
Identification of the BcLEA Gene Family and Functional Analysis of the BcLEA73 Gene in Wucai (Brassica campestris L.)

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Identification of the BrLEA Gene Family Members and Analysis of Physicochemical Properties
2.3. Analysis of Conserved Motifs and Gene Structure of the BrLEA Gene Family Members
2.4. Chromosome Location Analysis of the BrLEA Gene FAMILY
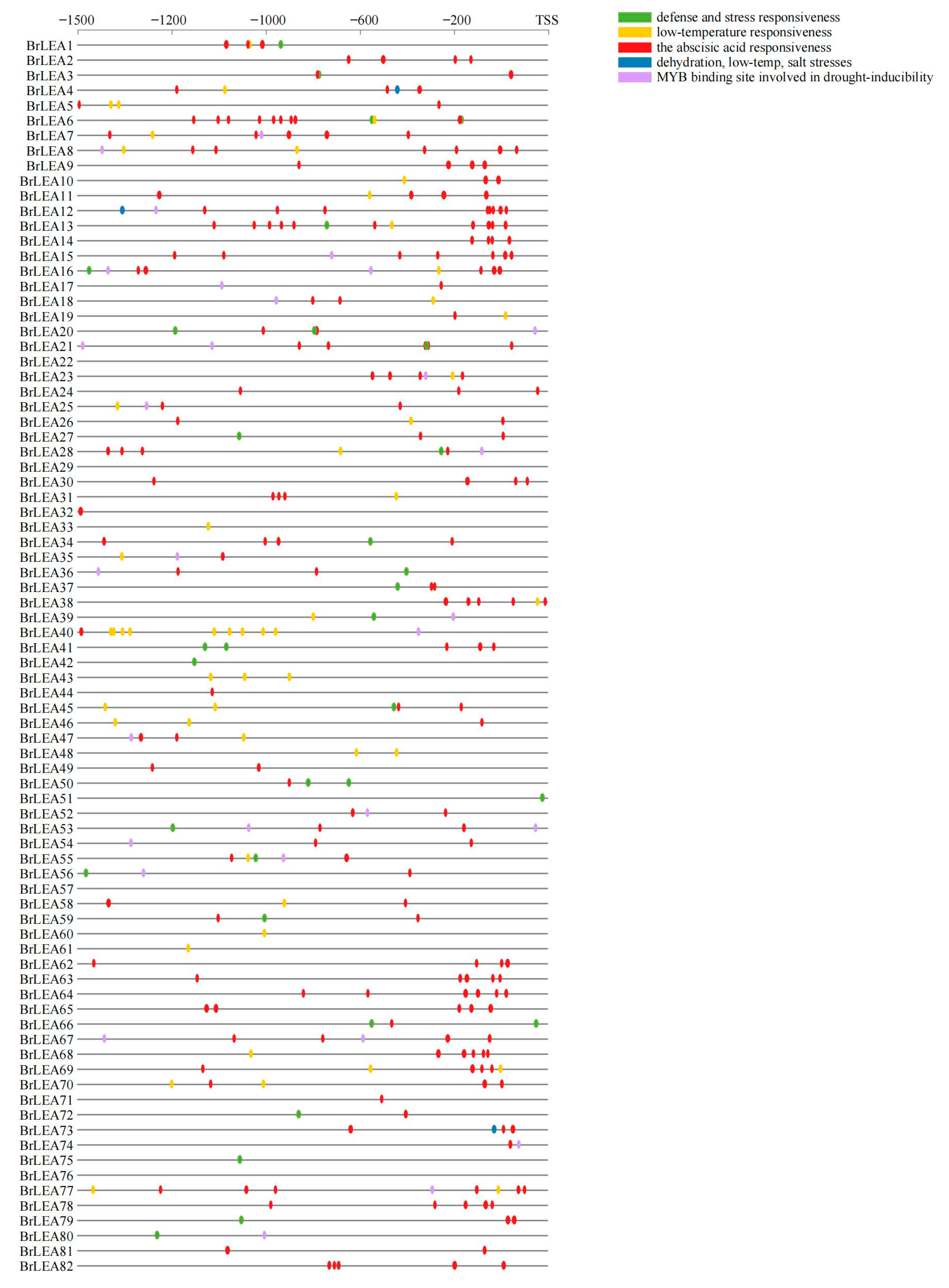
2.5. Cis-Acting Element Analysis of the BrLEA Gene Family
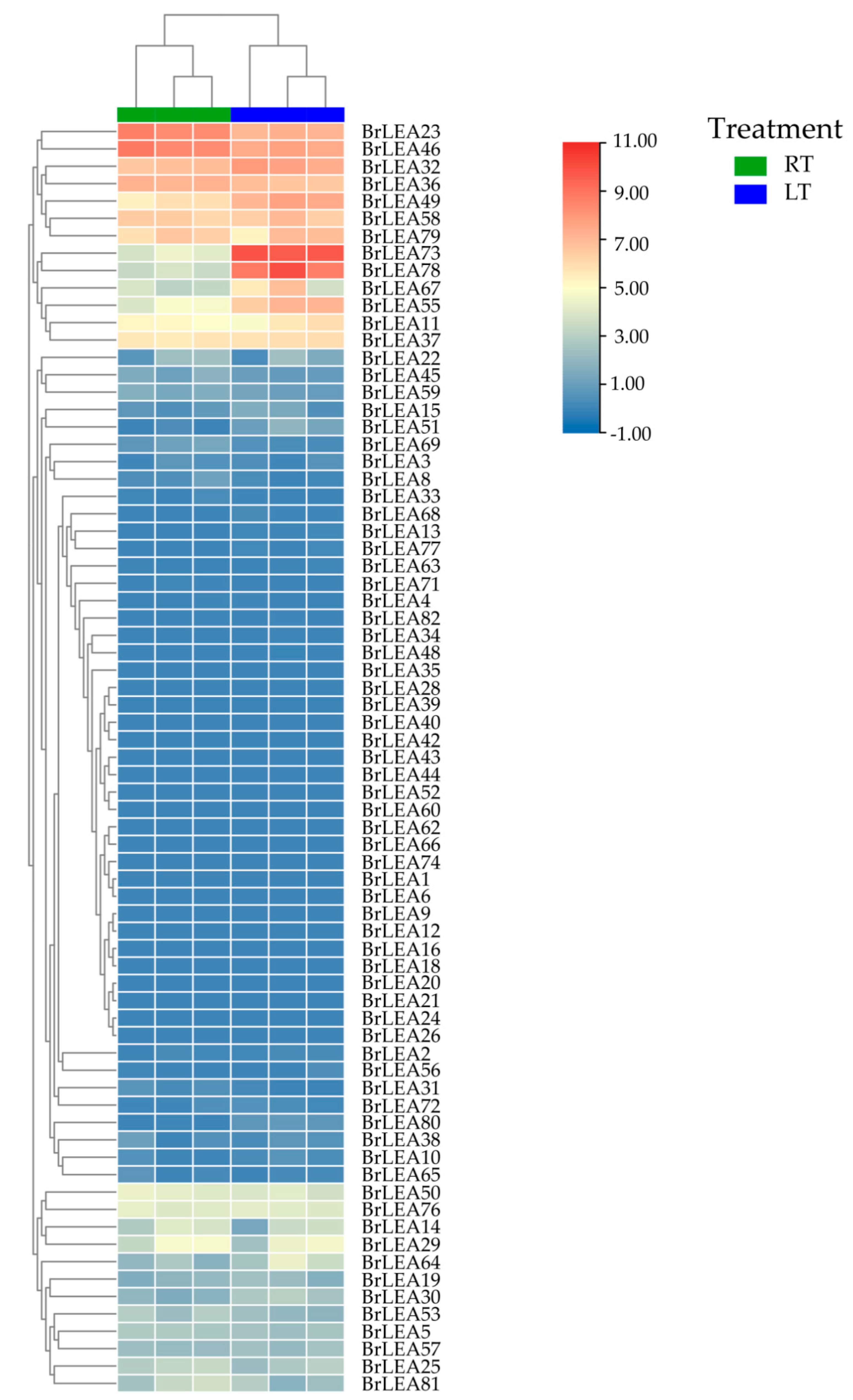
2.6. Screening of Differentially Expressed BcLEAs Based on RNA-Seq
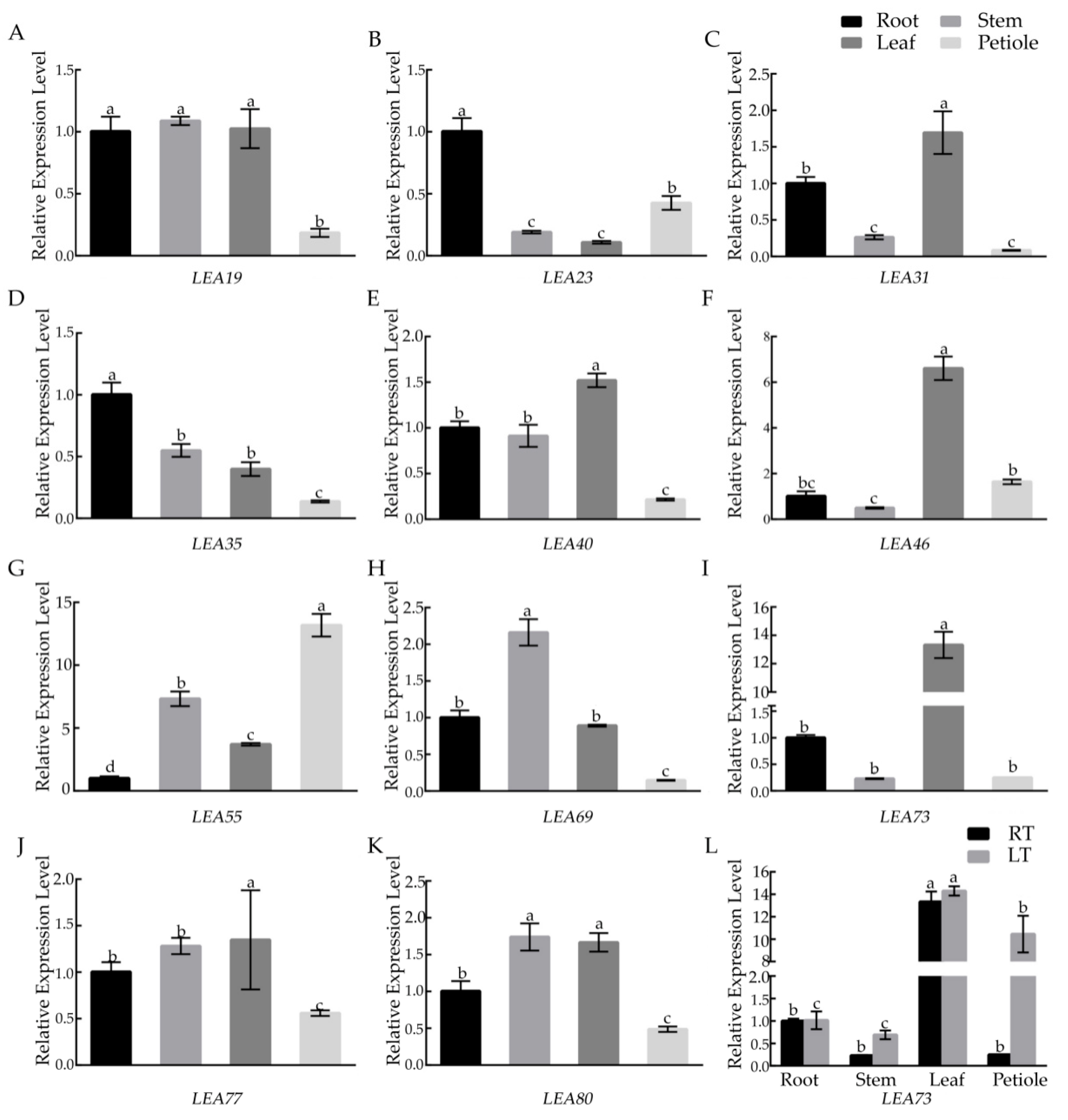
2.7. Analysis of the BcLEA Genes Expression Patterns in Wucai
2.8. Cloning of the BcLEA73 Gene of Wucai
2.9. Subcellular Localization Analysis of the BcLEA73 Gene in Wucai
2.10. Genetic Transformation Analysis of the BcLEA73 Gene in Arabidopsis
2.11. Analysis of Abiotic Stress on Seed Germination Rate and Root Length
2.12. Measurement of O2− Production Rate, H2O2 Content, Relative Electrolyte Leakage (REL), and Total Antioxidant Capacity (T-AOC)
