
OCT and ERG Techniques in High-Throughput Phenotyping of Mouse Vision

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. OCT
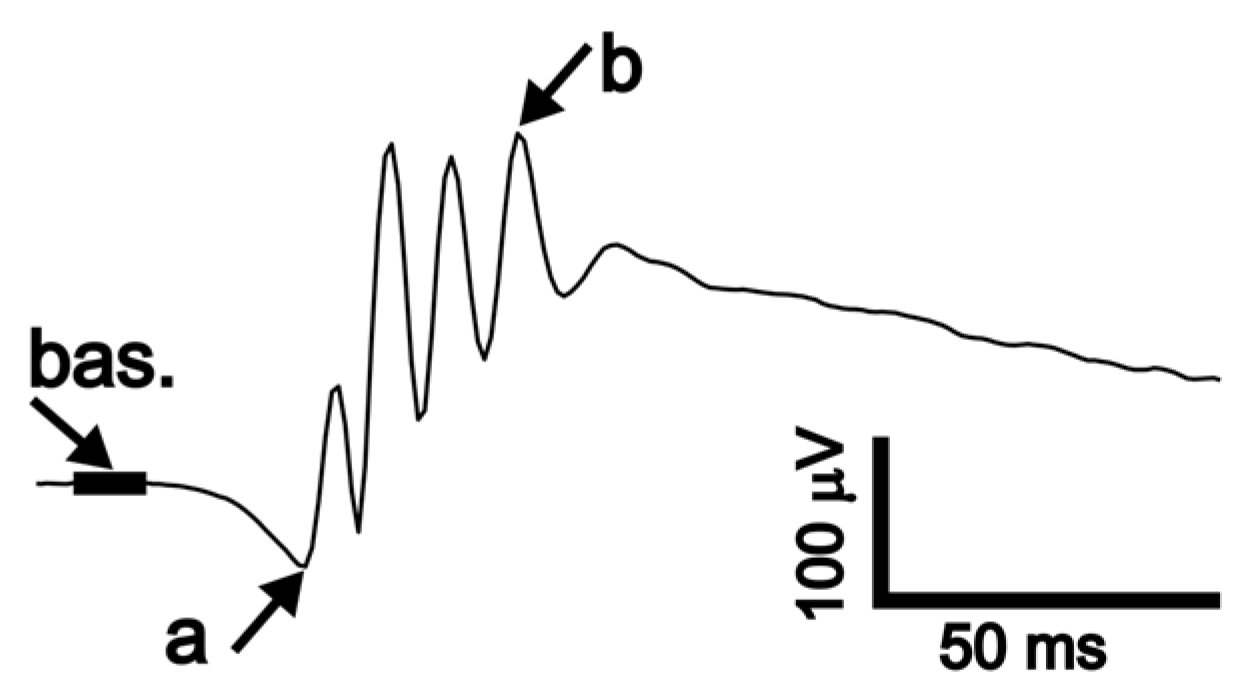
2.3. ERG
2.4. Statistical Analysis
3. Results
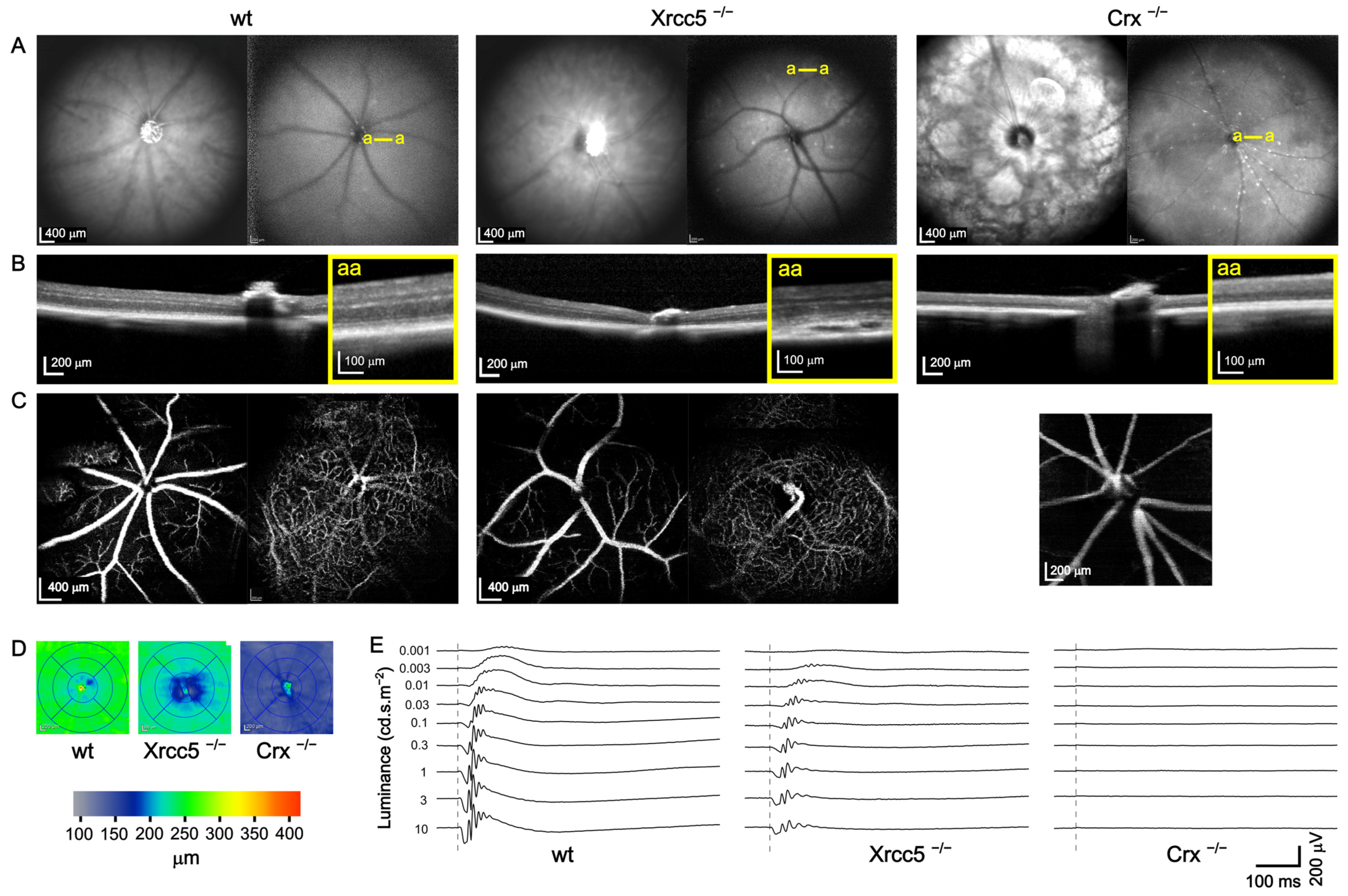
3.1. Eye-Screening Procedure
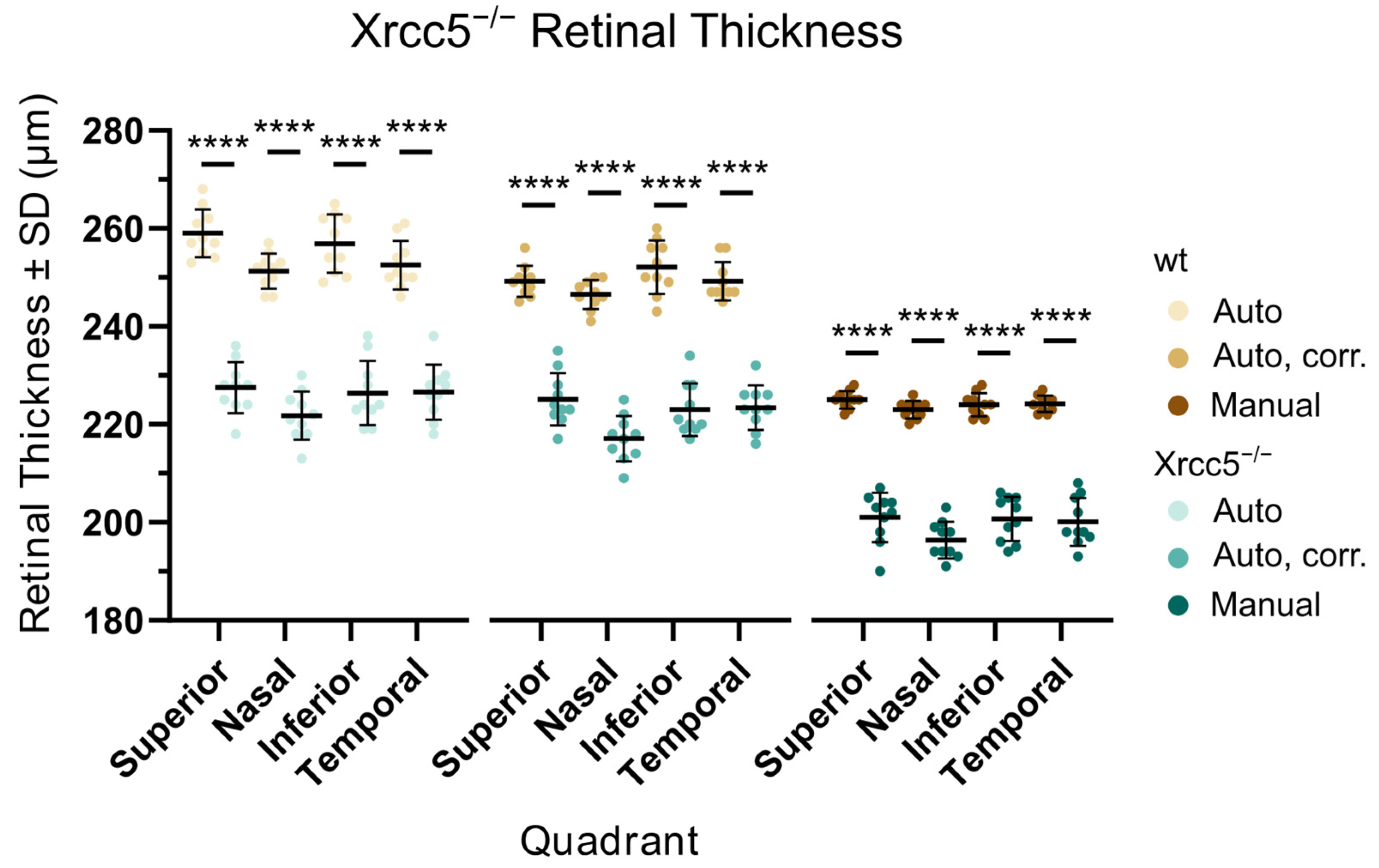
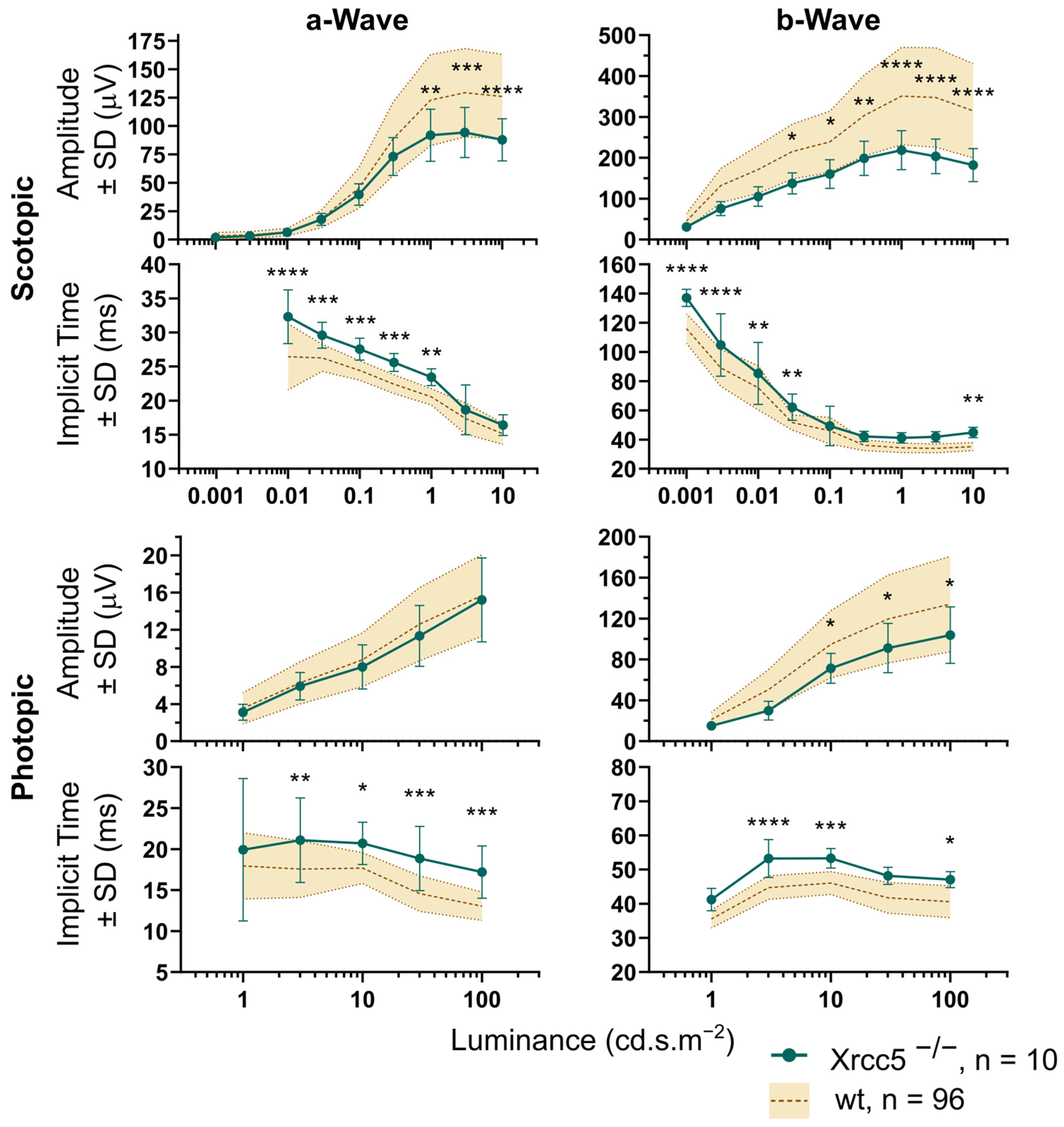
3.2. Xrcc5−/−
3.3. Crx−/−
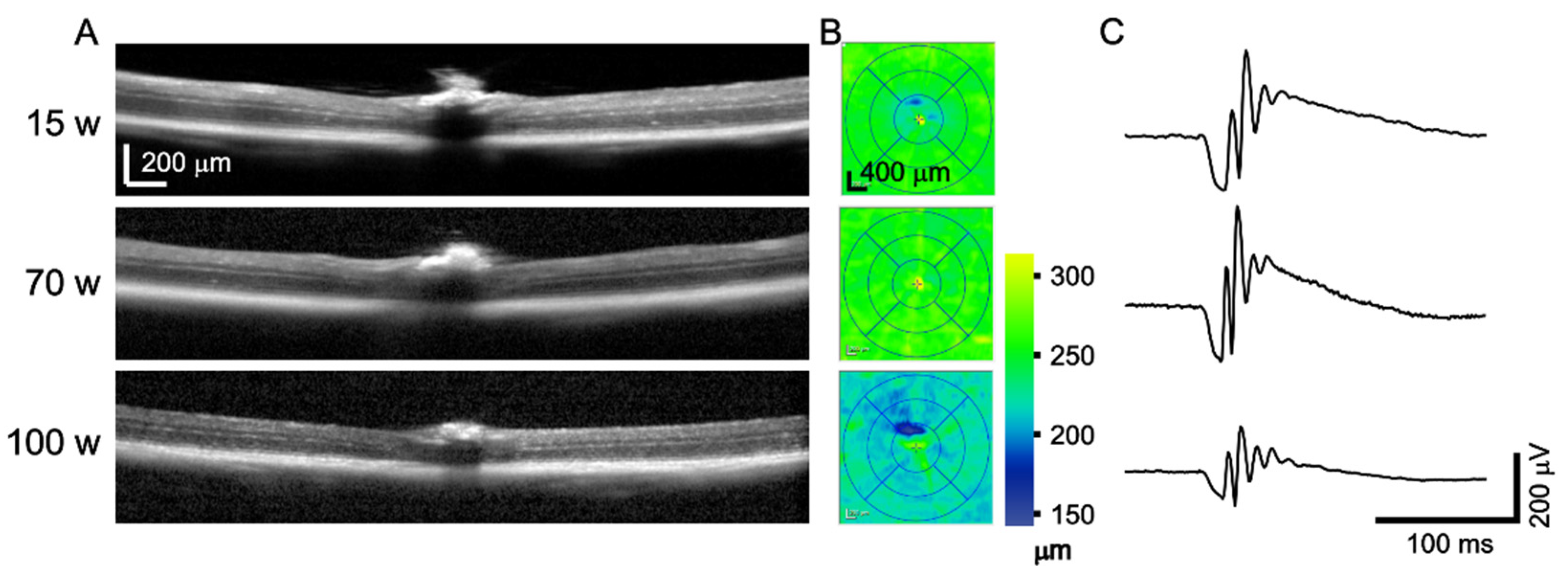
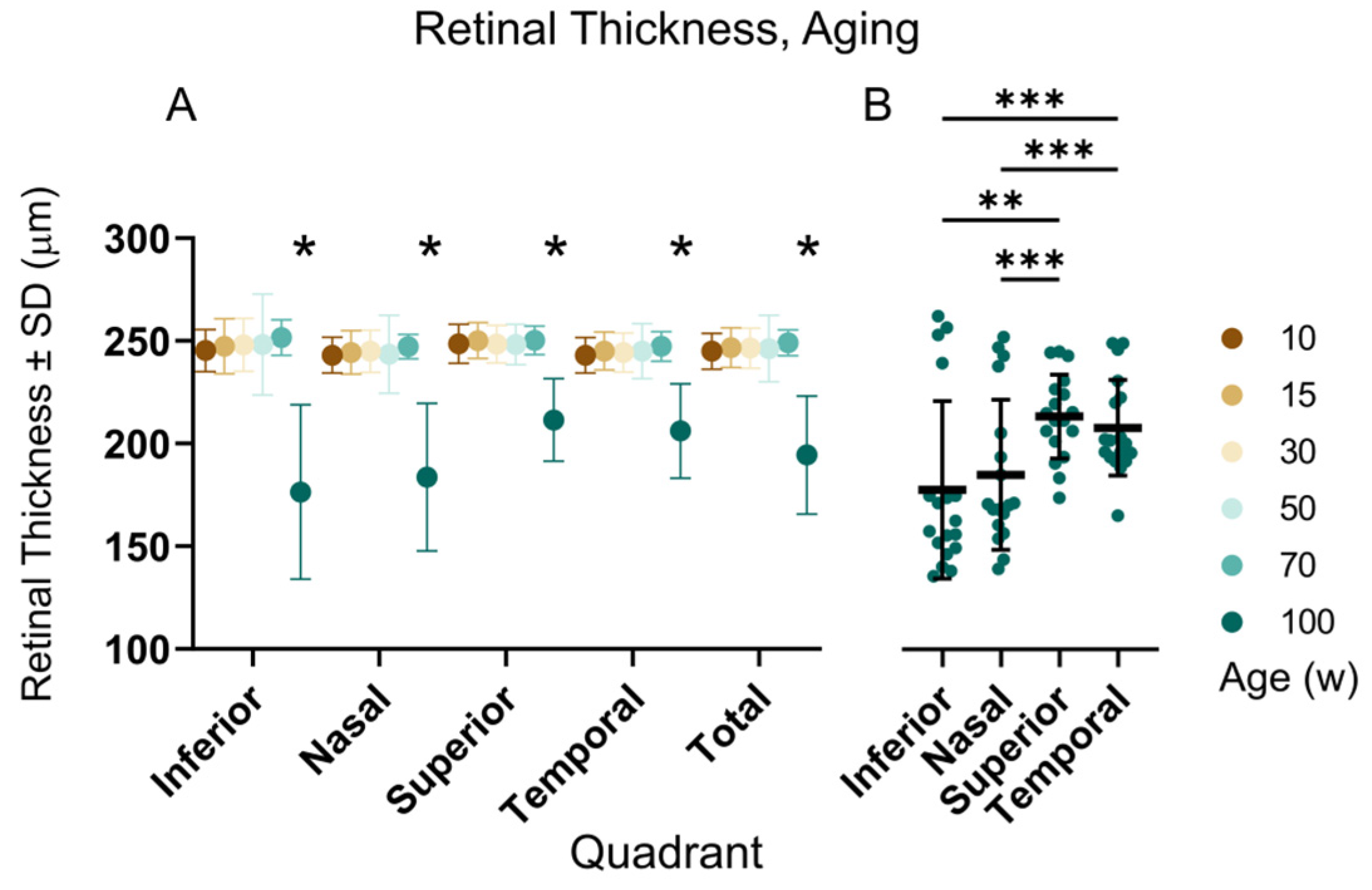
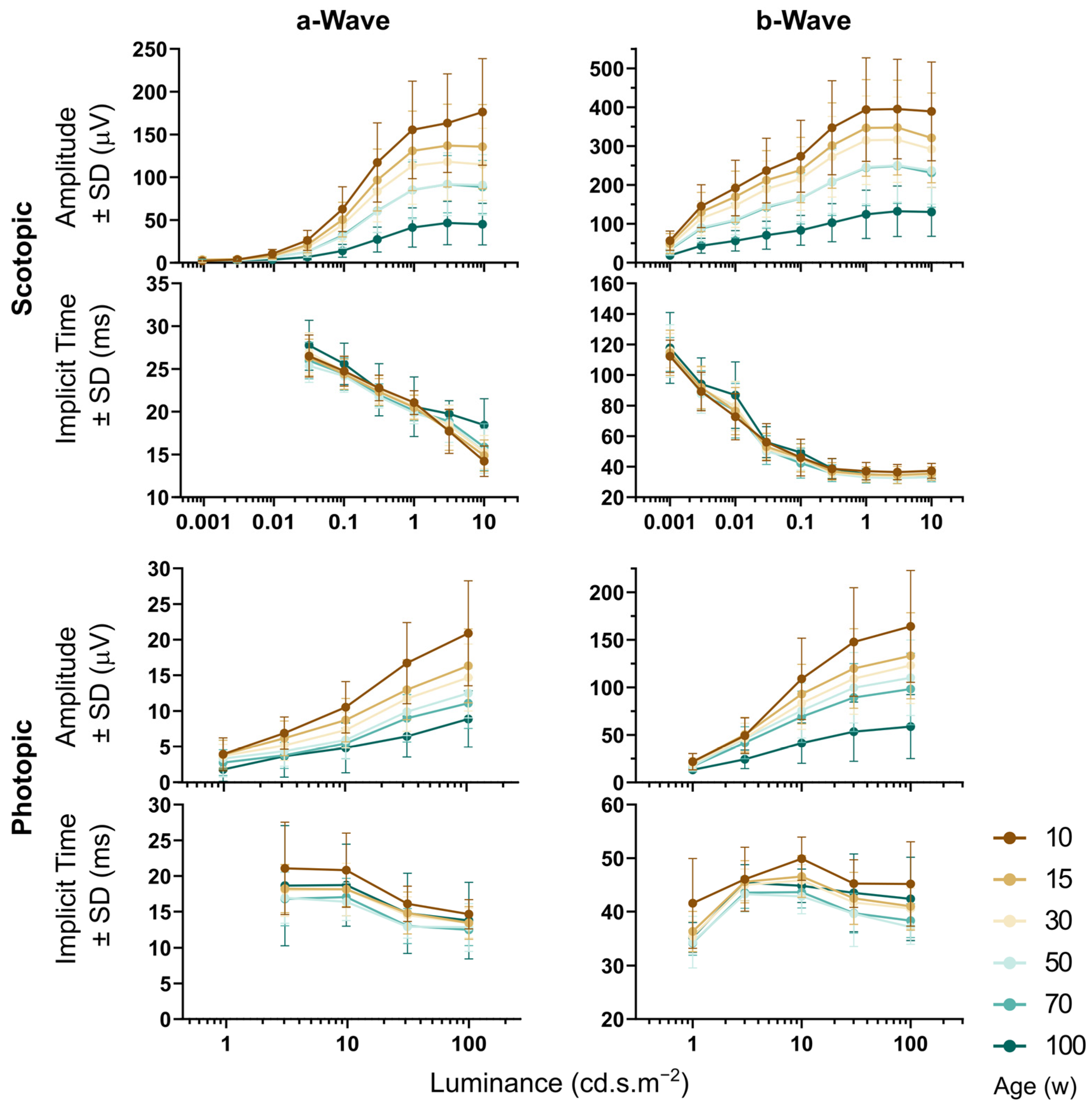
3.4. OCT and ERG during Aging
