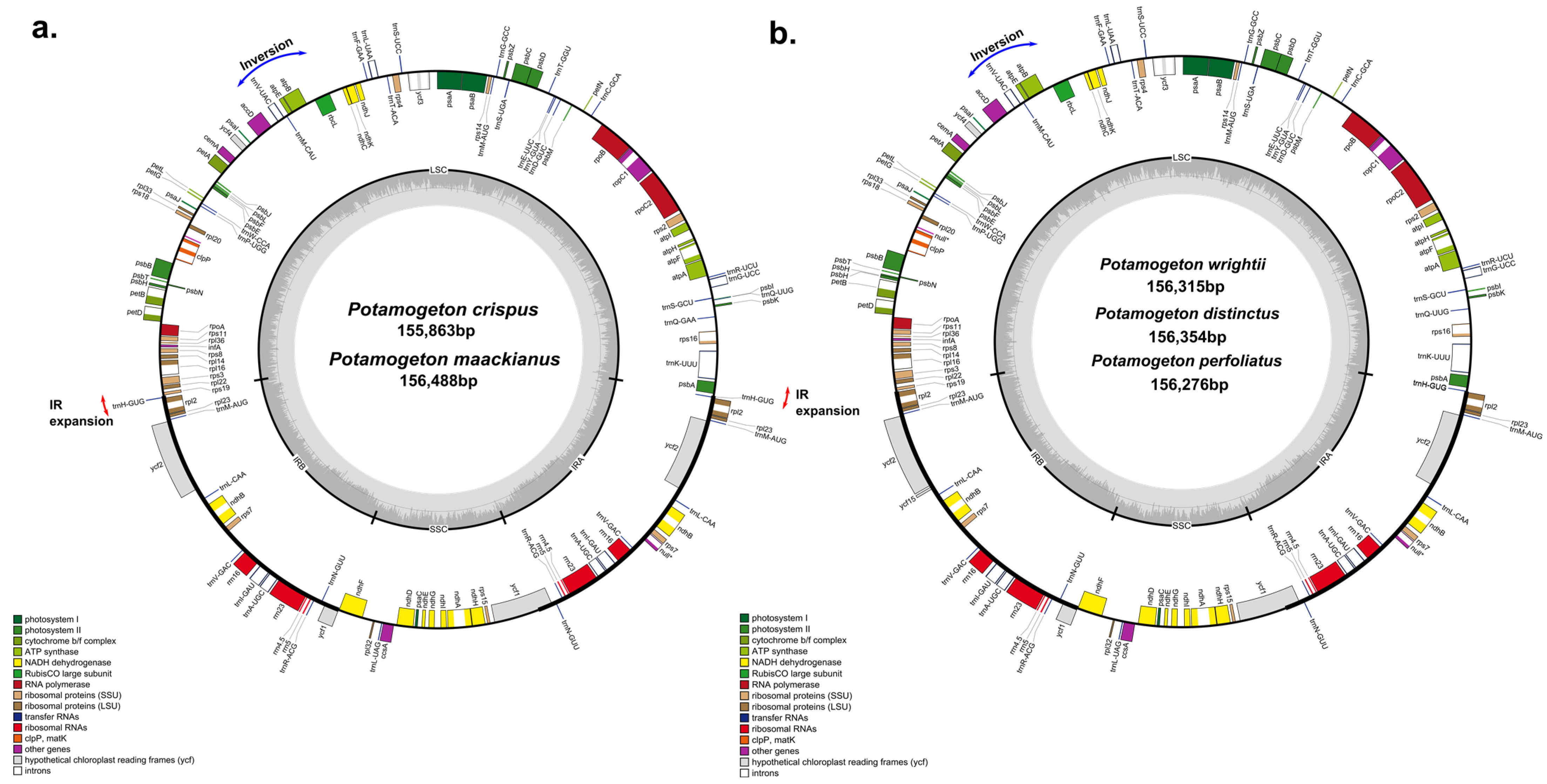{kind=link}
{kind=link}
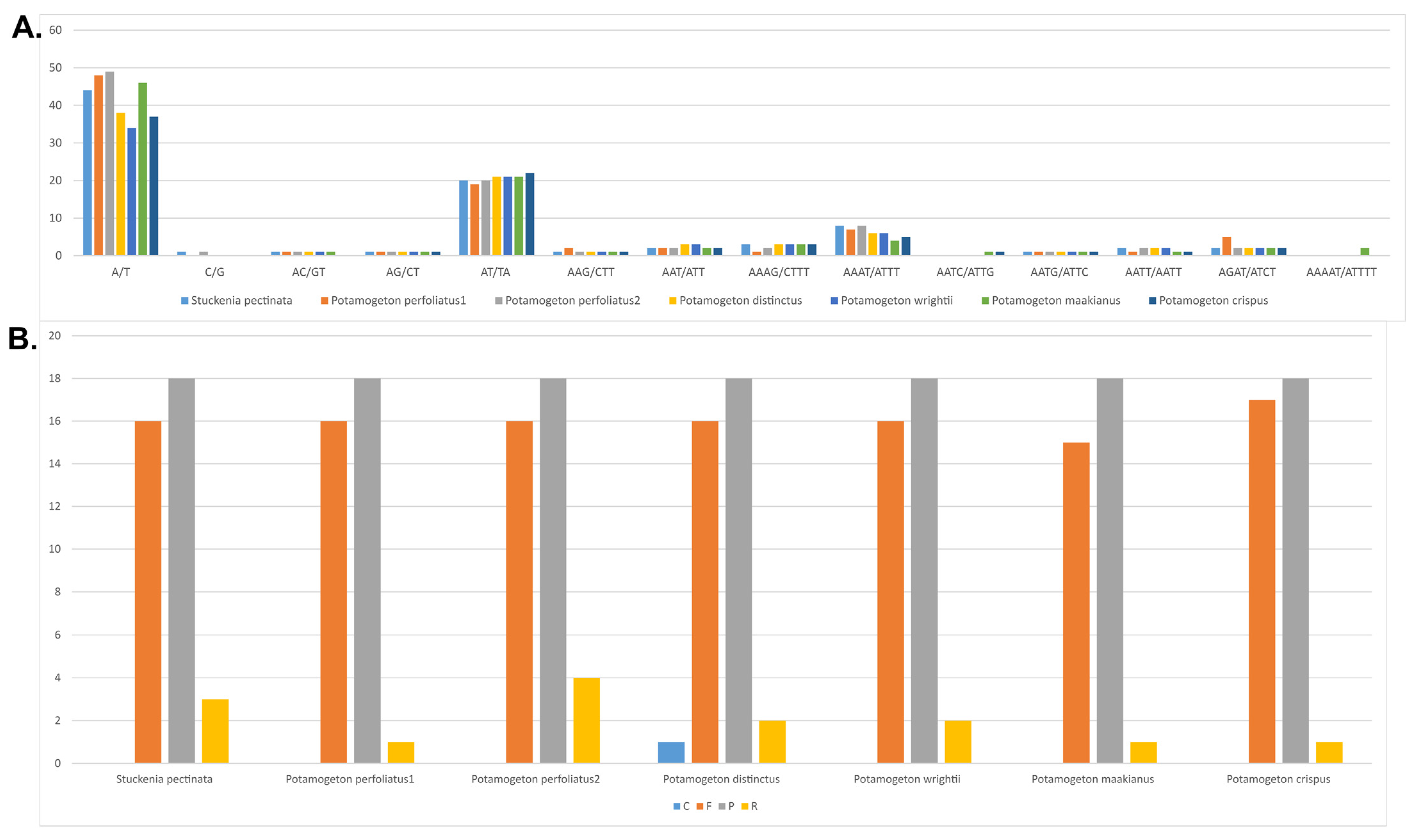{kind=link}
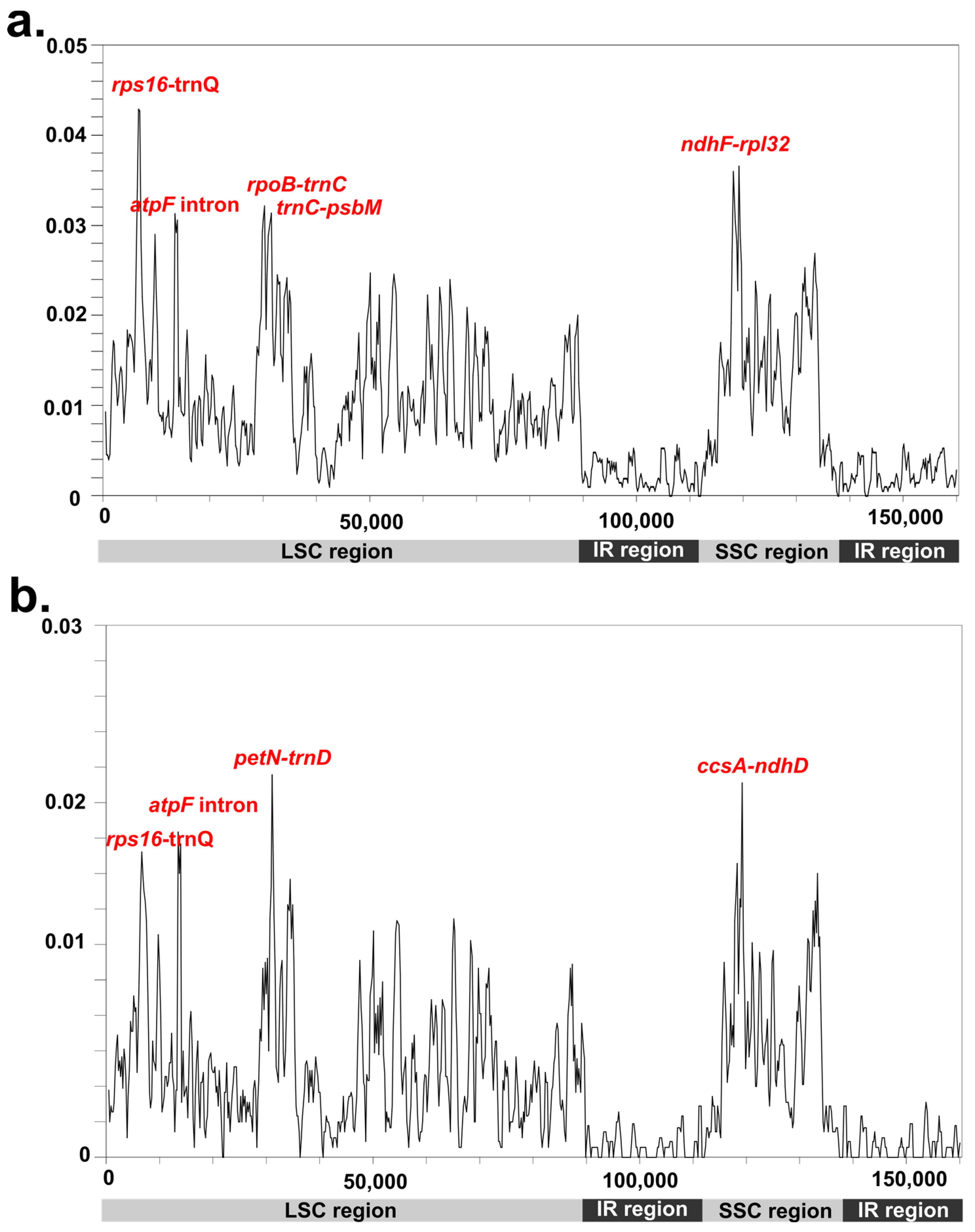{kind=link}
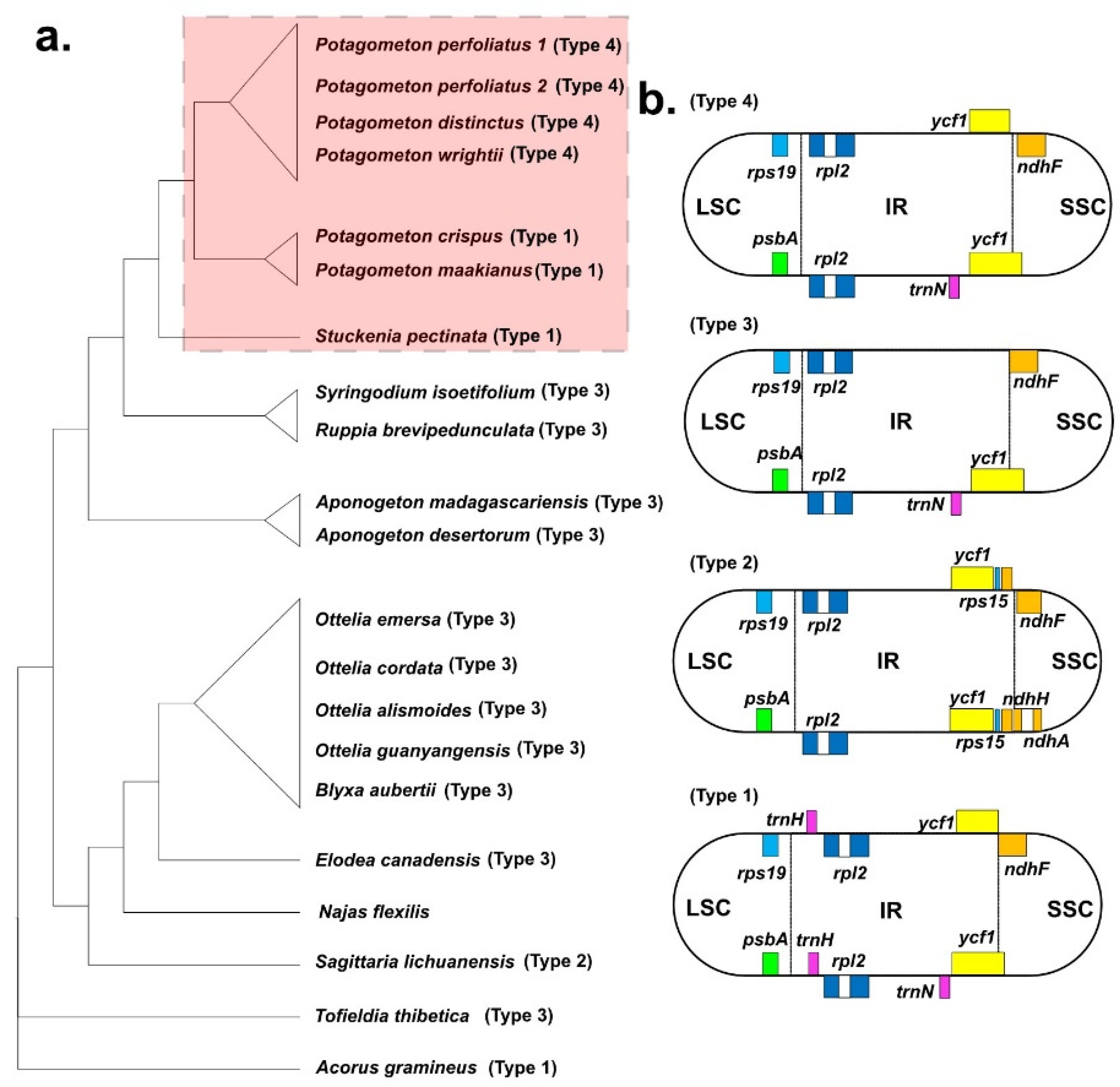{kind=link}
{kind=link}
{kind=link}
| Order | Family | Species | Length (bp) | GC Contents | NCBI Accession Number | |||
|---|---|---|---|---|---|---|---|---|
| Total | LSC | SSC | IR | |||||
| Acorales | Acoraceae | Acorus gramineus | 152,849 | 82,977 | 18,228 | 25,822 | 38.7% | NC_026299 |
| Alismatales | Tofieldiaceae | Tofieldia thibetica | 155,512 | 84,584 | 18,151 | 26,388 | 37.4% | NC_029813 |