3. Results
3.1. Bioinformatics Analysis of the BrLEA Gene Family
3.1.1. Identification of the BrLEA Gene Family Members and Analysis of Physicochemical Properties
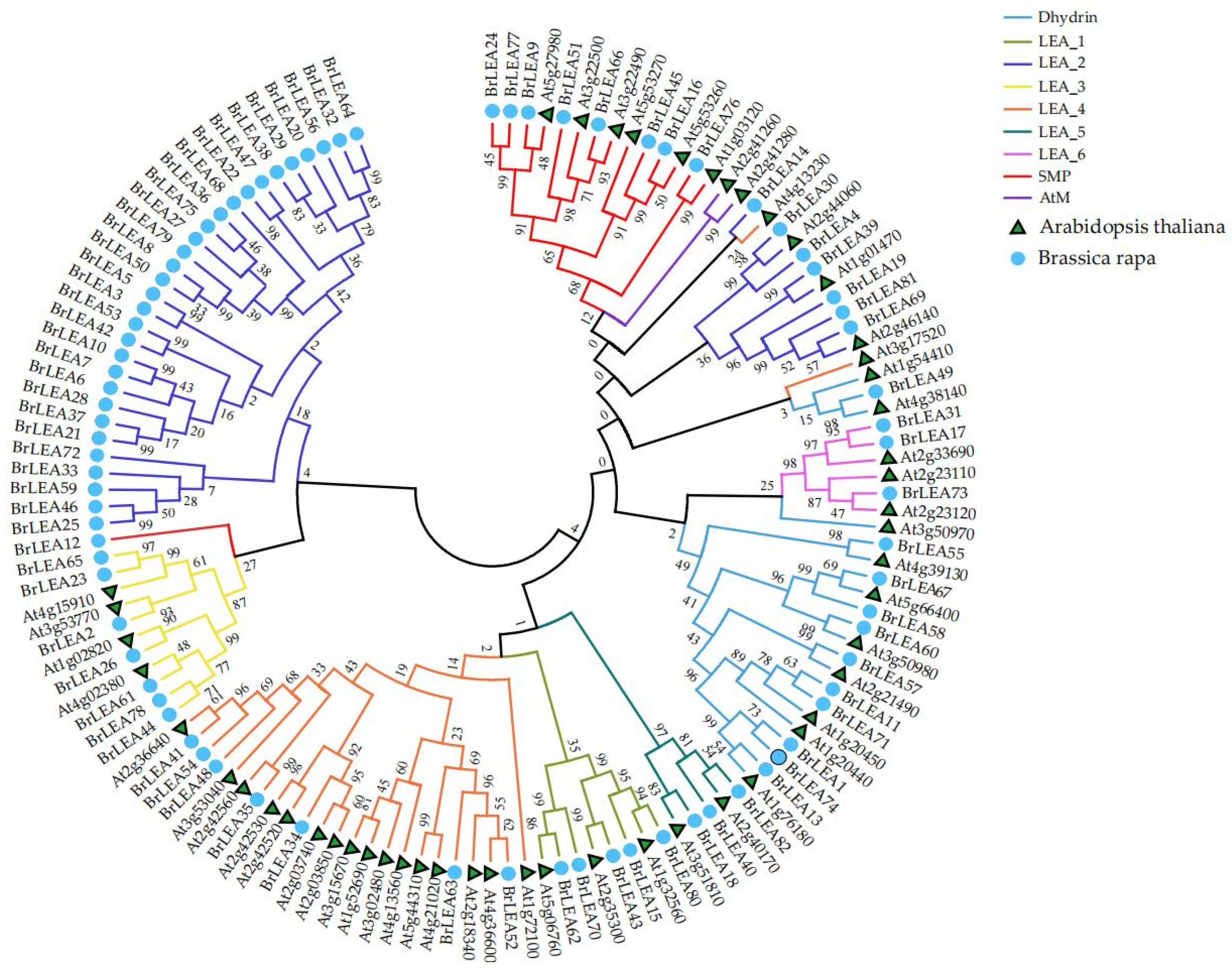
3.1.2. Construction of the Gene Family Evolutionary Tree
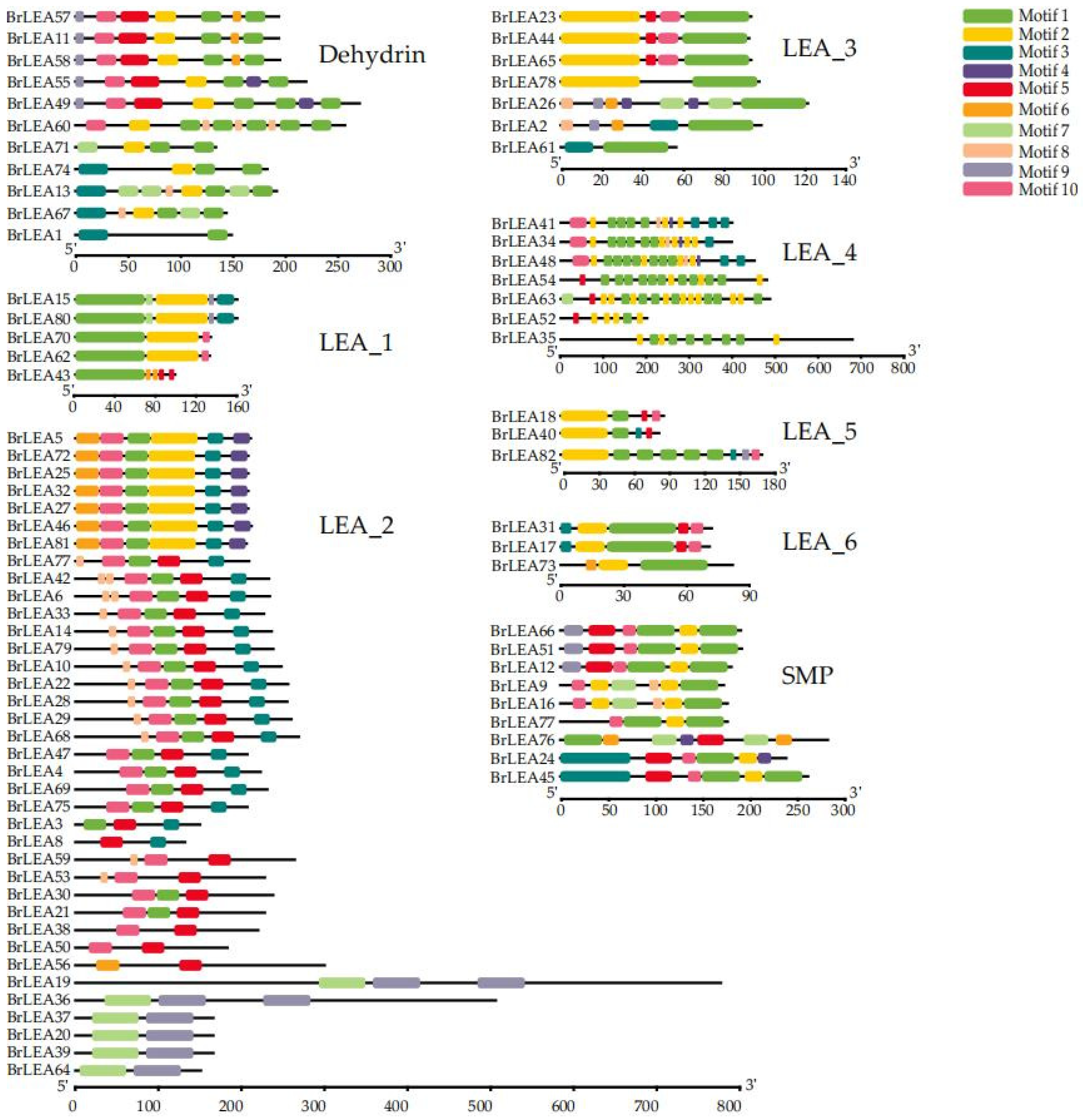
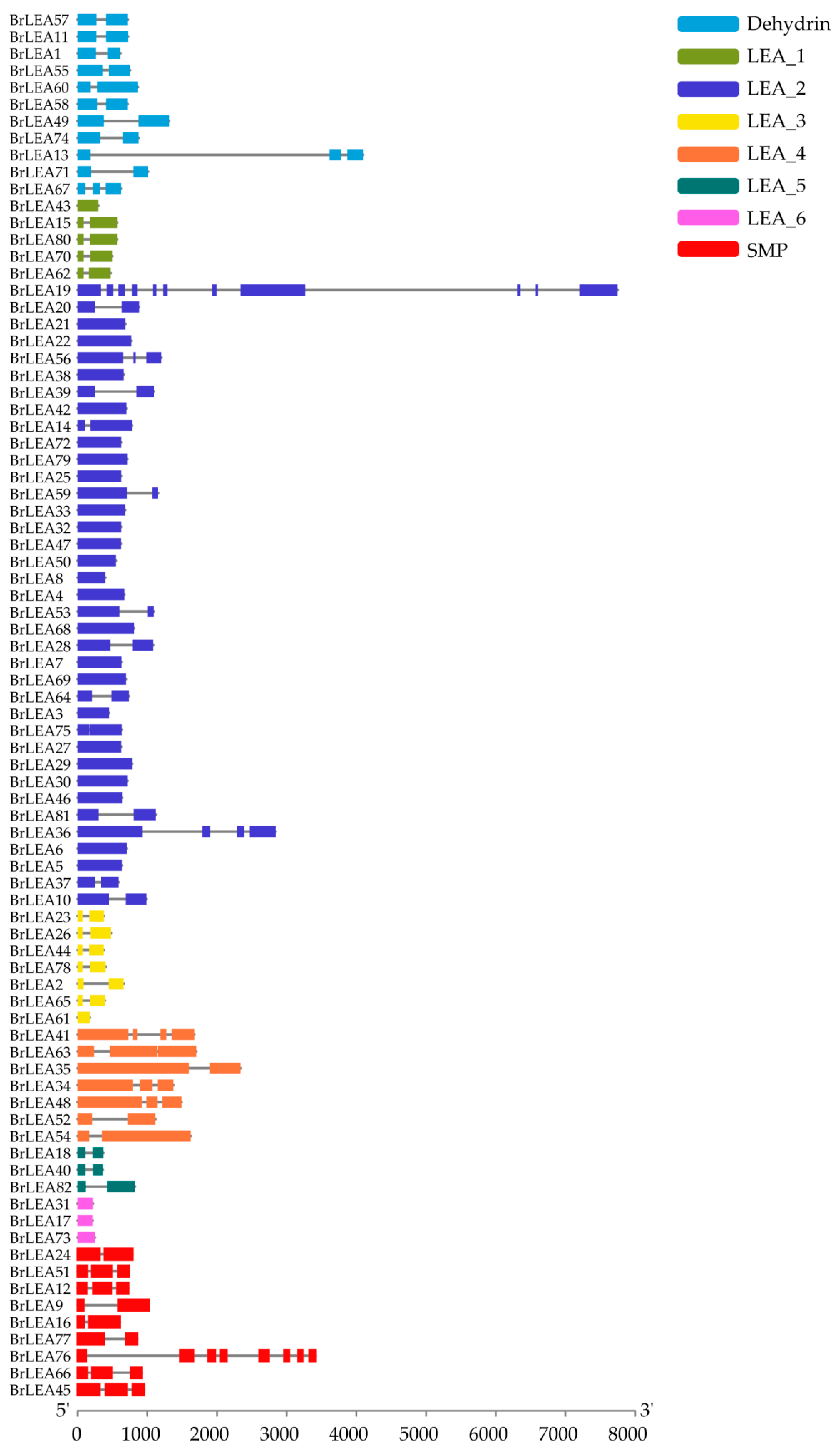
3.1.3. Analysis of the Conserved Motifs and Gene Structure of the Family Members
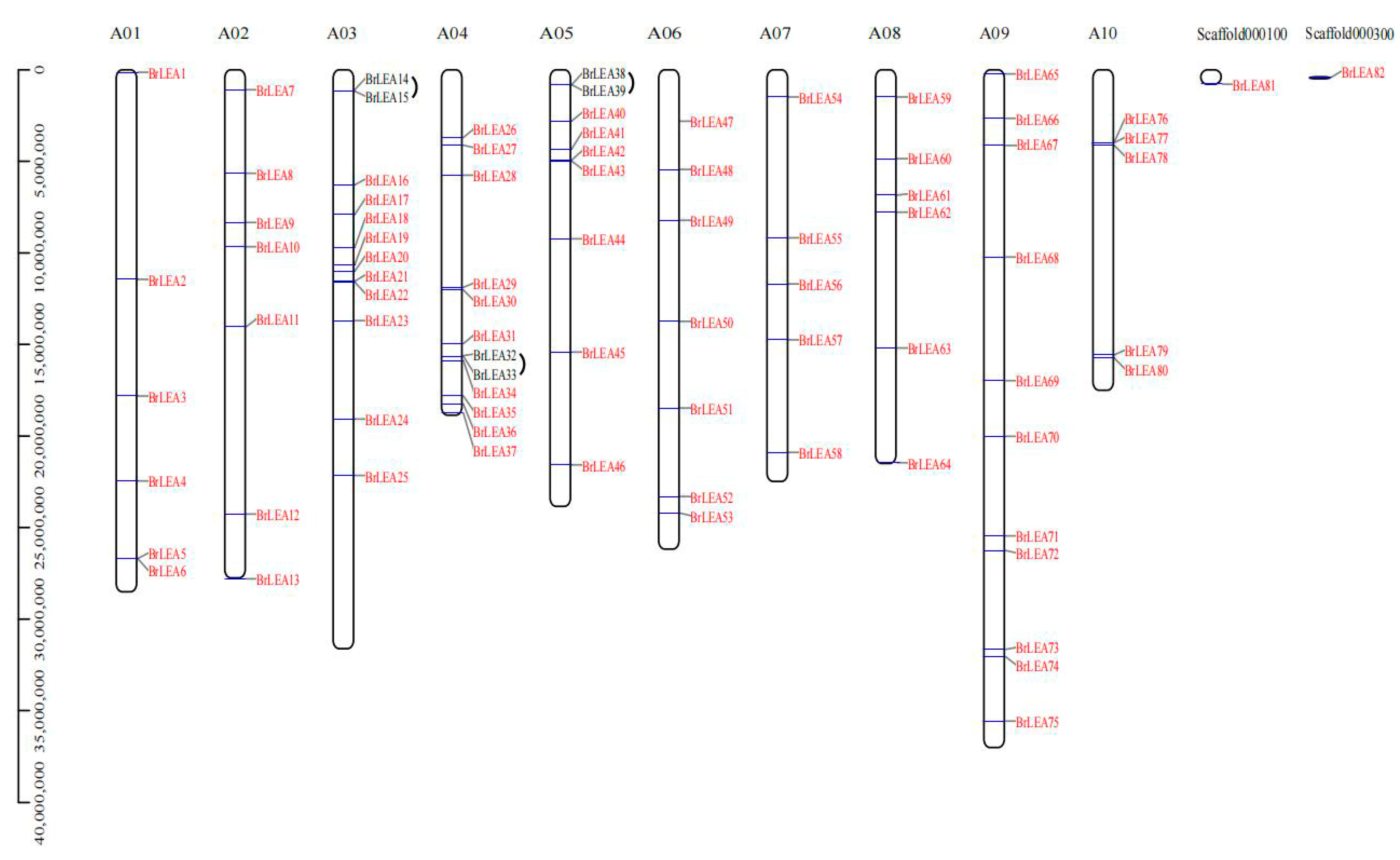
3.1.4. Chromosome Location Analysis
3.1.5. Cis-Acting Element Analysis
3.2. Analysis of the BcLEA Gene Transcriptome Data of Wucai
3.3. Analysis of BcLEA Gene Expression Patterns
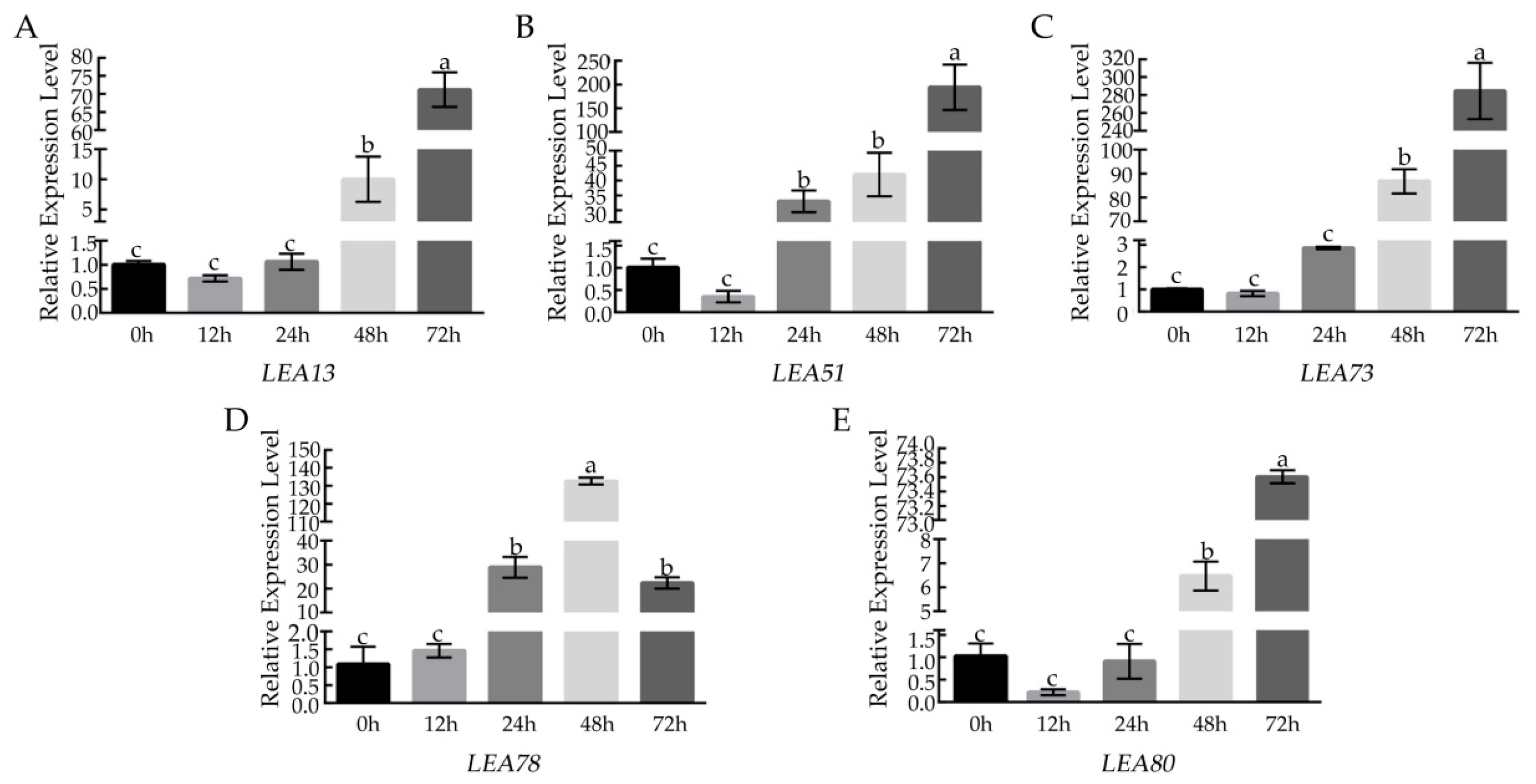
3.4. Analysis of BcLEA Gene Expression under Low-Temperature Treatment
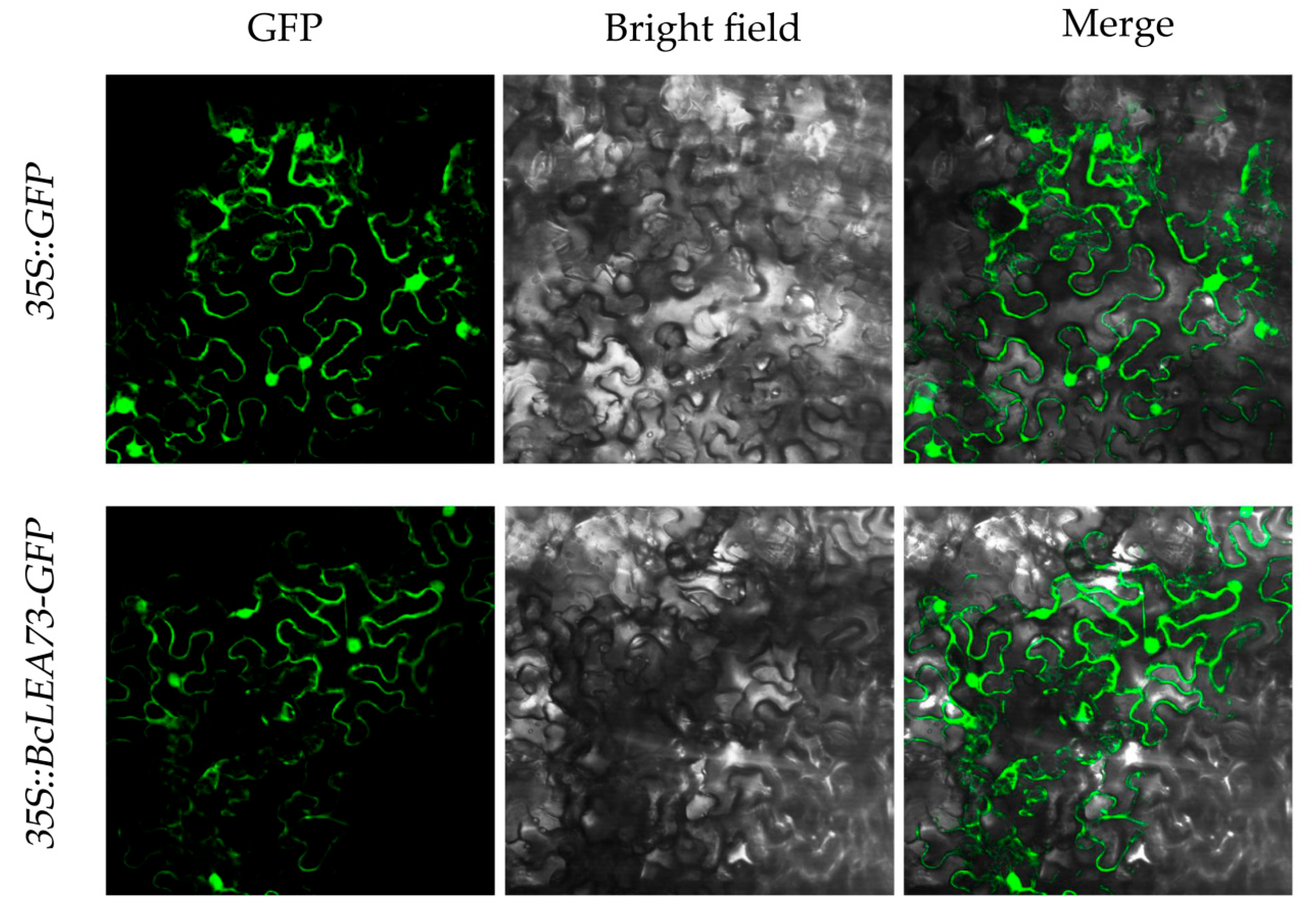
3.5. Subcellular Localization Analysis of BcLEA73 in Wucai
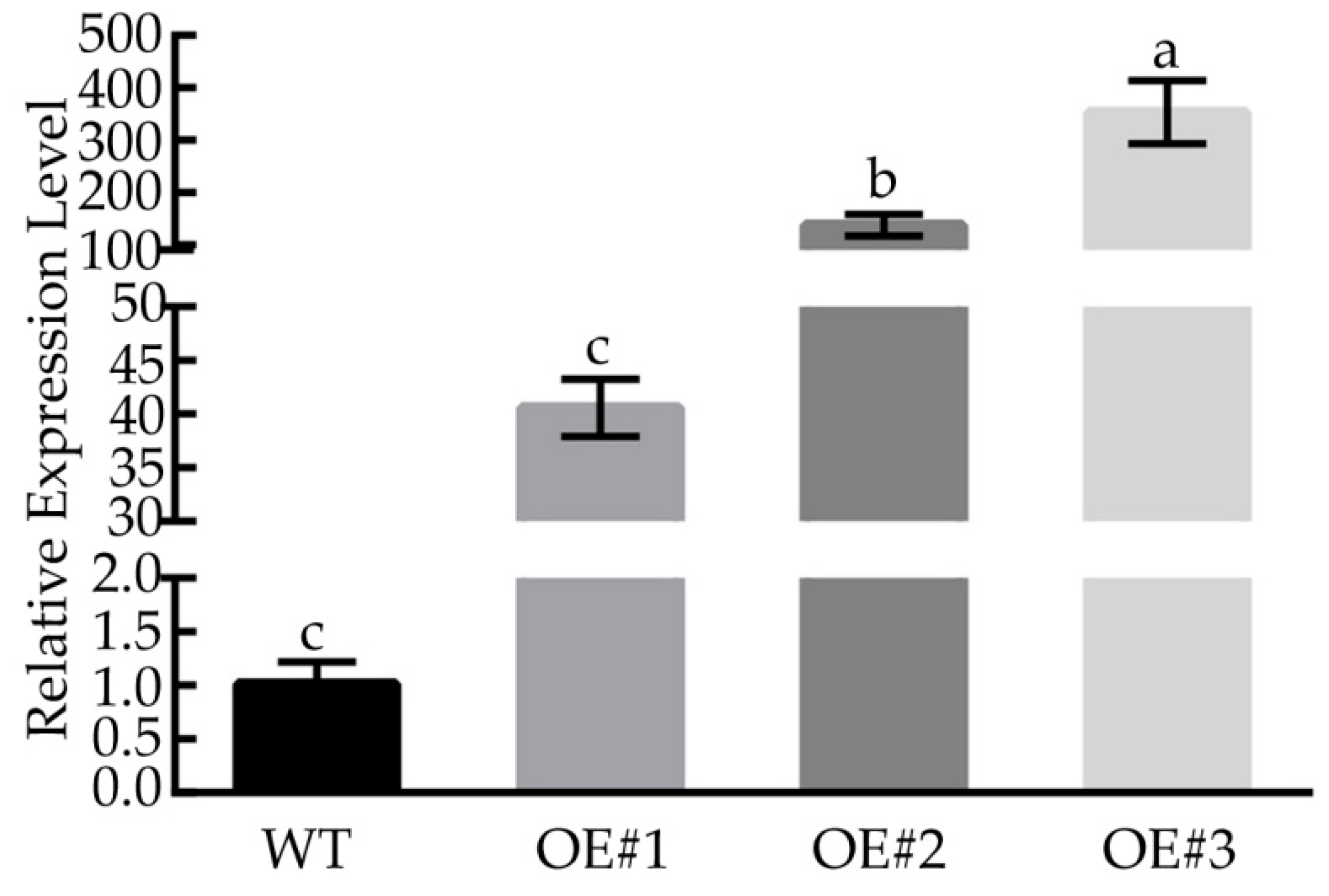
3.6. Overexpression of BcLEA73 in Arabidopsis Increases Abiotic Stress Tolerance
