
Competing Endogenous RNA Regulatory Networks of hsa_circ_0126672 in Pathophysiology of Coronary Heart Disease
 , , and
, , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Search for circRNAs Associated with CHD and Identification of Their Target miRNAs
2.2. Search for CHD Differentially Expressed Genes (DEGs)
2.3. Identification of Differentially Expressed miRNA Target Genes (miRNA-DEGs)
2.4. Functional Enrichment Analysis
2.5. Network Illustrations
2.6. miRNA and Target Sequence Homology Study
2.7. miRNA and Small Molecules Interaction
3. Results
3.1. Identification of Differentially Expressed Genes and Selected Circular RNAs
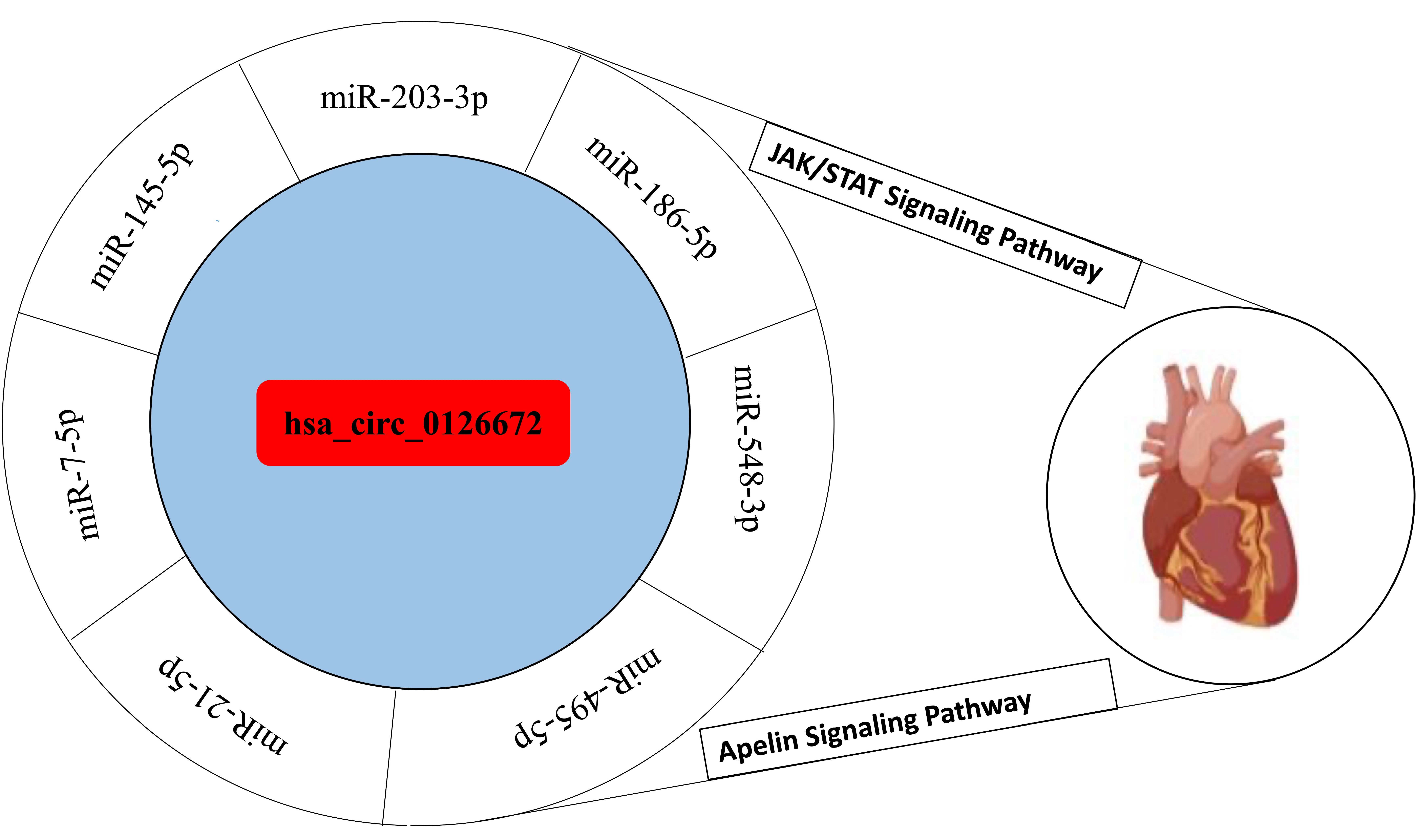
3.2. Interaction of Circular RNA with the Selected miRNAs
3.3. Gene Ontology and KEEG Pathway Analysis
3.4. Circular RNA Networks
3.5. Homology Study and miRNAs Interaction with Small Molecules
3.6. Interaction between miRNAs and Small Molecules
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wong, N.D. Epidemiological Studies of CHD and the Evolution of Preventive Cardiology. Nat. Rev. Cardiol. 2014, 11, 276–289. [Google Scholar] [CrossRef] [PubMed]
- Gurha, P. Noncoding RNAs in Cardiovascular Diseases. Curr. Opin. Cardiol. 2019, 34, 241–245. [Google Scholar] [CrossRef] [PubMed]
- Barolia, R.; Sayani, A.H. Risk Factors of Cardiovascular Disease and Its Recommendations in Pakistani Context. J. Pak. Med. Assoc. 2017, 67, 1723–1729. [Google Scholar]
- Ahmed, S.T.; Rehman, H.; Akeroyd, J.M.; Alam, M.; Shah, T.; Kalra, A.; Virani, S.S. Premature Coronary Heart Disease in South Asians: Burden and Determinants. Curr. Atheroscler. Rep. 2018, 20, 6–13. [Google Scholar] [CrossRef]
- Khera, A.V.; Emdin, C.A.; Drake, I.; Natarajan, P.; Bick, A.G.; Cook, N.R.; Chasman, D.I.; Baber, U.; Mehran, R.; Rader, D.J.; et al. Genetic Risk, Adherence to a Healthy Lifestyle, and Coronary Disease. N. Engl. J. Med. 2016, 375, 2349–2358. [Google Scholar] [CrossRef] [Green Version]
- Roberts, R. A Genetic Basis for Coronary Artery Disease. Trends Cardiovasc. Med. 2015, 25, 171–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatti, J.S.; Sehrawat, A.; Mishra, J.; Sidhu, I.S.; Navik, U.; Khullar, N.; Kumar, S.; Bhatti, G.K.; Reddy, P.H. Oxidative Stress in the Pathophysiology of Type 2 Diabetes and Related Complications: Current Therapeutics Strategies and Future Perspectives. Free Radic. Biol. Med. 2022, 184, 114–134. [Google Scholar] [CrossRef]
- Bassareo, P.P.; McMahon, C.J. Metabolomics: A New Tool in Our Understanding of Congenital Heart Disease. Children 2022, 9, 1803. [Google Scholar] [CrossRef]
- Griguolo, G.; Bottosso, M.; Vernaci, G.; Miglietta, F.; Dieci, M.V.; Guarneri, V. Gene-Expression Signatures to Inform Neoadjuvant Treatment Decision in HR+/HER2− Breast Cancer: Available Evidence and Clinical Implications. Cancer Treat Rev. 2022, 102, 102323. [Google Scholar] [CrossRef]
