

Physiological, Photosynthetic, and Transcriptomics Insights into the Influence of Shading on Leafy Sweet Potato
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Measurement of Photosynthesis
2.3. Measurement of Chlorophyll Contents
2.4. Measurement of Soluble Sugar, Sucrose, and Starch Contents
2.5. Total Phenolics and Flavonoids Determination
2.6. Chlorogenic Acid Determination
2.7. Transcriptome Analysis
2.8. qRT-PCR
2.9. Statistical Analysis
3. Results
3.1. Effect of Shading on Yield of Leafy Sweet Potato
3.2. Effect of Shading on Chlorophyll Concentration and Pn of Leafy Sweet Potato
3.3. Effect of Shading on Soluble Sugar, Sucrose, and Starch of Leafy Sweet Potato
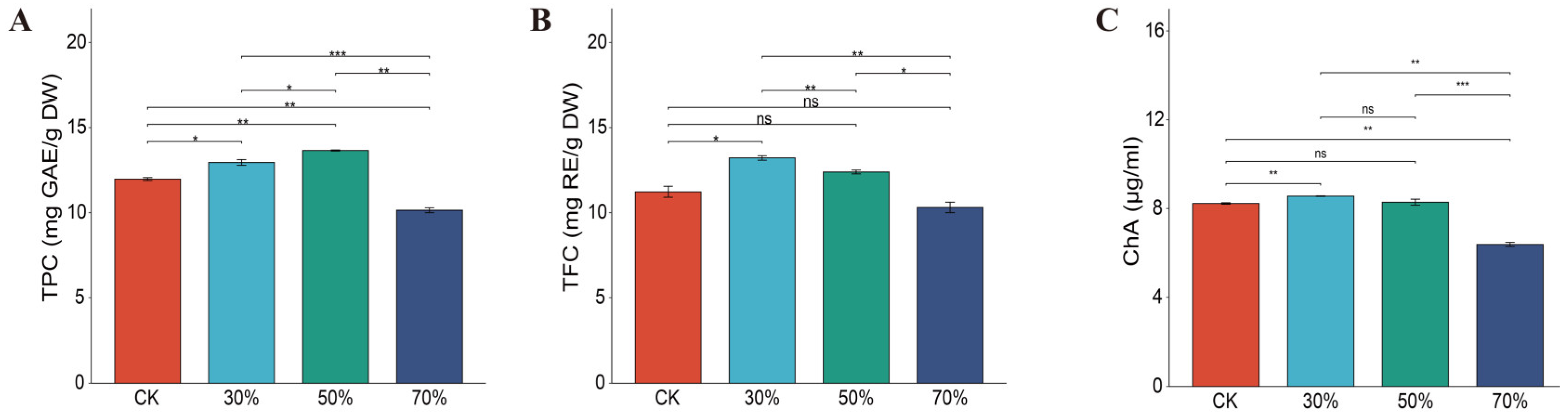
3.4. Effect of Shading on TPC, TFC, and Chlorogenic Acid in Leafy Sweet Potato
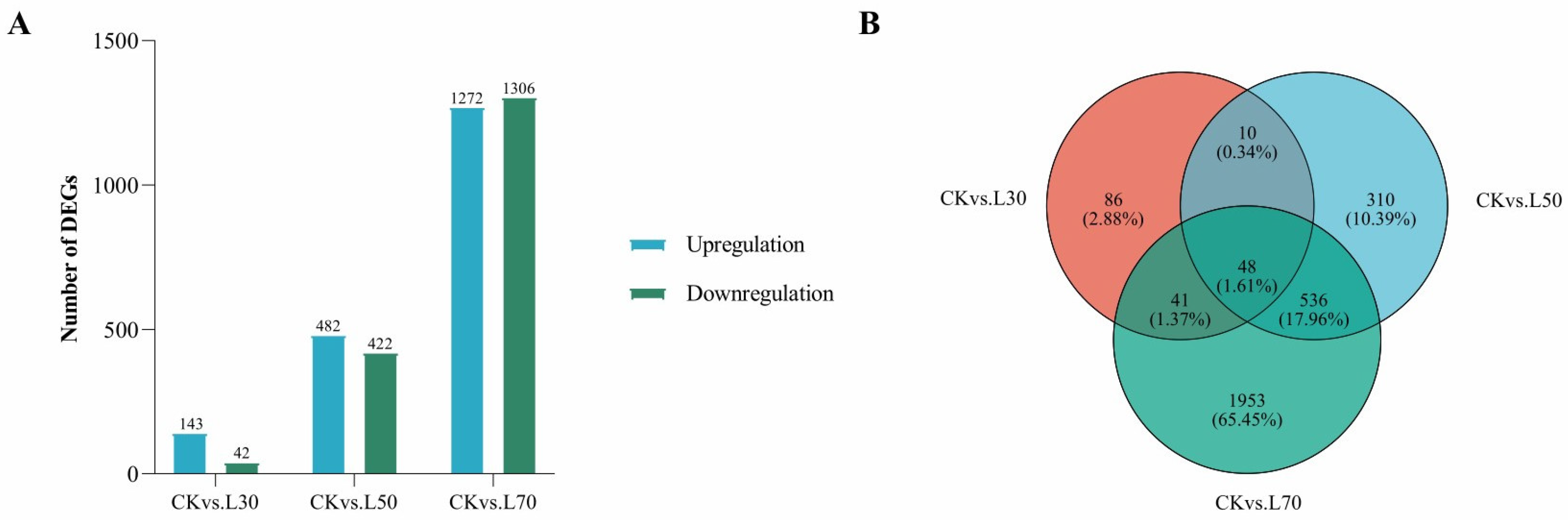
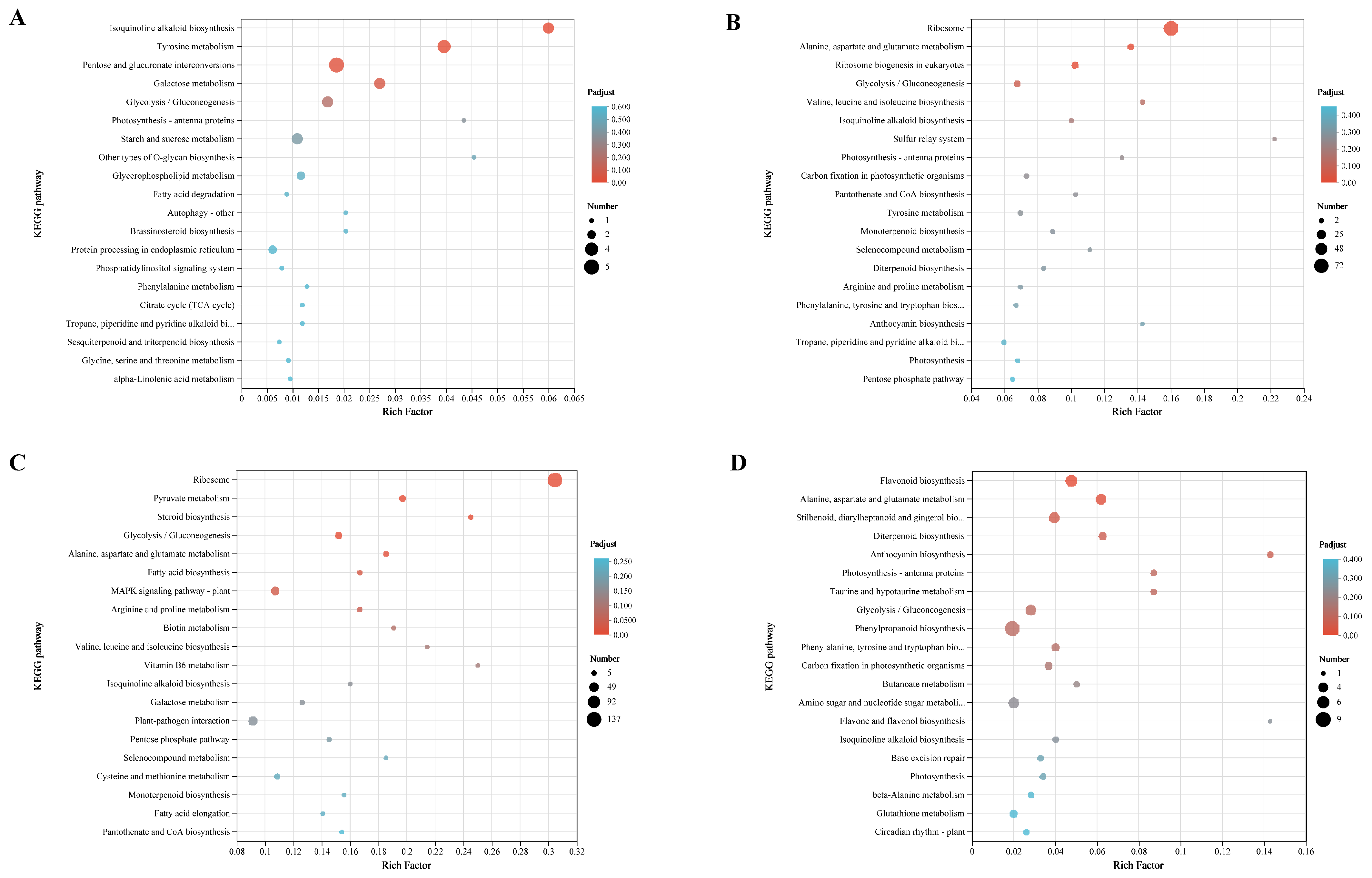
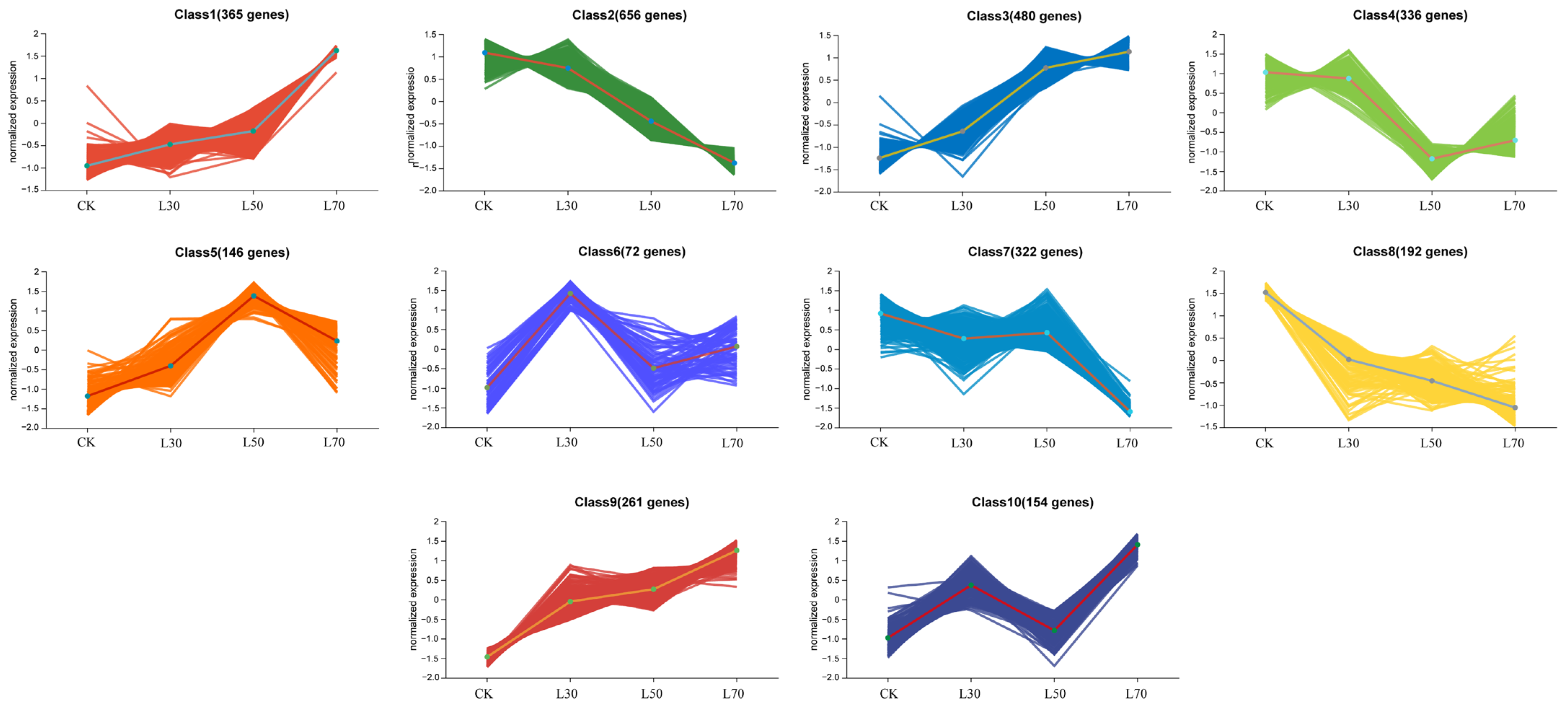
3.5. Transcriptome Analysis of Leaves under Different Shading Treatments
4. Discussion
4.1. Impact of Different Shading Treatments on Photosynthetic Characteristics and Carbohydrates Contents
4.2. Differentiation of Transcription Levels under Different Shading Conditions
4.3. Photosynthesis-Related DEGs in Eshu 10 Play an Important Role during Shading Conditions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Alam, M.K. A comprehensive review of sweet potato (Ipomoea batatas [L.] Lam): Revisiting the associated health benefits. Trends Food Sci. Technol. 2021, 115, 512–529. [Google Scholar] [CrossRef]
- Drapal, M.; Rossel, G.; Heider, B.; Fraser, P.D. Metabolic diversity in sweet potato (Ipomoea batatas, Lam.) leaves and storage roots. Hortic. Res. 2019, 6, 2. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.C.; Chen, C.C.; Lin, K.H.; Chao, P.Y.; Lin, H.H.; Huang, M.Y. Bioactive compounds, antioxidants, and health benefits of sweet potato leaves. Molecules 2021, 26, 1820. [Google Scholar] [CrossRef] [PubMed]
- Islam, S. Sweetpotato (Ipomoea batatas L.) leaf: Its potential effect on human health and nutrition. J. Food Sci. 2006, 71, 13–21. [Google Scholar] [CrossRef]
- Ishida, H.; Suzuno, H.; Sugiyama, N.; Innami, S.; Tadokoro, T.; Maekawa, A. Nutritive evaluation on chemical components of leaves, stalks and stems of sweet potatoes (Ipomoea batatas poir). Food Chem. 2000, 68, 359–367. [Google Scholar] [CrossRef]
- Sun, H.; Mu, T.; Xi, L.; Zhang, M.; Chen, J. Sweet potato (Ipomoea batatas L.) leaves as nutritional and functional foods. Food Chem. 2014, 156, 380–389. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.C.; Ameen, A.; Fang, B.P.; Liao, M.H.; Chen, J.Y.; Huang, L.F.; Zou, H.D.; Wang, Z.Y. Nutritional composition and health benefits of leaf-vegetable sweet potato in South China. J. Food Compos. Anal. 2021, 96, 103714. [Google Scholar] [CrossRef]