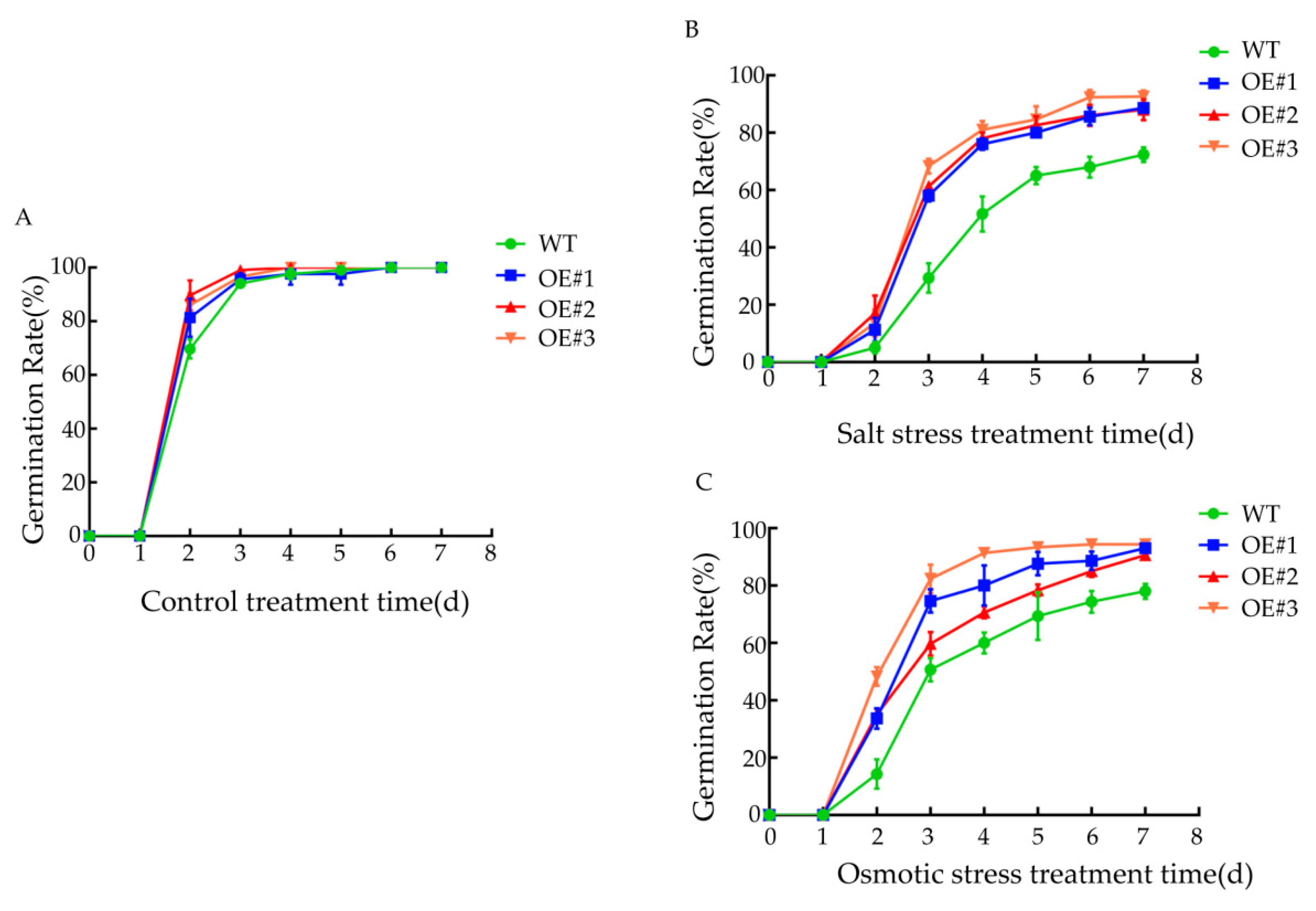
3.7. Effects of Various Stress Treatments on Seed Germination Rate
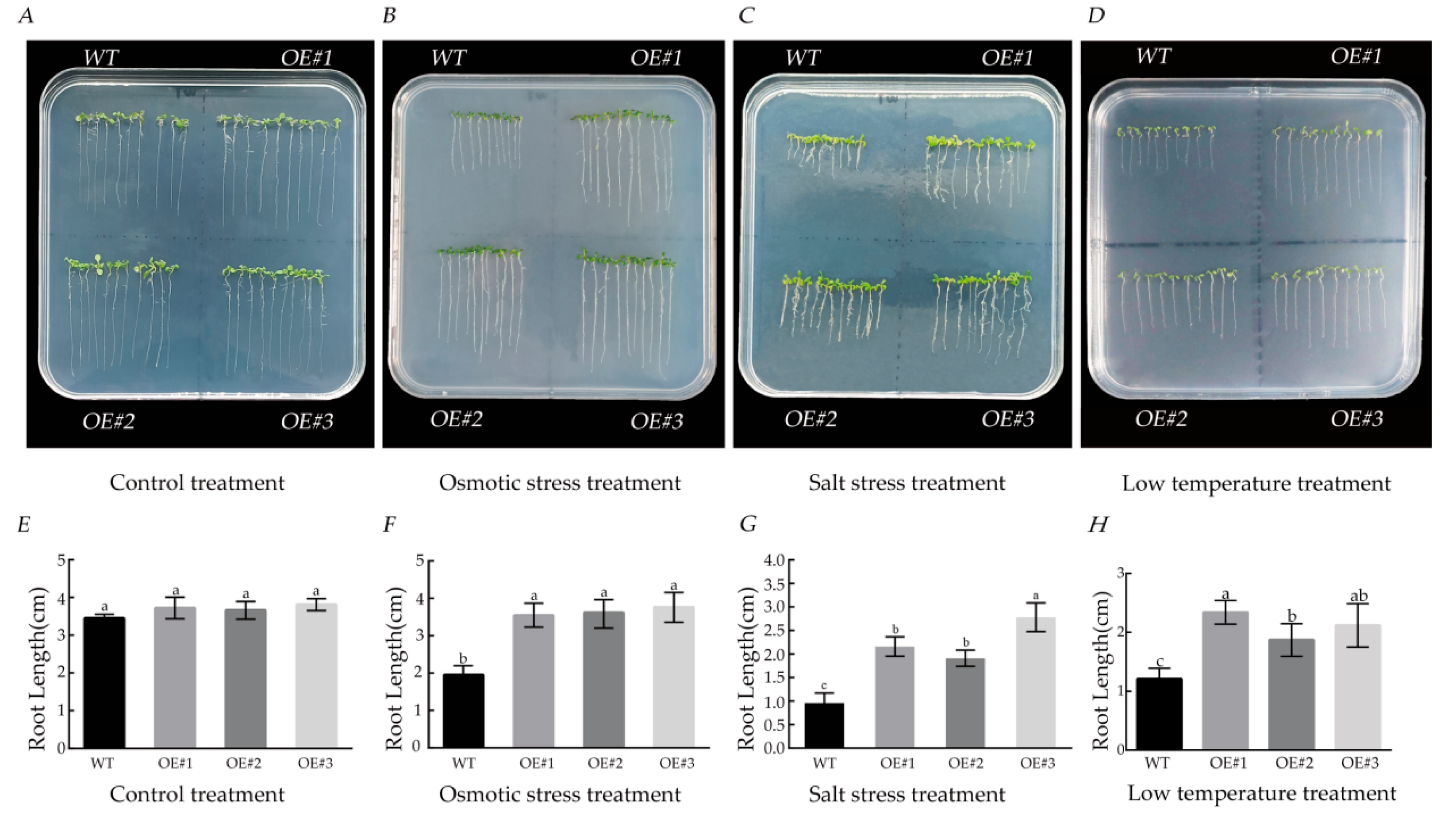
3.8. Effects of Different Stress Treatments on Root Length
3.9. Effect of Drought Stress on Seedling Survival Rate
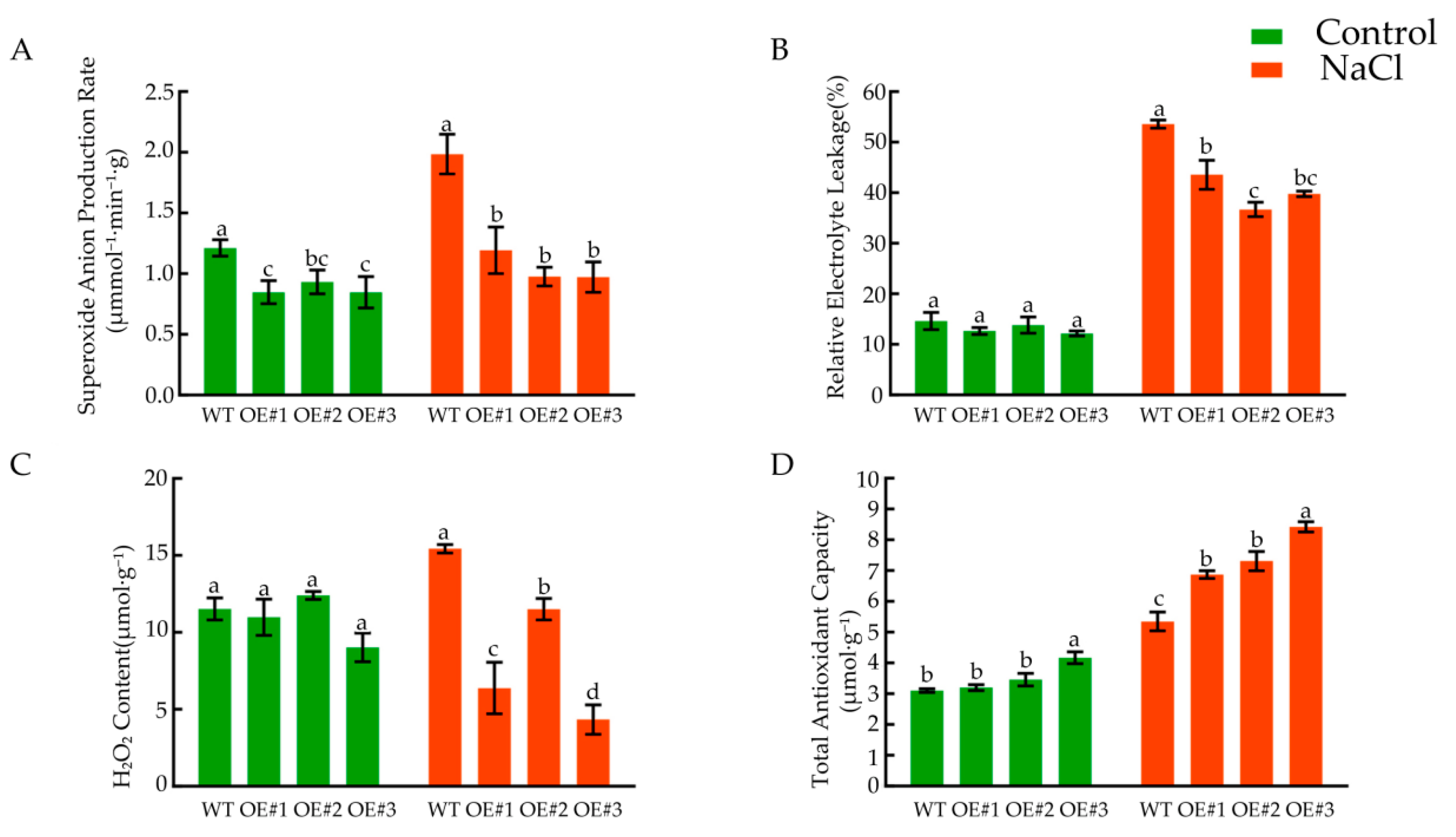
3.10. Effects of Salt Stress on the Physiological Indices of the Seedlings
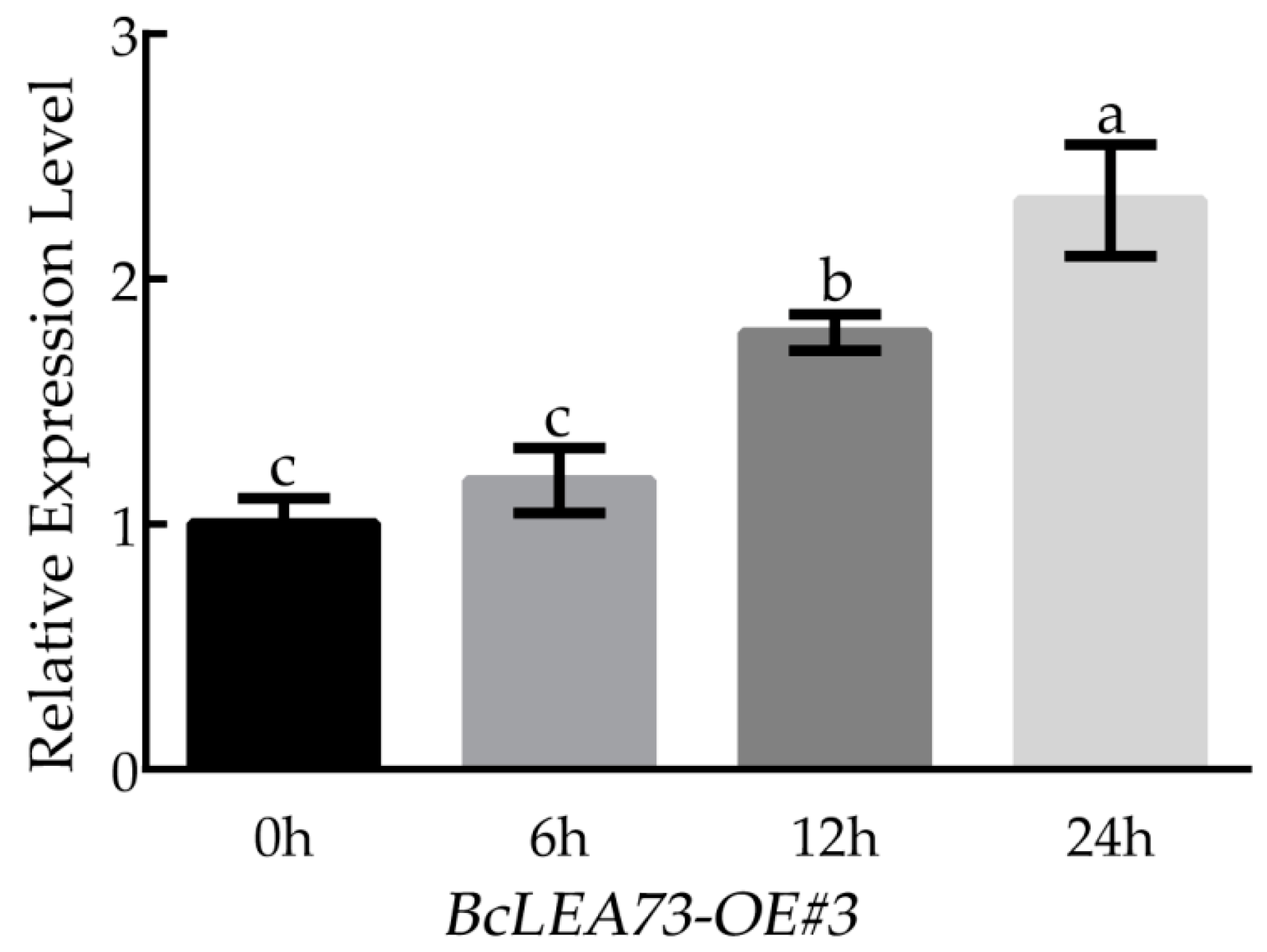
3.11. The Expression of BcLEA73 in Arabidopsis under Low-Temperature Treatment
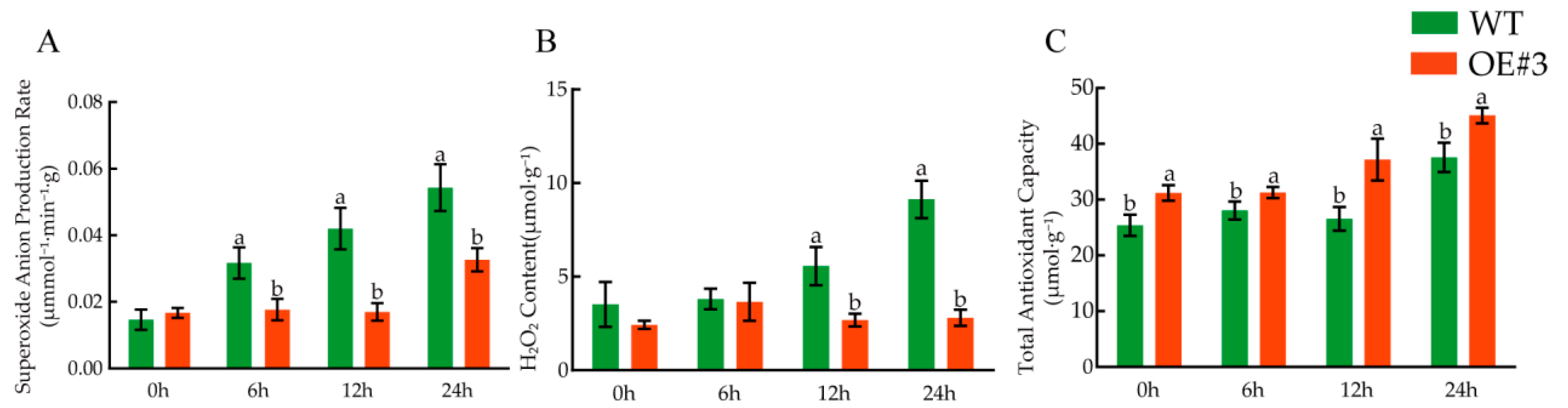
3.12. Determination of Related Physiological Indices under Low-Temperature Treatment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Dure, L., 3rd; Greenway, S.C.; Galau, G.A. Developmental biochemistry of cottonseed embryogenesis and germination: Changing messenger ribonucleic acid populations as shown by in vitro and in vivo protein synthesis. Biochemistry 1981, 20, 4162–4168. [Google Scholar] [CrossRef] [PubMed]
- Hara, M.; Terashima, S.; Kuboi, T. Characterization and cryoprotective activity of cold-responsive dehydrin from Citrus unshiu. J. Plant Physiol. 2001, 158, 1333–1339. [Google Scholar] [CrossRef]