- Kitajima, S.; Iwata, Y.; Furuichi, K.; Sagara, A.; Shinozaki, Y.; Toyama, T.; Sakai, N.; Shimizu, M.; Sakurai, T.; Kaneko, S.; et al. Messenger RNA Expression Profile of Sleep-Related Genes in Peripheral Blood Cells in Patients with Chronic Kidney Disease. Clin. Exp. Nephrol. 2016, 20, 218–225. [Google Scholar] [CrossRef] [Green Version]
- Fahed, G.; Aoun, L.; Zerdan, M.B.; Allam, S.; Zerdan, M.B.; Bouferraa, Y.; Assi, H.I. Metabolic Syndrome: Updates on Pathophysiology and Management in 2021. Int. J. Mol. Sci. 2022, 23, 786. [Google Scholar] [CrossRef]
- Hrovat, K.; Likozar, A.R.; Zupan, J.; Šebeštjen, M. Gene Expression Profiling of Markers of Inflammation, Angiogenesis, Coagulation and Fibrinolysis in Patients with Coronary Artery Disease with Very High Lipoprotein(a) Levels Treated with PCSK9 Inhibitors. J. Cardiovasc. Dev. Dis. 2022, 9, 211. [Google Scholar] [CrossRef]
- Li, K.; Kong, R.; Ma, L.; Cao, Y.; Li, W.; Chen, R.; Jiang, L. Identification of Potential M2 Macrophage-Associated Diagnostic Biomarkers in Coronary Artery Disease. Biosci. Rep. 2022, 42, BSR20221394. [Google Scholar] [CrossRef]
- Steinbach, D.; Hermann, J.; Viehmann, S.; Zintl, F.; Gruhn, B. Clinical Implications of PRAME Gene Expression in Childhood Acute Myeloid Leukemia. Cancer Genet. Cytogenet. 2002, 133, 118–123. [Google Scholar] [CrossRef]
- Maneerat, Y.; Prasongsukarn, K.; Benjathummarak, S.; Dechkhajorn, W. PPBP and DEFA1/DEFA3 Genes in Hyperlipidaemia as Feasible Synergistic Inflammatory Biomarkers for Coronary Heart Disease. Lipids Health Dis. 2017, 16, 80. [Google Scholar] [CrossRef] [Green Version]
- Veljkovic, N.; Zaric, B.; Djuric, I.; Obradovic, M.; Sudar-Milovanovic, E.; Radak, D.; Isenovic, E.R. Genetic Markers for Coronary Artery Disease. Medicina 2018, 54, 36. [Google Scholar] [CrossRef] [Green Version]
- Zakynthinos, E.; Pappa, N. Inflammatory Biomarkers in Coronary Artery Disease. J. Cardiol. 2009, 53, 317–333. [Google Scholar] [CrossRef] [Green Version]
- Beermann, J.; Piccoli, M.T.; Viereck, J.; Thum, T. Non-Coding Rnas in Development and Disease: Background, Mechanisms, and Therapeutic Approaches. Physiol. Rev. 2016, 96, 1297–1325. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.Y.; Kuo, H.C. Functional Roles and Networks of Non-Coding RNAs in the Pathogenesis of Neurodegenerative Diseases. J. Biomed. Sci. 2020, 27, 49. [Google Scholar] [CrossRef] [Green Version]
- Dandare, A.; Liaquat, A.; Rafiq, M.; Leonel Javeres, M.N.; Younis, S.; Khan, M.J. Circulating MiR-548c-3p Possesses Good Diagnostic Potential for Metabolic Syndrome. Genes Dis 2022. [Google Scholar] [CrossRef]
- Grillone, K.; Riillo, C.; Riillo, C.; Scionti, F.; Rocca, R.; Rocca, R.; Tradigo, G.; Guzzi, P.H.; Alcaro, S.; Alcaro, S.; et al. Non-Coding RNAs in Cancer: Platforms and Strategies for Investigating the Genomic “Dark Matter”. J. Exp. Clin. Cancer Res. 2020, 39, 117. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Wang, K.; Wu, F.; Wang, W.; Zhang, K.; Hu, H.; Liu, Y.; Jiang, T. CircRNA Disease: A Manually Curated Database of Experimentally Supported CircRNA-Disease Associations. Cell Death Dis. 2018, 9, 4–5. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.; Zhao, G.; Chen, Z.; Wang, X.; Lv, F.; Zhang, Y.; Yang, X.; Liang, W.; Cai, R.; Li, J.; et al. CircRNA-miRNA Association for Coronary Heart Disease. Mol. Med. Rep. 2019, 19, 2527–2536. [Google Scholar] [CrossRef] [Green Version]