- Krochmal-Marczak, B.; Cebulak, T.; Kapusta, I.; Oszmiański, J.; Kaszuba, J.; Żurek, N. The content of phenolic acids and flavonols in the leaves of nine varieties of sweet potatoes (Ipomoea batatas L.) depending on their development, grown in central Europe. Molecules 2020, 25, 3473. [Google Scholar] [CrossRef]
- Truong, V.D.; McFeeters, R.F.; Thompson, R.T.; Dean, L.L.; Shofran, B. Phenolic acid content and composition in leaves and roots of common commercial sweetpotato (Ipomea batatas L.) cultivars in the United States. J. Food Sci. 2007, 72, C343–C349. [Google Scholar] [CrossRef]
- Makori, S.I.; Mu, T.H.; Sun, H.N. Total polyphenol content, antioxidant activity, and individual phenolic composition of different edible parts of 4 sweet potato cultivars. Nat. Prod. Commun. 2020, 15, 1934578X20936931. [Google Scholar] [CrossRef]
- Wang, S.; Nie, S.; Zhu, F. Chemical constituents and health effects of sweet potato. Food Res. Int. 2016, 89, 90–116. [Google Scholar] [CrossRef] [PubMed]
- Karna, P.; Gundala, S.R.; Gupta, M.V.; Shamsi, S.A.; Pace, R.D.; Yates, C.; Narayan, S.; Aneja, R. Polyphenol-rich sweet potato greens extract inhibits proliferation and induces apoptosis in prostate cancer cells in vitro and in vivo. Carcinogenesis 2011, 32, 1872–1880. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.L.; Lee, S.L.; Chen, C.J.; Chen, H.C.; Kao, M.C.; Liu, C.H.; Chen, J.Y.; Lai, Y.T.; Wu, Y.C. Characterization of secondary metabolites from purple Ipomoea batatas leaves and their effects on glucose uptake. Molecules 2016, 21, 745. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Qin, L. Growth and photosynthetic characteristics of sweet potato (Ipomoea batatas) leaves grown under natural sunlight with supplemental LED lighting in a tropical greenhouse. J. Plant Physiol. 2020, 252, 153239. [Google Scholar] [CrossRef] [PubMed]
- Lyu, C.W.; Tang, D.; Zhang, K.; Wang, J. Influence of ridges for planting sweet potato on symbiotic ecological factors, photosynthetic abilities and population yield in relay intercropping system. Not. Bot. Horti Agrobot. 2020, 48, 849–861. [Google Scholar] [CrossRef]
- Song, D.; Tariq, A.; Pan, K.; Khan, S.U.; Saleh, T.A.; Gong, S.; Zhang, A.; Wu, X. Influence of planting distance and density on the yield and photosynthetic traits of sweet potato (Ipomoea balatas L.) under an intercropping system with walnut (Juglans regia) saplings. Soil Till. Res. 2020, 196, 104484. [Google Scholar] [CrossRef]