| Alismataceae | Sagittaria lichuanensis | 179,007 | 99,125 | 13,278 | 33,302 | 36.8% | NC_029815 | |
| Hydrocharitaceae | Najas flexilis | 156,366 | 88,697 | 15,266 | 31,201 | 38.2% | NC_021936 | |
| Eldea canadensis | 156,700 | 86,194 | 17,808 | 26,349 | 37.0% | NC_018541 | ||
| Blyxa aubertii | 158,187 | 87,799 | 18,804 | 25,792 | 36.5% | MK940507 | ||
| Ottelia guanyangensis | 157,362 | 87,230 | 19,004 | 25,564 | 36.7% | MK940522 | ||
| Ottelia alismoides | 157,880 | 87,699 | 19,067 | 25,557 | 36.6% | MK940517 | ||
| Ottelia cordata | 157,896 | 87,665 | 19,121 | 25,555 | 36.6% | MK940519 | ||
| Ottelia emersa | 157,896 | 87,665 | 19,121 | 25,555 | 36.6% | MK940520 | ||
| Aponogetonaceae | Aponogeton desertorum | 154,516 | 85,760 | 19,890 | 24,433 | 36.9% | MK570533 | |
| Aponogeton madagascariensis | 155,669 | 86,896 | 19,869 | 24,452 | 36.9% | MK570534 | ||
| Cymodoceaceae | Ruppia brevipedunculata | 158,943 | 88,857 | 19,130 | 25,478 | 35.8% | NC_051974 | |
| Syringodium isoetifolium | 159,333 | 89,055 | 19,160 | 25,559 | 35.9% | MZ325253 | ||
| Potamogetoncaceae | Stuckenia pectinata | 156,669 | 86,285 | 18,237 | 26,073 | 36.5% | MN661144 | |
| Potamogeton perfoliatus1 | 156,226 | 86,764 | 18,238 | 25,612 | 36.5% | NC_029814 | ||