4. Discussion
4.1. Eye-Screening Procedure
4.2. Xrcc5−/−
4.3. Crx−/−
4.4. OCT and ERG during Aging
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mouse Genome Sequencing, C.; Waterston, R.H.; Lindblad-Toh, K.; Birney, E.; Rogers, J.; Abril, J.F.; Agarwal, P.; Agarwala, R.; Ainscough, R.; Alexandersson, M.; et al. Initial sequencing and comparative analysis of the mouse genome. Nature 2002, 420, 520–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antony, B.J.; Kim, B.J.; Lang, A.; Carass, A.; Prince, J.L.; Zack, D.J. Automated segmentation of mouse OCT volumes (ASiMOV): Validation & clinical study of a light damage model. PLoS ONE 2017, 12, e0181059. [Google Scholar] [CrossRef] [Green Version]
- Fischer, M.D.; Huber, G.; Beck, S.C.; Tanimoto, N.; Muehlfriedel, R.; Fahl, E.; Grimm, C.; Wenzel, A.; Reme, C.E.; van de Pavert, S.A.; et al. Noninvasive, in vivo assessment of mouse retinal structure using optical coherence tomography. PLoS ONE 2009, 4, e7507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giannakaki-Zimmermann, H.; Kokona, D.; Wolf, S.; Ebneter, A.; Zinkernagel, M.S. Optical Coherence Tomography Angiography in Mice: Comparison with Confocal Scanning Laser Microscopy and Fluorescein Angiography. Transl. Vis. Sci. Technol. 2016, 5, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huber, G.; Beck, S.C.; Grimm, C.; Sahaboglu-Tekgoz, A.; Paquet-Durand, F.; Wenzel, A.; Humphries, P.; Redmond, T.M.; Seeliger, M.W.; Fischer, M.D. Spectral domain optical coherence tomography in mouse models of retinal degeneration. Investig. Ophthalmol. Vis. Sci. 2009, 50, 5888–5895. [Google Scholar] [CrossRef]
- Ochakovski, G.A.; Fischer, M.D. Phenotyping of Mouse Models with OCT. Methods Mol. Biol. 2019, 1834, 285–291. [Google Scholar] [CrossRef]
- Frishman, L.J.; Wang, M.H. Electroretinogram of Human, Monkey and Mouse. In Adler’s Physiology of the Eye; Elsevier: Amsterdam, The Netherlands, 2011. [Google Scholar]
- Perlman, I. The Electroretinogram: ERG by Ido Perlman. Available online: https://webvision.med.utah.edu/book/electrophysiology/the-electroretinogram-erg/ (accessed on 24 October 2022).