- Xiao, X.; Han, L.; Chen, H.; Wang, J.; Zhang, Y.; Hu, A. Intercropping enhances microbial community diversity and ecosystem functioning in maize fields. Front. Microbiol. 2023, 13, 1084452. [Google Scholar] [CrossRef] [PubMed]
- Wei, L. Study on the Growth and Development Characteristics of Sweet Potato in the Agriculture and Light Complementary System. Master’s Thesis, Zhejiang University, Hangzhou, China, 2017. [Google Scholar]
- He, Y.; Zhu, D.; Sun, Y.; Wang, Q.; Zhu, L.; Zeng, H. Metabonomic Profiling Analyses Reveal ANS Upregulation to Enhance the Flavonoid Pathway of Purple-Fleshed Sweet Potato Storage Root in Response to Deep Shading. Agronomy 2021, 11, 737. [Google Scholar] [CrossRef]
- Li, J.; Terzaghi, W.; Deng, X.W. Genomic basis for light control of plant development. Protein Cell 2012, 3, 106–116. [Google Scholar] [CrossRef]
- Park, W.; Chung, M.N.; Nam, S.S.; Kim, T.H.; Lee, H.W.; Goh, S.; Lee, I.B.; Shin, W.C. Evaluation of the Growth and Yield of Sweetpotato (Ipomoea Batatas L.) at Different Growth Stages under Low Light Intensity. Korean J. Crop Sci. 2021, 66, 146–154. [Google Scholar]
- Wei, L.; Yu, M.; Qin, N.; Huang, C.; Xie, Y.; Sun, W.; Wu, L.; Wang, W.; Wang, G. Effects of agro-photovoltaic integrating system on field illumination and sweet potato growth. J. Zhejiang Univ. (Agric. Life Sci.) 2019, 45, 288–295. [Google Scholar]
- Zhang, S.; Zuo, L.; Zhang, J.; Chen, P.; Wang, J.; Yang, M. Transcriptome analysis of Ulmus pumila ‘Jinye’ responses to different shading involved in chlorophyll metabolism. Tree Genet. Genomes 2017, 13, 64. [Google Scholar] [CrossRef]
- Ma, Z.; Yang, S.; Mao, J.; Li, W.; Li, W.; Zuo, C.; Chu, M.; Zhao, X.; Zhou, Q.; Chen, B. Effects of Shading on the Synthesis of Volatile Organic Compounds in ‘Marselan’ Grape Berries (Vitis vinifera L.). J. Plant Growth Regul. 2020, 40, 679–693. [Google Scholar] [CrossRef]
- Han, S.; Zhang, Q.; Wang, H.; Pei, D. Comparison of the transcriptomic responses of two Chrysanthemum morifolium cultivars to low light. Mol. Biol. Rep. 2021, 48, 7293–7301. [Google Scholar] [CrossRef] [PubMed]
- Zaman, S.; Shen, J.; Wang, S.; Song, D.; Wang, H.; Ding, S.; Pang, X.; Wang, M.; Sabir, I.A.; Wang, Y.; et al. Effect of shading on physiological attributes and comparative transcriptome analysis of Camellia sinensis cultivar reveals tolerance mechanisms to low temperatures. Front. Plant Sci. 2023, 14, 1114988. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Mu, T.; Xi, L.; Song, Z. Effects of domestic cooking methods on polyphenols and antioxidant activity of sweet potato leaves. J. Agric. Food Chem. 2014, 62, 8982–8989. [Google Scholar] [CrossRef]
- Sun, H.N.; Mu, T.H.; Xi, L.S. Effect of pH, heat, and light treatments on the antioxidant activity of sweet potato leaf polyphenols. Int. J. Food Prop. 2017, 20, 318–332. [Google Scholar] [CrossRef]