- Bravo, L.A.; Gallardo, J.; Navarrete, A.; Olave, N.; Martínez, J.; Alberdi, M.; Close, T.J.; Corcuera, L.J. Cryoprotective activity of a cold-induced dehydrin purified from barley. Physiol. Plant. 2003, 118, 262–269. [Google Scholar] [CrossRef]
- Lee, S.C.; Luan, S. ABA signal transduction at the crossroad of biotic and abiotic stress responses. Plant Cell Environ. 2011, 35, 53–60. [Google Scholar] [CrossRef]
- Bao, F.; Du, D.; An, Y.; Yang, W.; Wang, J.; Cheng, T.; Zhang, Q. Overexpression of Prunus mume Dehydrin Genes in Tobacco Enhances Tolerance to Cold and Drought. Front. Plant Sci. 2017, 8, 151. [Google Scholar] [CrossRef]
- Skriver, K.; Mundy, J. Gene expression in response to abscisic acid and osmotic stress. Plant Cell 1990, 2, 503–512. [Google Scholar]
- Wang, X.; Wang, H.; Wang, J.; Sun, R.; Wu, J.; Liu, S.; Bai, Y.; Mun, J.-H.; Bancroft, I.; Cheng, F.; et al. The genome of the mesopolyploid crop species Brassica rapa. Nat. Genet. 2011, 43, 1035–1039. [Google Scholar] [CrossRef]