- Penn, R.D.; Hagins, W.A. Signal transmission along retinal rods and the origin of the electroretinographic a-wave. Nature 1969, 223, 201–204. [Google Scholar] [CrossRef]
- Green, D.G.; Kapousta-Bruneau, N.V. A dissection of the electroretinogram from the isolated rat retina with microelectrodes and drugs. Vis. Neurosci. 1999, 16, 727–741. [Google Scholar] [CrossRef]
- Moore, B.A.; Leonard, B.C.; Sebbag, L.; Edwards, S.G.; Cooper, A.; Imai, D.M.; Straiton, E.; Santos, L.; Reilly, C.; Griffey, S.M.; et al. Identification of genes required for eye development by high-throughput screening of mouse knockouts. Commun. Biol. 2018, 1, 236. [Google Scholar] [CrossRef] [Green Version]
- Peachey, N.S.; Ball, S.L. Electrophysiological analysis of visual function in mutant mice. Doc. Ophthalmol. 2003, 107, 13–36. [Google Scholar] [CrossRef]
- Schmucker, C.; Schaeffel, F. In vivo biometry in the mouse eye with low coherence interferometry. Vis. Res. 2004, 44, 2445–2456. [Google Scholar] [CrossRef] [Green Version]
- Schmucker, C.; Schaeffel, F. A paraxial schematic eye model for the growing C57BL/6 mouse. Vis. Res. 2004, 44, 1857–1867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, B.A.; Roux, M.J.; Sebbag, L.; Cooper, A.; Edwards, S.G.; Leonard, B.C.; Imai, D.M.; Griffey, S.; Bower, L.; Clary, D.; et al. A Population Study of Common Ocular Abnormalities in C57BL/6N rd8 Mice. Investig. Ophthalmol. Vis. Sci. 2018, 59, 2252–2261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobs, G.H.; Neitz, J.; Deegan, J.F. Retinal receptors in rodents maximally sensitive to ultraviolet light. Nature 1991, 353, 655–656. [Google Scholar] [CrossRef] [PubMed]
- Calderone, J.B.; Jacobs, G.H. Regional variations in the relative sensitivity to UV light in the mouse retina. Vis. Neurosci. 1995, 12, 463–468. [Google Scholar] [CrossRef]
- Jeon, C.J.; Strettoi, E.; Masland, R.H. The major cell populations of the mouse retina. J. Neurosci. 1998, 18, 8936–8946. [Google Scholar] [CrossRef] [Green Version]
- Kostic, C.; Arsenijevic, Y. Animal modelling for inherited central vision loss. J. Pathol. 2016, 238, 300–310. [Google Scholar] [CrossRef] [Green Version]
- Frishman, L.; Sustar, M.; Kremers, J.; McAnany, J.J.; Sarossy, M.; Tzekov, R.; Viswanathan, S. ISCEV extended protocol for the photopic negative response (PhNR) of the full-field electroretinogram. Doc. Ophthalmol. 2018, 136, 207–211. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.A.; Jeffrey, B.G.; Messias, A.M.V.; Robson, A.G. ISCEV extended protocol for the stimulus-response series for the dark-adapted full-field ERG b-wave. Doc. Ophthalmol. 2019, 138, 217–227. [Google Scholar] [CrossRef] [Green Version]
- McCulloch, D.L.; Kondo, M.; Hamilton, R.; Lachapelle, P.; Messias, A.M.V.; Robson, A.G.; Ueno, S. ISCEV extended protocol for the stimulus-response series for light-adapted full-field ERG. Doc. Ophthalmol. 2019, 138, 205–215. [Google Scholar] [CrossRef]
- McCulloch, D.L.; Marmor, M.F.; Brigell, M.G.; Hamilton, R.; Holder, G.E.; Tzekov, R.; Bach, M. ISCEV Standard for full-field clinical electroretinography (2015 update). Doc. Ophthalmol. 2015, 130, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sustar, M.; Holder, G.E.; Kremers, J.; Barnes, C.S.; Lei, B.; Khan, N.W.; Robson, A.G. ISCEV extended protocol for the photopic On-Off ERG. Doc. Ophthalmol. 2018, 136, 199–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, I.N.; Yang, J.T.; Huang, C.; Huang, H.C.; Wu, Y.P.; Chen, J.C. Elevated XRCC5 expression level can promote temozolomide resistance and predict poor prognosis in glioblastoma. Oncol. Lett. 2021, 21, 443. [Google Scholar] [CrossRef] [PubMed]
- Savva, C.; Sadiq, M.; Sheikh, O.; Karim, S.; Trivedi, S.; Green, A.R.; Rakha, E.A.; Madhusudan, S.; Arora, A. Werner Syndrome Protein Expression in Breast Cancer. Clin. Breast Cancer 2021, 21, 57–73.e57. [Google Scholar] [CrossRef] [PubMed]