- Jia, Z.S.; Tang, M.; Wu, J. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar]
- Liu, Y.; Su, W.; Wang, L.; Lei, J.; Chai, S.; Zhang, W.; Yang, X. Integrated transcriptome, small RNA and degradome sequencing approaches proffer insights into chlorogenic acid biosynthesis in leafy sweet potato. PLoS ONE 2021, 16, e0245266. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Wu, Y.S.; Yang, F.; Gong, W.Z.; Ahmed, S.; Fan, Y.F.; Wu, X.L.; Yong, T.W.; Liu, W.G.; Shu, K.; Liu, J.; et al. Shade adaptive response and yield analysis of different soybean genotypes in relay intercropping systems. J. Integr. Agric. 2017, 16, 1331–1340. [Google Scholar] [CrossRef]
- Milenković, L.; Mastilović, J.; Kevrešan, Ž.; Bajić, A.; Gledić, A.; Stanojević, L.; Cvetković, D.; Šunić, L.; Ilić, Z.S. Effect of shading and grafting on yield and quality of tomato. J. Sci. Food Agric. 2019, 100, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Ye, K.; Xu, Y.; Zhao, Y.; Zhao, D. Effect of shading on the morphological, physiological, and biochemical characteristics as well as the transcriptome of matcha green tea. Int. J. Mol. Sci. 2022, 23, 14169. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Feng, L.; Liu, Q.; Wu, X.; Fan, Y.; Raza, M.A.; Cheng, Y.; Chen, J.; Wang, X.; Yong, T. Effect of interactions between light intensity and red-to-far-red ratio on the photosynthesis of soybean leaves under shade condition. Environ. Exp. Bot. 2018, 150, 79–87. [Google Scholar] [CrossRef]
- Chen, M. Chlorophyll modifications and their spectral extension in oxygenic photosynthesis. Annu. Rev. Biochem. 2014, 83, 317–340. [Google Scholar] [CrossRef]
- Loughlin, P.C.; Willows, R.D.; Chen, M. In vitro conversion of vinyl to formyl groups in naturally occurring chlorophylls. Sci. Rep. 2014, 4, 6069. [Google Scholar] [CrossRef]
- Gong, X.; Liu, C.; Dang, K.; Wang, H.; Du, W.; Qi, H.; Jiang, Y.; Feng, B. Mung bean (Vigna radiata L.) source leaf adaption to shading stress affects not only photosynthetic physiology metabolism but also control of key gene expression. Front. Plant Sci. 2022, 13, 753264. [Google Scholar] [CrossRef]
- Luo, Y.L. The Effect of Ridging Modes on Growth, Photosynthetic Physiology and Yield of Sweet Potato in Relay-Cropping System. Master’s Thesis, Southwest University, Chongqing, China, 2017. [Google Scholar]
- Björn, L.O.; Papageorgiou, G.C.; Blankenship, R.E.; Govindjee. A viewpoint: Why chlorophyll a? Photosynth. Res. 2009, 99, 85–98. [Google Scholar] [CrossRef]