- Pan, R.Y.; Liu, P.; Zhou, H.T.; Sun, W.X.; Song, J.; Shu, J.; Cui, G.J.; Yang, Z.J.; Jia, E.Z. Circular RNAs Promote TRPM3 Expression by Inhibiting Hsa-MiR- 130a-3p in Coronary Artery Disease Patients. Oncotarget 2017, 8, 60280–60290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Z.; Li, X.; Gao, C.; Jian, D.; Hao, P.; Rao, L.; Li, M. Peripheral Blood Circular RNA Hsa_circ_0124644 Can Be Used as a Diagnostic Biomarker of Coronary Artery Disease. Sci. Rep. 2017, 7, 39918. [Google Scholar] [CrossRef] [Green Version]
- Hou, C.; Gu, L.; Guo, Y.; Zhou, Y.; Hua, L.; Chen, J.; He, S.; Zhang, S.; Jia, Q.; Zhao, C.; et al. Association between Circular RNA Expression Content and Severity of Coronary Atherosclerosis in Human Coronary Artery. J. Clin. Lab. Anal. 2020, 34, e23552. [Google Scholar] [CrossRef]
- Wu, W.P.; Pan, Y.H.; Cai, M.Y.; Cen, J.M.; Chen, C.; Zheng, L.; Liu, X.; Xiong, X.D. Plasma-Derived Exosomal Circular RNA Hsa_circ_0005540 as a Novel Diagnostic Biomarker for Coronary Artery Disease. Dis. Markers 2020, 2020, 3178642. [Google Scholar] [CrossRef]
- Dudekula, D.B.; Panda, A.C.; Grammatikakis, I. CircInteractome: A Web Tool for Exploring Circular RNAs and Their Interacting Proteins and MicroRNAs. RNA Biol. 2016, 13, 34–42. [Google Scholar] [CrossRef] [Green Version]
- Dandare, A.; Rabia, G.; Khan, M.J. In Silico Analysis of Non-Coding RNAs and Putative Target Genes Implicated in Metabolic Syndrome. Comput. Biol. Med. 2021, 130, 104229. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, X. MiRDB: An Online Database for Prediction of Functional MicroRNA Targets. Nucleic Acids Res. 2020, 48, D127–D131. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics Enrichment Tools: Paths toward the Comprehensive Functional Analysis of Large Gene Lists. Nucleic Acids Res. 2009, 37, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and Integrative Analysis of Large Gene Lists Using DAVID Bioinformatics Resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S.; Kawashima, S.; Nakaya, A. Thed KEGG Databases at GenomeNet. Nucleic Acids Res. 2002, 30, 42–46. [Google Scholar] [CrossRef] [Green Version]
- Smoot, M.E.; Ono, K.; Ruscheinski, J.; Wang, P.L.; Ideker, T. Cytoscape 2.8: New Features for Data Integration and Network Visualization. Bioinformatics 2011, 27, 431–432. [Google Scholar] [CrossRef] [Green Version]