- Shang, B.; Jia, Y.; Chen, G.; Wang, Z. Ku80 correlates with neoadjuvant chemotherapy resistance in human lung adenocarcinoma, but reduces cisplatin/pemetrexed-induced apoptosis in A549 cells. Respir. Res. 2017, 18, 56. [Google Scholar] [CrossRef] [Green Version]
- Oshitari, T.; Kitahashi, M.; Mizuno, S.; Baba, T.; Kubota-Taniai, M.; Takemoto, M.; Yokote, K.; Yamamoto, S.; Roy, S. Werner syndrome with refractory cystoid macular edema and immunohistochemical analysis of WRN proteins in human retinas. BMC Ophthalmol. 2014, 14, 31. [Google Scholar] [CrossRef] [Green Version]
- Muller, B.; Serafin, F.; Laucke, L.L.; Rheinhard, W.; Wimmer, T.; Stieger, K. Characterization of Double-Strand Break Repair Protein Ku80 Location Within the Murine Retina. Investig. Ophthalmol. Vis. Sci. 2022, 63, 22. [Google Scholar] [CrossRef]
- Furukawa, T.; Morrow, E.M.; Li, T.; Davis, F.C.; Cepko, C.L. Retinopathy and attenuated circadian entrainment in Crx-deficient mice. Nat. Genet. 1999, 23, 466–470. [Google Scholar] [CrossRef]
- Berger, W.; Kloeckener-Gruissem, B.; Neidhardt, J. The molecular basis of human retinal and vitreoretinal diseases. Prog. Retin. Eye Res. 2010, 29, 335–375. [Google Scholar] [CrossRef] [Green Version]
- Tran, N.M.; Chen, S. Mechanisms of blindness: Animal models provide insight into distinct CRX-associated retinopathies. Dev. Dyn. 2014, 243, 1153–1166. [Google Scholar] [CrossRef]
- Gresh, J.; Goletz, P.W.; Crouch, R.K.; Rohrer, B. Structure-function analysis of rods and cones in juvenile, adult, and aged C57bl/6 and Balb/c mice. Vis. Neurosci. 2003, 20, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Grenell, A.; Zhong, F.; Yam, M.; Hauer, A.; Gregor, E.; Zhu, S.; Lohner, D.; Zhu, J.; Du, J. Metabolic signature of the aging eye in mice. Neurobiol. Aging 2018, 71, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Tsantilas, K.A.; Cleghorn, W.M.; Bisbach, C.M.; Whitson, J.A.; Hass, D.T.; Robbings, B.M.; Sadilek, M.; Linton, J.D.; Rountree, A.M.; Valencia, A.P.; et al. An Analysis of Metabolic Changes in the Retina and Retinal Pigment Epithelium of Aging Mice. Investig. Ophthalmol. Vis. Sci. 2021, 62, 20. [Google Scholar] [CrossRef] [PubMed]
- Kolesnikov, A.V.; Fan, J.; Crouch, R.K.; Kefalov, V.J. Age-related deterioration of rod vision in mice. J. Neurosci. 2010, 30, 11222–11231. [Google Scholar] [CrossRef] [Green Version]
- Salobrar-Garcia, E.; Lopez-Cuenca, I.; Sanchez-Puebla, L.; de Hoz, R.; Fernandez-Albarral, J.A.; Ramirez, A.I.; Bravo-Ferrer, I.; Medina, V.; Moro, M.A.; Saido, T.C.; et al. Retinal Thickness Changes Over Time in a Murine AD Model APP (NL-F/NL-F). Front. Aging Neurosci. 2020, 12, 625642. [Google Scholar] [CrossRef]
- Ferdous, S.; Liao, K.L.; Gefke, I.D.; Summers, V.R.; Wu, W.; Donaldson, K.J.; Kim, Y.K.; Sellers, J.T.; Dixon, J.A.; Shelton, D.A.; et al. Age-Related Retinal Changes in Wild-Type C57BL/6J Mice Between 2 and 32 Months. Investig. Ophthalmol. Vis. Sci. 2021, 62, 9. [Google Scholar] [CrossRef]
- Dysli, C.; Enzmann, V.; Sznitman, R.; Zinkernagel, M.S. Quantitative Analysis of Mouse Retinal Layers Using Automated Segmentation of Spectral Domain Optical Coherence Tomography Images. Transl. Vis. Sci. Technol. 2015, 4, 9. [Google Scholar] [CrossRef] [Green Version]
- Mattapallil, M.J.; Wawrousek, E.F.; Chan, C.C.; Zhao, H.; Roychoudhury, J.; Ferguson, T.A.; Caspi, R.R. The Rd8 mutation of the Crb1 gene is present in vendor lines of C57BL/6N mice and embryonic stem cells, and confounds ocular induced mutant phenotypes. Investig. Ophthalmol. Vis. Sci. 2012, 53, 2921–2927. [Google Scholar] [CrossRef] [Green Version]
- Moshiri, A. Animals Models of Inherited Retinal Disease. Int. Ophthalmol. Clin. 2021, 61, 113–130. [Google Scholar] [CrossRef]
- Aleman, T.S.; Cideciyan, A.V.; Aguirre, G.K.; Huang, W.C.; Mullins, C.L.; Roman, A.J.; Sumaroka, A.; Olivares, M.B.; Tsai, F.F.; Schwartz, S.B.; et al. Human CRB1-associated retinal degeneration: Comparison with the rd8 Crb1-mutant mouse model. Investig. Ophthalmol. Vis. Sci. 2011, 52, 6898–6910. [Google Scholar] [CrossRef]