- Field, K.J.; George, R.; Fearn, B.; Quick, W.P.; Davey, M.P. Best of both worlds: Simultaneous high-light and shade-tolerance adaptations within individual leaves of the living stone Lithops aucampiae. PLoS ONE 2013, 8, e75671. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Zhou, H.; Zhu, Q.; Li, C.; Zhang, H.; Wu, J.J.; Xie, F. Photosynthetic response of soybean leaf to wide light-fluctuation in maize-soybean intercropping system. Front. Plant Sci. 2017, 8, 1695. [Google Scholar] [CrossRef] [PubMed]
- Tiessen, A.; Padilla-Chacon, D. Subcellular compartmentation of sugar signaling: Links among carbon cellular status, route of sucrolysis, sink-source allocation, and metabolic partitioning. Front. Plant Sci. 2013, 3, 306. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.N.; Chen, J.J.; Bian, Q.Q.; Hu, L.L.; Zhang, L.; Yin, Y.M.; Qiao, S.C.; Cao, G.Z.; Kang, Z.H.; Zhao, G.R.; et al. Integrated analysis of transcriptome and metabolome reveals the metabolic response pathways of sweetpotato under shade stress. Acta Agron. Sin. 2023, 49, 1785–1798. [Google Scholar]
- Wang, Y.D. Effects of Different Shading Intensities on the Growth and Anthocyanin Synthesis of Purple-Fleshed Sweet Potatoes. Master’s Thesis, Hainan University, Haikou, China, 2022. [Google Scholar]
- Mittler, R.; Zandalinas, S.I.; Fichman, Y.; Van Breusegem, F. Reactive oxygen species signalling in plant stress responses. Nat. Rev. Mol. Cell Biol. 2022, 23, 663–679. [Google Scholar] [CrossRef] [PubMed]
- Boeckx, T.; Winters, A.L.; Webb, K.J.; Kingston-Smith, A.H. Polyphenol oxidase in leaves: Is there any significance to the chloroplastic localization? J. Exp. Bot. 2015, 66, 3571–3579. [Google Scholar] [CrossRef]
- Skorupa, M.; Szczepanek, J.; Yolcu, S.; Mazur, J.; Tretyn, A.; Tyburski, J. Characteristic of the ascorbate oxidase gene family in Beta vulgaris and analysis of the role of AAO in response to salinity and drought in beet. Int. J. Mol. Sci. 2022, 23, 12773. [Google Scholar] [CrossRef]
- Park, S.Y.; Ryu, S.H.; Kwon, S.Y.; Lee, H.S.; Kim, J.G.; Kwak, S.S. Differential expression of six novel peroxidase cDNAs from cell cultures of sweetpotato in response to stress. Mol. Genet. Genom. 2003, 269, 542–552. [Google Scholar] [CrossRef]
- Chen, S.; Wu, F.; Li, Y.; Qian, Y.; Pan, X.; Li, F.; Wang, Y.; Wu, Z.; Fu, C.; Lin, H.; et al. NtMYB4 and NtCHS1 are critical factors in the regulation of flavonoid biosynthesis and are involved in salinity responsiveness. Front. Plant Sci. 2019, 10, 178. [Google Scholar] [CrossRef]
- Tan, B.C.; Tan, S.K.; Wong, S.M.; Ata, N.; Rahman, N.A.; Khalid, N. Distribution of flavonoids and cyclohexenyl chalcone derivatives in conventional propagated and in vitro-derived field-grown Boesenbergia rotunda (L.) Mansf. Evid.-Based Complement. Altern. Med. 2015, 2015, 451870. [Google Scholar] [CrossRef]