- Rehmsmeier, M.; Steffen, P.; Höchsmann, M.; Giegerich, R. Fast and Effective Prediction of MicroRNA/Target Duplexes. RNA 2004, 10, 1507–1517. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Mallick, B.; Long, D.; Rennie, W.A.; Wolenc, A.; Carmack, C.S.; Ding, Y. CLIP-Based Prediction of Mammalian MicroRNA Binding Sites. Nucleic Acids Res. 2013, 41, e138. [Google Scholar] [CrossRef] [Green Version]
- Alves-Junior, L.; Niemeier, S.; Hauenschild, A.; Rehmsmeier, M.; Merkle, T. Comprehensive Prediction of Novel MicroRNA Targets in Arabidopsis Thaliana. Nucleic Acids Res. 2009, 37, 4010–4021. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Lei, K.; Wu, Z.; Li, W.; Liu, G.; Liu, J.; Cheng, F.; Tang, Y. Network-Based Identification of MicroRNAs as Potential Pharmacogenomic Biomarkers for Anticancer Drugs. Oncotarget 2016, 7, 45584–45596. [Google Scholar] [CrossRef] [Green Version]
- Dandare, A.; Rafiq, M.; Liaquat, A.; Raja, A.A.; Khan, M.J. Identification of Hsa_circ_0092576 Regulatory Network in the Pathogenesis of Coronary Heart Disease. Genes Dis. 2022. [Google Scholar] [CrossRef]
- Wong, C.M.; Tsang, F.H.; Ng, I.O.L. Non-Coding RNAs in Hepatocellular Carcinoma: Molecular Functions and Pathological Implications. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 137–151. [Google Scholar] [CrossRef]
- Jiménez-Lucena, R.; Camargo, A.; Alcalá-Diaz, J.F.; Romero-Baldonado, C.; Luque, R.M.; van Ommen, B.; Delgado-Lista, J.; Ordovás, J.M.; Pérez-Martínez, P.; Rangel-Zúñiga, O.A.; et al. A Plasma Circulating MiRNAs Profile Predicts Type 2 Diabetes Mellitus and Prediabetes: From the CORDIOPREV Study. Exp. Mol. Med. 2018, 50, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Long, B.; Liu, F.; Wang, J.X.; Liu, C.Y.; Zhao, B.; Zhou, L.Y.; Sun, T.; Wang, M.; Yu, T.; et al. A Circular RNA Protects the Heart from Pathological Hypertrophy and Heart Failure by Targeting MiR-223. Eur. Heart J. 2016, 37, 2602a–2611a. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Shen, C.; Wang, Y.; Zou, T.; Zhu, H.; Lu, X.; Li, L.; Yang, B.; Chen, J.; Chen, S.; et al. Identification of Circular RNA Hsa_circ_0001879 and Hsa_circ_0004104 as Novel Biomarkers for Coronary Artery Disease. Atherosclerosis 2019, 286, 88–96. [Google Scholar] [CrossRef]
- Garikipati, V.N.S.; Verma, S.K.; Cheng, Z.; Liang, D.; Truongcao, M.M.; Cimini, M.; Yue, Y.; Huang, G.; Wang, C.; Benedict, C.; et al. Circular RNA CircFndc3b Modulates Cardiac Repair after Myocardial Infarction via FUS/VEGF-A Axis. Nat. Commun. 2019, 10, 4317. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zhao, Z.; Jian, D.; Li, W.; Tang, H.; Li, M. Hsa-CircRNA11783-2 in Peripheral Blood Is Correlated with Coronary Artery Disease and Type 2 Diabetes Mellitus. Diabetes Vasc. Dis. Res. 2017, 14, 510–515. [Google Scholar] [CrossRef] [Green Version]
- Sun, D.; Liu, J.; Zhou, L. Upregulation of Circular RNA Circ-FAM53B Predicts Adverse Prognosis and Accelerates the Progression of Ovarian Cancer via the MiR-646/VAMP2 and MiR-647/MDM2 Signaling Pathways. Oncol. Rep. 2019, 42, 2728–2737. [Google Scholar] [CrossRef]