- Finn, R.D.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Mistry, J.; Mitchell, A.L.; Potter, S.C.; Punta, M.; Qureshi, M.; Sangrador-Vegas, A.; et al. The Pfam protein families database: Towards a more sustainable future. Nucleic Acids Res. 2016, 44, D279–D285. [Google Scholar] [CrossRef]
- Letunic, I.; Doerks, T.; Bork, P. SMART: Recent updates, new developments and status in 2015. Nucleic Acids Res. 2015, 43, D257–D260. [Google Scholar] [CrossRef]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, w202–w208. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; Van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, M.; Zhou, J.; Gao, X.; Zhu, S.; Yuan, L.; Hou, X.; Liu, T.; Chen, G.; Tang, X.; et al. Transcriptome analysis and differential gene expression profiling of wucai (Brassica campestris L.) in response to cold stress. BMC Genom. 2022, 23, 137. [Google Scholar] [CrossRef]
- Hundertmark, M.; Hincha, D.K. LEA (Late Embryogenesis Abundant) proteins and their encoding genes in Arabidopsis thaliana. BMC Genom. 2008, 9, 118. [Google Scholar] [CrossRef]
- Cao, J.; Shi, F.; Liu, X.; Huang, G.; Zhou, M. Phylogenetic analysis and evolution of aromatic amino acid hydroxylase. FEBS Lett. 2010, 584, 4775–4782. [Google Scholar] [CrossRef]
- Cao, J.; Huang, J.; Yang, Y.; Hu, X. Analyses of the oligopeptide transporter gene family in poplar and grape. BMC Genom. 2011, 12, 465. [Google Scholar] [CrossRef]
- Cao, J.; Shi, F. Evolution of the RALF Gene Family in Plants: Gene Duplication and Selection Patterns. Evol. Bioinform. 2012, 8, 271–292. [Google Scholar] [CrossRef]
- Altunoglu, Y.C.; Baloglu, M.C.; Baloglu, P.; Yer, E.N.; Kara, S. Genome-wide identification and comparative expression analysis of LEA genes in watermelon and melon genomes. Physiol. Mol. Biol. Plants 2017, 23, 5–21. [Google Scholar] [CrossRef]
- Ding, M.; Wang, L.; Zhan, W.; Sun, G.; Jia, X.; Chen, S.; Ding, W.; Yang, J. Genome-wide identification and expression analysis of late embryogenesis abundant protein-encoding genes in rye (Secale cereale L.). PLoS ONE 2021, 16, e0249757. [Google Scholar] [CrossRef]
- Ibrahime, M.; Kibar, U.; Kazan, K.; Özmen, C.Y.; Mutaf, F.; Aşçı, S.D.; Aydemir, B.; Ergül, A. Genome-wide identification of the LEA protein gene family in grapevine (Vitis vinifera L.). Tree Genet. Genomes 2019, 15, 55. [Google Scholar] [CrossRef]
- Li, X.; Cao, J. Late Embryogenesis Abundant (LEA) Gene Family in Maize: Identification, Evolution, and Expression Profiles. Plant Mol. Biol. Rep. 2015, 34, 15–28. [Google Scholar] [CrossRef]
- Zan, T.; Li, L.; Li, J.; Zhang, L.; Li, X. Genome-wide identification and characterization of late embryogenesis abundant protein-encoding gene family in wheat: Evolution and expression profiles during development and stress. Gene 2020, 736, 144422. [Google Scholar] [CrossRef] [PubMed]
- Hua, L. Cloning and function analysis of CkLEA4-1 and CkLEA6-1 from Caragana korshinskii Kom. 2017. Available online: https://kns.cnki.net/KCMS/detail/detail.aspx?dbname=CMFD201801&filename=1017211776.nh (accessed on 18 January 2023). (In Chinese).