- Isa, N.M.; Abdelwahab, S.I.; Mohan, S.; Abdul, A.B.; Sukari, M.A.; Taha, M.M.; Syam, S.; Narrima, P.; Cheah, S.C.; Ahmad, S.; et al. In vitro anti-inflammatory, cytotoxic and antioxidant activities of boesenbergin A, a chalcone isolated from Boesenbergia rotunda (L.) (fingerroot). Braz. J. Med. Biol. Res. 2012, 45, 524–530. [Google Scholar] [CrossRef]
- Karlson, C.K.S.; Mohd Noor, S.N.; Khalid, N.; Tan, B.C. CRISPRi-mediated down-regulation of the cinnamate-4-hydroxylase (C4H) gene enhances the flavonoid biosynthesis in Nicotiana tabacum. Biology 2022, 11, 1127. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Li, Z.; Wang, J. Transcriptional analyses reveal the molecular mechanism governing shade tolerance in the invasive plant Solidago canadensis. Ecol. Evol. 2020, 10, 4391–4406. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Mumtaz, M.A.; Zhang, Y.; Shu, H.; Hao, Y.; Lu, X.; Cheng, S.; Zhu, G.; Wang, Z. Response of anthocyanin accumulation in pepper (Capsicum annuum) fruit to light days. Int. J. Mol. Sci. 2022, 23, 8357. [Google Scholar] [CrossRef] [PubMed]
- An, X.; Tan, T.; Song, Z.; Guo, X.; Zhang, X.; Zhu, Y.; Wang, D. Physiological response of anthocyanin synthesis to different light intensities in blueberry. PLoS ONE 2023, 18, e0283284. [Google Scholar] [CrossRef] [PubMed]
- Khatri, P.; Chen, L.; Rajcan, I.; Dhaubhadel, S. Functional characterization of Cinnamate 4-hydroxylase gene family in soybean (Glycine max). PLoS ONE 2023, 18, e0285698. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Huang, H.; Gao, Q.; Li, Y.; Xiang, H.; Zeng, W.; Xu, L.; Liu, X.; Li, J.; Mi, Q.; et al. Effects of editing DFR genes on flowers, leaves, and roots of tobacco. BMC Plant Biol. 2023, 23, 349. [Google Scholar] [CrossRef] [PubMed]
- Shaipulah, N.F.; Muhlemann, J.K.; Woodworth, B.D.; Van Moerkercke, A.; Verdonk, J.C.; Ramirez, A.A.; Haring, M.A.; Dudareva, N.; Schuurink, R.C. CCoAOMT down-regulation activates anthocyanin biosynthesis in petunia. Plant Physiol. 2016, 170, 717–731. [Google Scholar] [CrossRef] [PubMed]
- Song, J.L.; Wang, Z.Y.; Wang, Y.H.; Du, J.; Wang, C.Y.; Zhang, X.Q.; Chen, S.; Huang, X.L.; Xie, X.M.; Zhong, T.X. Overexpression of Pennisetum purpureum CCoAOMT contributes to lignin deposition and drought tolerance by promoting the accumulation of flavonoids in transgenic tobacco. Front. Plant Sci. 2022, 13, 884456. [Google Scholar] [CrossRef]
- Zhang, M.; Su, J.; Zhang, Y.; Xu, J.; Zhang, S. Conveying endogenous and exogenous signals: MAPK cascades in plant growth and defense. Curr. Opin. Plant Biol. 2018, 45, 1–10. [Google Scholar] [CrossRef]