- Day, E.K.; Sosale, N.G.; Lazzara, M.J.; Opin, C.; Author, B. Cell Signaling Regulation by Protein Phosphorylation: A Multivariate, Heterogeneous, and Context-Dependent Process Graphical Abstract HHS Public Access Author Manuscript. Curr. Opin. Biotechnol. 2016, 40, 185–192. [Google Scholar] [CrossRef] [Green Version]
- Walker, L.A.; Fullerton, D.A.; Buttrick, P.M. Contractile Protein Phosphorylation Predicts Human Heart Disease Phenotypes. Am. J. Physiol. Heart Circ. Physiol. 2013, 304, H1644–H1650. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Cheng, Z.; Yang, J. MiR-23 Regulates Cell Proliferation and Apoptosis of Vascular Smooth Muscle Cells in Coronary Heart Disease. Pathol. Res. Pract. 2018, 214, 1873–1878. [Google Scholar] [CrossRef]
- Huang, S.; Zeng, Z.; Sun, Y.; Cai, Y.; Xu, X.; Li, H.; Wu, S. Association Study of Hsa_circ_0001946, Hsa-MiR-7-5p and PARP1 in Coronary Atherosclerotic Heart Disease. Int. J. Cardiol. 2021, 328, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Gu, G.L.; Xu, X.L.; Sun, X.T.; Zhang, J.; Guo, C.F.; Wang, C.S.; Sun, B.; Guo, G.L.; Ma, K.; Huang, Y.Y.; et al. Cardioprotective Effect of MicroRNA-21 in Murine Myocardial Infarction. Cardiovasc. Ther. 2015, 33, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Martino, T.A.; Young, M.E. Influence of the Cardiomyocyte Circadian Clock on Cardiac Physiology and Pathophysiology. J. Biol. Rhythm. 2015, 30, 183–205. [Google Scholar] [CrossRef] [PubMed]
- Glen Pyle, W.; Martino, T.A. Circadian Rhythms Influence Cardiovascular Disease Differently in Males and Females: Role of Sex and Gender. Curr. Opin. Physiol. 2018, 5, 30–37. [Google Scholar] [CrossRef]
- Yeung, S.L.A.; Lam, H.S.H.; Schooling, C.M. Vascular Endothelial Growth Factor and Ischemic Heart Disease Risk: A Mendelian Randomization Study. J. Am. Heart Assoc. 2017, 6, e005619. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.Z.; Wen, L.; Wei, X.; Wang, Q.R.; Xu, L.W.; Zhang, H.M.; Liu, W.C. Inhibition of MiR-7 Promotes Angiogenesis in Human Umbilical Vein Endothelial Cells by Upregulating VEGF via KLF4. Oncol. Rep. 2016, 36, 1569–1575. [Google Scholar] [CrossRef] [Green Version]
- Han, N.; Xu, H.; Yu, N.; Wu, Y.; Yu, L. MiR-203a-3p Inhibits Retinal Angiogenesis and Alleviates Proliferative Diabetic Retinopathy in Oxygen-Induced Retinopathy (OIR) Rat Model via Targeting VEGFA and HIF-1α. Clin. Exp. Pharmacol. Physiol. 2020, 47, 85–94. [Google Scholar] [CrossRef]
- Chen, X.; Sun, R.; Yang, D.; Jiang, C.; Liu, Q. LINC00167 Regulates RPE Differentiation by Targeting the MiR-203a-3p/SOCS3 Axis. Mol. Ther.-Nucleic Acids 2020, 19, 1015–1026. [Google Scholar] [CrossRef]
- Rothman, A.; Restrepo, H.; Sarukhanov, V.; Evans, W.N.; Wiencek, R.G.; Williams, R.; Hamburger, N.; Anderson, K.; Balsara, J.; Mann, D. Assessment of MicroRNA and Gene Dysregulation in Pulmonary Hypertension by Endoarterial Biopsy. Pulm. Circ. 2017, 7, 455–464. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Wei, L. Regulation of JAK/STAT Signalling by SOCS in the Myocardium. Cardiovasc. Res. 2012, 96, 345–347. [Google Scholar] [CrossRef] [Green Version]