| Potamogeton perfoliatus2 | 156,276 | 86,821 | 18,231 | 25,612 | 36.5% | This study (OQ561452) | ||
| Potamogeton maackianus | 156,488 | 86,833 | 18,221 | 25,717 | 36.5% | This study (OQ561451) | ||
| Potamogeton crispus | 155,863 | 86,191 | 18,182 | 25,745 | 36.5% | This study (OQ561449) | ||
| Potamogeton wrightii | 156,315 | 86,827 | 18,282 | 25,603 | 36.5% | This study (OQ561453) | ||
| Potamogeton distinctus | 156,354 | 86,898 | 18,286 | 25,585 | 36.5% | This study (OQ561450) | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, K.; Hwang, Y.; Hong, J.-K.; Kang, J.-S. Comparative Plastid Genome and Phylogenomic Analyses of Potamogeton Species. Genes 2023, 14, 1914. https://doi.org/10.3390/genes14101914
Choi K, Hwang Y, Hong J-K, Kang J-S. Comparative Plastid Genome and Phylogenomic Analyses of Potamogeton Species. Genes. 2023; 14(10):1914. https://doi.org/10.3390/genes14101914
Chicago/Turabian StyleChoi, KyoungSu, Yong Hwang, Jeong-Ki Hong, and Jong-Soo Kang. 2023. "Comparative Plastid Genome and Phylogenomic Analyses of Potamogeton Species" Genes 14, no. 10: 1914. https://doi.org/10.3390/genes14101914
APA StyleChoi, K., Hwang, Y., Hong, J. -K., & Kang, J. -S. (2023). Comparative Plastid Genome and Phylogenomic Analyses of Potamogeton Species. Genes, 14(10), 1914. https://doi.org/10.3390/genes14101914