- Ding, M. The Identification of LEA Genes Family in Secale cereale and Function Exploration of LEA19 in Arabidopsis. 2021. Available online: https://kns.cnki.net/KCMS/detail/detail.aspx?dbname=CMFD202201&filename=1021056379.nh (accessed on 18 January 2023). (In Chinese).
- Liang, Y.; Xiong, Z.; Zheng, J.; Xu, D.; Zhu, Z.; Xiang, J.; Gan, J.; Raboanatahiry, N.; Yin, Y.; Li, M. Genome-wide identification, structural analysis and new insights into late embryogenesis abundant (LEA) gene family formation pattern in Brassica napus. Sci. Rep. 2016, 6, 24265. [Google Scholar] [CrossRef]
- Pedrosa, A.M.; Martins, C.D.P.S.; Gonçalves, L.P.; Costa, M.G.C. Late Embryogenesis Abundant (LEA) Constitutes a Large and Diverse Family of Proteins Involved in Development and Abiotic Stress Responses in Sweet Orange (Citrus sinensis L. Osb.). PLoS ONE 2015, 10, e0145785. [Google Scholar] [CrossRef]
- Xie, D.W.; Wang, X.N.; Fu, L.S.; Sun, J.; Zheng, W.; Li, Z.F. Identification of the trehalose-6-phosphate synthase gene family in winter wheat and expression analysis under conditions of freezing stress. J. Genet. 2015, 94, 55–65. [Google Scholar] [CrossRef]
- Chen, Y.; Hao, X.; Cao, J. Small auxin upregulated RNA (SAUR) gene family in maize: Identification, evolution, and its phylogenetic comparison with Arabidopsis, rice, and sorghum. J. Integr. Plant Biol. 2014, 56, 133–150. [Google Scholar] [CrossRef]
- Cao, J.; Li, X. Identification and phylogenetic analysis of late embryogenesis abundant proteins family in tomato (Solanum lycopersicum). Planta 2014, 241, 757–772. [Google Scholar] [CrossRef]
- Nagaraju, M.; Kumar, S.A.; Reddy, P.S.; Kumar, A.; Rao, D.M.; Kishor, P.B.K. Genome-scale identification, classification, and tissue specific expression analysis of late embryogenesis abundant (LEA) genes under abiotic stress conditions in Sorghum bicolor L. PLoS ONE 2019, 14, e0209980. [Google Scholar] [CrossRef]
- Shu, Y.; Liu, Y.; Zhang, J.; Song, L.; Guo, C. Genome-Wide Analysis of the AP2/ERF Superfamily Genes and their Responses to Abiotic Stress in Medicago truncatula. Front. Plant Sci. 2016, 6, 1247. [Google Scholar] [CrossRef]
- Liu, H. Genome-wide Identification of Late Embryogenesis Abundant (LEA) GeneFamily and Regulatory Mechanism of Dehydrin WZY2 in Response to Drought Stress in Wheat. 2021. Available online: https://kns.cnki.net/KCMS/detail/detail.aspx?dbname=CDFDLAST2022&filename=1021711649.nh (accessed on 18 January 2023). (In Chinese).
- Zheng, J.; Su, H.; Lin, R.; Zhang, H.; Xia, K.; Jian, S.; Zhang, M. Isolation and characterization of an atypical LEA gene (IpLEA) from Ipomoea pes-caprae conferring salt/drought and oxidative stress tolerance. Sci. Rep. 2019, 9, 14838. [Google Scholar] [CrossRef]
- Wang, M.; Li, P.; Li, C.; Pan, Y.; Jiang, X.; Zhu, D.; Zhao, Q.; Yu, J. SiLEA14, a novel atypical LEA protein, confers abiotic stress resistance in foxtail millet. BMC Plant Biol. 2014, 14, 290. [Google Scholar] [CrossRef]
- Olvera-Carrillo, Y.; Campos, F.; Reyes, J.L.; Garciarrubio, A.; Covarrubias, A.A. Functional analysis of the group 4 late embryogenesis abundant proteins reveals their relevance in the adaptive response during water deficit in Arabidopsis. Plant Physiol. 2010, 154, 373–390. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, L.; Xing, X.; Sun, L.; Pan, J.; Kong, X.; Zhang, M.; Li, D. ZmLEA3, a multifunctional group 3 LEA protein from maize (Zea mays L.), is involved in biotic and abiotic stresses. Plant Cell Physiol. 2013, 54, 944–959. [Google Scholar] [CrossRef]
- Xie, C.; Zhang, R.; Qu, Y.; Miao, Z.; Zhang, Y.; Shen, X.; Wang, T.; Dong, J. Overexpression of MtCAS31 enhances drought tolerance in transgenic Arabidopsis by reducing stomatal density. New Phytol. 2012, 195, 124–135. [Google Scholar] [CrossRef]
- Hara, M.; Fujinaga, M.; Kuboi, T. Radical scavenging activity and oxidative modification of citrus dehydrin. Plant Physiol. Biochem. 2004, 42, 657–662. [Google Scholar] [CrossRef]
- Dean, R.T.; Fu, S.; Stocker, R.; Davies, M.J. Biochemistry and pathology of radical-mediated protein oxidation. Biochem. J. 1997, 324, 1–18. [Google Scholar] [CrossRef]
- Xing, X.; Liu, Y.; Kong, X.; Liu, Y.; Li, D. Overexpression of a maize dehydrin gene, ZmDHN2b, in tobacco enhances tolerance to low temperature. Plant Growth Regul. 2011, 65, 109–118. [Google Scholar] [CrossRef]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence (5’—3’) |
|---|---|
| RT-BcActin (F) | TGGGTTTGCTGGTGACGAT |
| RT-BcActin (R) | TGCCTAGGACGACCAACAATACT |
| RT-AtActin (F) | GACCTTTAACTCTCCCGCTA |
| RT-AtActin (R) | GGAAGAGAGAAACCCTCGTA |
| RT-BrLEA13 (F) | CGAAGGATACGGGACAGGAAC |
| RT-BrLEA13 (R) | GTGAAGCATTCCTCCCAAGCC |
| RT-BrLEA51 (F) | AATTACCGTCAGACAAACCAG |
| RT-BrLEA51 (R) | ACTCCGGTTGGGTAAGTAGTG |
| RT-BrLEA73 (F) | GTGGACGAATCTGGTAGCTTG |
| RT-BrLEA73 (R) | TAGATCCACCACCGAGGCCAG |
| RT-BrLEA78 (F) | TCTCCAACGCCATCTACAGAC |
| RT-BrLEA78(R) | CTCGCTTGACTCTTCCCCAAC |
| RT-BrLEA80 (F) | TGGAGGAGAAGGCTGAGAAGA |
| RT-BrLEA80 (R) | TCCACTGGCTTCTTTCATGAC |
| Primer Name | Primer Sequence (5’—3’) |
|---|---|
| BrLEA73(F) | ATGGAGGCAGAGAAGACAC |
| BrLEA73(R) | TCAAGGAGCTTTCTCAGCAGT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, Y.; Zhang, S.; Xu, H.; Tian, H.; Zhang, M.; Zhu, S.; Wang, C.; Hou, J.; Chen, G.; Tang, X.; et al. Identification of the BcLEA Gene Family and Functional Analysis of the BcLEA73 Gene in Wucai (Brassica campestris L.). Genes 2023, 14, 415. https://doi.org/10.3390/genes14020415
Jiang Y, Zhang S, Xu H, Tian H, Zhang M, Zhu S, Wang C, Hou J, Chen G, Tang X, et al. Identification of the BcLEA Gene Family and Functional Analysis of the BcLEA73 Gene in Wucai (Brassica campestris L.). Genes. 2023; 14(2):415. https://doi.org/10.3390/genes14020415
Chicago/Turabian StyleJiang, Yueyue, Shengnan Zhang, Hongcheng Xu, Hong Tian, Mengyun Zhang, Shidong Zhu, Chenggang Wang, Jinfeng Hou, Guohu Chen, Xiaoyan Tang, and et al. 2023. "Identification of the BcLEA Gene Family and Functional Analysis of the BcLEA73 Gene in Wucai (Brassica campestris L.)" Genes 14, no. 2: 415. https://doi.org/10.3390/genes14020415
APA StyleJiang, Y., Zhang, S., Xu, H., Tian, H., Zhang, M., Zhu, S., Wang, C., Hou, J., Chen, G., Tang, X., Wang, W., Wu, J., Huang, X., Zhang, J., & Yuan, L. (2023). Identification of the BcLEA Gene Family and Functional Analysis of the BcLEA73 Gene in Wucai (Brassica campestris L.). Genes, 14(2), 415. https://doi.org/10.3390/genes14020415