- Terrell, A.M.; Crisostomo, P.R.; Wairiuko, G.M.; Wang, M.; Morrell, E.D.; Meldrum, D.R. JAK/STAT/SOCS Signaling Circuits and Associated Cytokine-Mediated Inflammation and Hypertrophy in the Heart. Shock 2006, 26, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Witte, S.; Muljo, S.A. Integrating Non-Coding RNAs in JAK-STAT Regulatory Networks. JAK-STAT 2014, 3, e28055. [Google Scholar] [CrossRef]
- Vasuri, F.; Visani, M.; Acquaviva, G.; Brand, T.; Fiorentino, M.; Pession, A.; Tallini, G.; D’Errico, A.; De Biase, D. Role of MicroRNAs in the Main Molecular Pathways of Hepatocellular Carcinoma. World J. Gastroenterol. 2018, 24, 2647–2660. [Google Scholar] [CrossRef] [PubMed]
- Li, H.W.; Zeng, H.S. Regulation of JAK/STAT Signal Pathway by MiR-21 in the Pathogenesis of Juvenile Idiopathic Arthritis. World J. Pediatr. 2020, 16, 502–513. [Google Scholar] [CrossRef] [Green Version]
- Li, H.W.; Xie, Y.; Li, F.; Sun, G.C.; Chen, Z.; Zeng, H.S. Effect of MiR-19a and MiR-21 on the JAK/STAT Signaling Pathway in the Peripheral Blood Mononuclear Cells of Patients with Systemic Juvenile Idiopathic Arthritis. Exp. Ther. Med. 2016, 11, 2531–2536. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Cui, M.; Zhang, A.; Tong, L.; Wang, K.; Li, K.; Wang, X.; Sun, Z.; Zhang, H. MiR-548c Impairs Migration and Invasion of Endometrial and Ovarian Cancer Cells via Downregulation of Twist. J. Exp. Clin. Cancer Res. 2016, 35, 10. [Google Scholar] [CrossRef] [Green Version]
- Kasiviswanathan, D.; Chinnasamy Perumal, R.; Bhuvaneswari, S.; Kumar, P.; Sundaresan, L.; Philip, M.; Puthenpurackal Krishnankutty, S.; Chatterjee, S. Interactome of MiRNAs and Transcriptome of Human Umbilical Cord Endothelial Cells Exposed to Short-Term Simulated Microgravity. npj Microgravity 2020, 6, 18. [Google Scholar] [CrossRef]
- Ortiz-Muñoz, G.; Martin-Ventura, J.L.; Hernandez-Vargas, P.; Mallavia, B.; Lopez-Parra, V.; Lopez-Franco, O.; Muñoz-Garcia, B.; Fernandez-Vizarra, P.; Ortega, L.; Egido, J.; et al. Suppressors of Cytokine Signaling Modulate JAK/STAT-Mediated Cell Responses during Atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 525–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grote, K.; Luchtefeld, M.; Schieffer, B. JANUS under Stress-Role of JAK/STAT Signaling Pathway in Vascular Diseases. Vascul. Pharmacol. 2005, 43, 357–363. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, Y.; Qiao, S.; Yin, L. Effects of Apelin on Cardiovascular Aging. Front. Physiol. 2017, 8, 1035. [Google Scholar] [CrossRef] [Green Version]
- Földes, G.; Horkay, F.; Szokodi, I.; Vuolteenaho, O.; Ilves, M.; Lindstedt, K.A.; Mäyränpää, M.; Sármán, B.; Seres, L.; Skoumal, R.; et al. Circulating and Cardiac Levels of Apelin, the Novel Ligand of the Orphan Receptor APJ, in Patients with Heart Failure. Biochem. Biophys. Res. Commun. 2003, 308, 480–485. [Google Scholar] [CrossRef]