- Jagodzik, P.; Tajdel-Zielinska, M.; Ciesla, A.; Marczak, M.; Ludwikow, A. Mitogen-activated protein kinase cascades in plant hormone signaling. Front. Plant Sci. 2018, 9, 1387. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhang, S. Mitogen-activated protein kinase cascades in plant signaling. J. Integr. Plant Biol. 2022, 64, 301–341. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, E.; Correa Galvis, V.; Armbruster, U. Efficient photosynthesis in dynamic light environments: A chloroplast’s perspective. Biochem. J. 2019, 476, 2725–2741. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Grimm, B. Connecting chlorophyll metabolism with accumulation of the photosynthetic apparatus. Trends Plant Sci. 2021, 26, 484–495. [Google Scholar] [CrossRef] [PubMed]
- Aryamanesh, N.; Ruwe, H.; Sanglard, L.V.; Eshraghi, L.; Bussell, J.D.; Howell, K.A.; Small, I.; des Francs-Small, C.C. The pentatricopeptide repeat protein EMB2654 is essential for trans-splicing of a chloroplast small ribosomal subunit transcript. Plant Physiol. 2017, 173, 1164–1176. [Google Scholar] [CrossRef] [PubMed]
- Zeng, C.; Jiao, Q.; Jia, T.; Hu, X. Updated progress on group II intron splicing factors in plant chloroplasts. Curr. Issues Mol. Biol. 2022, 44, 4229–4239. [Google Scholar] [CrossRef] [PubMed]
- Andrade-Marcial, M.; Pacheco-Arjona, R.; Góngora-Castillo, E.; De-la-Peña, C. Chloroplastic pentatricopeptide repeat proteins (PPR) in albino plantlets of Agave angustifolia Haw. reveal unexpected behavior. BMC Plant Biol. 2022, 22, 352. [Google Scholar] [CrossRef] [PubMed]
- Asakura, Y.; Barkan, A. A CRM domain protein functions dually in group I and group II intron splicing in land plant chloroplasts. Plant Cell 2007, 19, 3864–3875. [Google Scholar] [CrossRef]
- Hammani, K.; Barkan, A. An mTERF domain protein functions in group II intron splicing in maize chloroplasts. Nucleic Acids Res. 2014, 42, 5033–5042. [Google Scholar] [CrossRef]
- Small, I.D.; Peeters, N. The PPR motif-a TPR-related motif prevalent in plant organellar proteins. Trends Biochem. Sci. 2000, 25, 46–47. [Google Scholar] [CrossRef]
- Lermontova, I.; Grimm, B. Reduced activity of plastid protoporphyrinogen oxidase causes attenuated photodynamic damage during high-light compared to low-light exposure. Plant J. 2006, 48, 499–510. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.Y.; Shi, J.C.; Wang, D.J.; Liang, X.; Wei, F.; Gong, C.M.; Qiu, L.J.; Zhou, H.C.; Folta, K.M.; Wen, Y.Q.; et al. A point mutation in the gene encoding magnesium chelatase I subunit influences strawberry leaf color and metabolism. Plant Physiol. 2023, 192, 2737–2755. [Google Scholar] [CrossRef] [PubMed]