- Saul, A.B.; Cui, X.; Markand, S.; Smith, S.B. Detailed electroretinographic findings in rd8 mice. Doc. Ophthalmol. 2017, 134, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Rosch, S.; Johnen, S.; Muller, F.; Pfarrer, C.; Walter, P. Correlations between ERG, OCT, and Anatomical Findings in the rd10 Mouse. J. Ophthalmol. 2014, 2014, 874751. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Unit | Wt (Mean ± SD, n) | KO (Mean ± SD, n) | p |
|---|---|---|---|---|
| Total retinal thickness, autosegmentation | µm | 254.9 ± 3.1, 10 | 225.6 ± 4.2, 10 | <0.0001 |
| Total retinal thickness, auto with corrections | µm | 249.3 ± 3.0, 10 | 222.2 ± 3.7, 10 | <0.0001 |
| Total retinal thickness, manual measurement | µm | 224.1 ± 1.3, 10 | 199.6 ± 2.5, 10 | <0.0001 |
| Scotopic a amplitude, 10 cd.s/m2 | µV | 126.0 ± 37.1, 96 | 87.7 ± 18.7, 10 | <0.0001 |
| Scotopic b amplitude, 10 cd.s/m2 | µV | 314.9 ± 115.5, 96 | 181.9 ± 40.5, 10 | <0.0001 |
| Photopic a amplitude, 100 cd.s/m2 | µV | 15.7 ± 4.4, 96 | 15.2 ± 4.5, 10 | 0.9956 |
| Photopic b amplitude, 100 cd.s/m2 | µV | 134.1 ± 46.7, 96 | 103.8 ± 27.7, 10 | 0.0245 |
| Scotopic a implicit time, 10 cd.s/m2 | ms | 15.1 ± 1.5, 96 | 16.4 ± 1.5, 10 | 0.5465 |
| Scotopic b implicit time, 10 cd.s/m2 | ms | 35.1 ± 2.8, 96 | 44.8 ± 3.6, 10 | 0.0086 |
| Photopic a implicit time, 100 cd.s/m2 | ms | 13.0 ± 1.7, 96 | 17.2 ± 3.2, 10 | 0.0003 |
| Photopic b implicit time, 100 cd.s/m2 | ms | 40.6 ± 4.7, 96 | 47.1 ± 2.4, 10 | 0.0248 |
| Parameter | Unit | 10 w | 15 w | 30 w | 50 w | 70 w | 100 w |
|---|---|---|---|---|---|---|---|
| Total retinal thickness, auto-segmentation | µm | 244.9 ± 9.1, 74 | 246.7 ± 10.3, 66 | 246.4 ± 10.2, 81 | 246.2 ± 16.5, 22 | 249.0 ± 7.0, 36 | 194.4 ± 30.0, 18 |
| Scotopic a amplitude, 10 cd.s/m2 | µV | 176.3 ± 62.3, 143 | 135.7 ± 49.2, 403 | 114.9 ± 42.1, 153 | 91.0 ± 35.3, 50 | 88.6 ± 30.9, 78 | 45.1 ± 24.2, 31 |
| Scotopic b amplitude, 10 cd.s/m2 | µV | 389.0 ± 127.2, 143 | 321.0 ± 115.3, 403 | 292.2 ± 99.4, 153 | 236.9 ± 86.5, 50 | 231.6 ± 89, 78 | 130.3 ± 62.5, 31 |
| Photopic a amplitude, 100 cd.s/m2 | µV | 20.8 ± 7.3, 88 | 16.3 ± 5.1, 358 | 14.6 ± 4.6, 136 | 12.4 ± 3.4, 46 | 11.0 ± 3.5, 64 | 8.8 ± 3.9, 20 |
| Photopic b amplitude, 100 cd.s/m2 | µV | 164.2 ± 58.9, 88 | 133.2 ± 45.2, 358 | 123.0 ± 40.2, 136 | 110.1 ± 39.8, 46 | 98.3 ± 37.7, 64 | 58.7 ± 33.7, 20 |
| Scotopic a implicit time, 10 cd.s/m2 | ms | 14.2 ± 1.7, 143 | 14.8 ± 1.8, 403 | 15.2 ± 1.9, 153 | 15.3 ± 2.5, 50 | 15.8 ± 2.7, 78 | 18.4 ± 3.0, 31 |
| Scotopic b implicit time, 10 cd.s/m2 | ms | 37.3 ± 4.9, 143 | 35.5 ± 4.5, 403 | 34.7 ± 2.5, 153 | 33.1 ± 2.8, 50 | 33.5 ± 3.4, 78 | 34.9 ± 3.0, 31 |
| Photopic a implicit time, 100 cd.s/m2 | ms | 14.6 ± 2.0, 88 | 13.4 ± 2.2, 358 | 13.4 ± 2.0, 136 | 12.8 ± 3.3, 46 | 12.4 ± 2.1, 64 | 13.7 ± 5.3, 20 |
| Photopic b implicit time, 100 cd.s/m2 | ms | 45.2 ± 7.9, 88 | 41.0 ± 4.4, 358 | 40.6 ± 3.8, 136 | 37.1 ± 3.2, 46 | 38.3 ± 3.2, 64 | 42.4 ± 7.8, 20 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lindovsky, J.; Palkova, M.; Symkina, V.; Raishbrook, M.J.; Prochazka, J.; Sedlacek, R. OCT and ERG Techniques in High-Throughput Phenotyping of Mouse Vision. Genes 2023, 14, 294. https://doi.org/10.3390/genes14020294
Lindovsky J, Palkova M, Symkina V, Raishbrook MJ, Prochazka J, Sedlacek R. OCT and ERG Techniques in High-Throughput Phenotyping of Mouse Vision. Genes. 2023; 14(2):294. https://doi.org/10.3390/genes14020294
Chicago/Turabian StyleLindovsky, Jiri, Marcela Palkova, Viktoriia Symkina, Miles Joseph Raishbrook, Jan Prochazka, and Radislav Sedlacek. 2023. "OCT and ERG Techniques in High-Throughput Phenotyping of Mouse Vision" Genes 14, no. 2: 294. https://doi.org/10.3390/genes14020294
APA StyleLindovsky, J., Palkova, M., Symkina, V., Raishbrook, M. J., Prochazka, J., & Sedlacek, R. (2023). OCT and ERG Techniques in High-Throughput Phenotyping of Mouse Vision. Genes, 14(2), 294. https://doi.org/10.3390/genes14020294