- Zhong, J.C.; Zhang, Z.Z.; Wang, W.; McKinnie, S.M.K.; Vederas, J.C.; Oudit, G.Y. Targeting the Apelin Pathway as a Novel Therapeutic Approach for Cardiovascular Diseases. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 1942–1950. [Google Scholar] [CrossRef]
- Chen, T.; Wu, B.; Lin, R. Association of Apelin and Apelin Receptor with the Risk of Coronary Artery Disease: A Meta-Analysis of Observational Studies. Oncotarget 2017, 8, 57345–57355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Zhao, M.; Du, Y.; Liu, Y.; Zhao, G.; Ye, L.; Li, Q.; Li, H.; Wang, X.; Liu, X.; et al. MicroRNA-195 Suppresses the Progression of Lung Adenocarcinoma by Directly Targeting Apelin. Thorac. Cancer 2019, 10, 1419–1430. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Deng, L.; Zhao, D.; Chen, L.; Yao, Z.; Guo, X.; Liu, X.; Lv, L.; Leng, B.; Xu, W.; et al. MicroRNA-503 Promotes Angiotensin II-Induced Cardiac Fibrosis by Targeting Apelin-13. J. Cell Mol. Med. 2016, 20, 495–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitkin, S.L.; Maguire, J.J.; Kuc, R.E.; Davenport, A.P. Modulation of the Apelin/APJ System in Heart Failureand Atherosclerosis in Man. Br. J. Pharmacol. 2010, 160, 1785–1795. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, T.; Kihara, M.; Imai, N.; Yoshida, S.I.; Shimoyamada, H.; Yasuzaki, H.; Ishida, J.; Toya, Y.; Kiuchi, Y.; Hirawa, N.; et al. Requirement of Apelin-Apelin Receptor System for Oxidative Stress-Linked Atherosclerosis. Am. J. Pathol. 2007, 171, 1705–1712. [Google Scholar] [CrossRef] [Green Version]
- Chun, H.J.; Ali, Z.A.; Kojima, Y.; Kundu, R.K.; Sheikh, A.Y.; Agrawal, R.; Zheng, L.; Leeper, N.J.; Pearl, N.E.; Patterson, A.J.; et al. Apelin Signaling Antagonizes Ang II Effects in Mouse Models of Atherosclerosis. J. Clin. Investig. 2008, 118, 3343–3354. [Google Scholar] [CrossRef] [Green Version]
- Riolo, G.; Cantara, S.; Marzocchi, C.; Ricci, C. MiRNA Targets: From Prediction Tools to Experimental Validation. Methods Protoc. 2021, 4, 1. [Google Scholar] [CrossRef]
- Peterson, S.M.; Thompson, J.A.; Ufkin, M.L.; Sathyanarayana, P.; Liaw, L.; Congdon, C.B. Common Features of MicroRNA Target Prediction Tools. Front. Genet. 2014, 5, 23. [Google Scholar] [CrossRef] [Green Version]
- Ab Mutalib, N.-S.; Sulaiman, S.A.; Jamal, R. Computational Tools for MicroRNA Target Prediction; Elsevier Inc.: Amsterdam, The Netherlands, 2009; ISBN 9780128145135. [Google Scholar]
- Shah, M.Y.; Pan, X.; Fix, L.N.; Farwell, M.A.; Zhang, B. 5-Fluorouracil Drug Alters the MicroRNA Expression Profiles in MCF-7 Breast Cancer Cells. J. Cell Physiol. 2011, 226, 1868–1878. [Google Scholar] [CrossRef]
- Collins, C.; Weiden, P.L. Cardiotoxicity of 5-Fluorouracil. Cancer Treat Rep. 1987, 71, 733–736. [Google Scholar] [PubMed]
- Yuan, C.; Parekh, H.; Allegra, C.; George, T.J.; Starr, J.S. 5-FU Induced Cardiotoxicity: Case Series and Review of the Literature. Cardio-Oncology 2019, 5, 13. [Google Scholar] [CrossRef] [Green Version]