- Bellaoui, M.; Keddie, J.S.; Gruissem, W. DCL is a plant-specific protein required for plastid ribosomal RNA processing and embryo development. Plant Mol. Biol. 2003, 53, 531–543. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, M.; Sparvoli, S.; Edmunds, C.; Garosi, P.; Findlay, K.; Martin, C. DAG, a gene required for chloroplast differentiation and palisade development in Antirrhinum majus. EMBO J. 1996, 15, 4194–4207. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Xu, F.; Guan, L.; Qian, P.; Liu, Y.; Zhang, H.; Huang, Y.; Hou, S. The tetratricopeptide repeat-containing protein slow green1 is required for chloroplast development in Arabidopsis. J. Exp. Bot. 2014, 65, 1111–1123. [Google Scholar] [CrossRef] [PubMed]
- Kovács-Bogdán, E.; Benz, J.P.; Soll, J.; Bölter, B. Tic20 forms a channel independent of Tic110 in chloroplasts. BMC Plant Biol. 2011, 11, 133. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Y.; Sun, J.L.; Tian, Y.Y.; Liu, J.X. The FtsH-inactive protein FtsHi5 is required for chloroplast development and protein accumulation in chloroplasts at low ambient temperature in Arabidopsis. Front. Plant Sci. 2022, 12, 830390. [Google Scholar] [CrossRef]
- Lin, D.; Jiang, Q.; Zheng, K.; Chen, S.; Zhou, H.; Gong, X.; Xu, J.; Teng, S.; Dong, Y. Mutation of the rice ASL2 gene encoding plastid ribosomal protein L21 causes chloroplast developmental defects and seedling death. Plant Biol. 2015, 17, 599–607. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety | Treatment | Yield (kg) |
|---|---|---|
| Eshu10 | CK | 1.35 ± 0.29 a |
| L30 | 0.62 ± 0.10 b | |
| L50 | 0.52 ± 0.08 b | |
| L70 | 0.12 ± 0.02 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jing, X.; Chen, P.; Jin, X.; Lei, J.; Wang, L.; Chai, S.; Yang, X. Physiological, Photosynthetic, and Transcriptomics Insights into the Influence of Shading on Leafy Sweet Potato. Genes 2023, 14, 2112. https://doi.org/10.3390/genes14122112
Jing X, Chen P, Jin X, Lei J, Wang L, Chai S, Yang X. Physiological, Photosynthetic, and Transcriptomics Insights into the Influence of Shading on Leafy Sweet Potato. Genes. 2023; 14(12):2112. https://doi.org/10.3390/genes14122112
Chicago/Turabian StyleJing, Xiaojing, Peiru Chen, Xiaojie Jin, Jian Lei, Lianjun Wang, Shasha Chai, and Xinsun Yang. 2023. "Physiological, Photosynthetic, and Transcriptomics Insights into the Influence of Shading on Leafy Sweet Potato" Genes 14, no. 12: 2112. https://doi.org/10.3390/genes14122112
APA StyleJing, X., Chen, P., Jin, X., Lei, J., Wang, L., Chai, S., & Yang, X. (2023). Physiological, Photosynthetic, and Transcriptomics Insights into the Influence of Shading on Leafy Sweet Potato. Genes, 14(12), 2112. https://doi.org/10.3390/genes14122112