- Hardy, R.S.; Raza, K.; Cooper, M.S. Therapeutic Glucocorticoids: Mechanisms of Actions in Rheumatic Diseases. Nat. Rev. Rheumatol. 2020, 16, 133–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Libby, P. History of Discovery Inflammation in Atherosclerosis Inflammation: An Enduring Flame. Arter. Thromb. Vasc. Biol. 2012, 32, 2045–2051. [Google Scholar] [CrossRef] [Green Version]
- Recio, C.; Oguiza, A.; Lazaro, I.; Mallavia, B.; Egido, J.; Gomez-Guerrero, C. Suppressor of Cytokine Signaling 1-Derived Peptide Inhibits Janus Kinase/Signal Transducers and Activators of Transcription Pathway and Improves Inflammation and Atherosclerosis in Diabetic Mice. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 1953–1960. [Google Scholar] [CrossRef] [Green Version]
- Husain, K.; Hernandez, W.; Ansari, R.A.; Ferder, L. Inflammation, Oxidative Stress and Renin Angiotensin System in Atherosclerosis. World J. Biol. Chem. 2015, 6, 209. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession ID | Gene Expression Platform | Sample Type | Number of Controls | Number of Cases | Total Samples | Location |
|---|---|---|---|---|---|---|
| GSE71226 | GPL570 Affymetrix Human Genome U133 Plus 2.0 Array | Peripheral blood | 03 | 03 | 06 | China |
| GSE12288 | GPL96 Affymetrix Human Genome U133A Array | Peripheral blood | 112 | 110 | 222 | Switzerland |
| GSE56885 | GPL15207 Affymetrix Human Gene Expression Array | Peripheral blood mononuclear cells | 02 | 04 | 06 | India |
| GSE42148 | GPL13607 Agilent-028004 SurePrint G3 Human GE 8x60K Microarray | Peripheral blood | 11 | 13 | 23 | India |
| GSE20681 | GPL4133 Agilent-014850 Whole Human Genome Microarray 4x44K G4112F | Peripheral blood | 90 | 90 | 198 | USA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rafiq, M.; Dandare, A.; Javed, A.; Liaquat, A.; Raja, A.A.; Awan, H.M.; Khan, M.J.; Naeem, A. Competing Endogenous RNA Regulatory Networks of hsa_circ_0126672 in Pathophysiology of Coronary Heart Disease. Genes 2023, 14, 550. https://doi.org/10.3390/genes14030550
Rafiq M, Dandare A, Javed A, Liaquat A, Raja AA, Awan HM, Khan MJ, Naeem A. Competing Endogenous RNA Regulatory Networks of hsa_circ_0126672 in Pathophysiology of Coronary Heart Disease. Genes. 2023; 14(3):550. https://doi.org/10.3390/genes14030550
Chicago/Turabian StyleRafiq, Muhammad, Abdullahi Dandare, Arham Javed, Afrose Liaquat, Afraz Ahmad Raja, Hassaan Mehboob Awan, Muhammad Jawad Khan, and Aisha Naeem. 2023. "Competing Endogenous RNA Regulatory Networks of hsa_circ_0126672 in Pathophysiology of Coronary Heart Disease" Genes 14, no. 3: 550. https://doi.org/10.3390/genes14030550
APA StyleRafiq, M., Dandare, A., Javed, A., Liaquat, A., Raja, A. A., Awan, H. M., Khan, M. J., & Naeem, A. (2023). Competing Endogenous RNA Regulatory Networks of hsa_circ_0126672 in Pathophysiology of Coronary Heart Disease. Genes, 14(3), 550. https://doi.org/10.3390/genes14030550