
The Synchronized Progression from Mitosis to Meiosis in Female Primordial Germ Cells between Layers and Broilers
Abstract
:1. Introduction
2. Materials and Methods
2.1. PGCs: Isolation and Culture
2.2. Alkaline Phosphatase Staining
2.3. Immunofluorescence Staining
2.4. Cell Counting Kit-8 Assay
2.5. EdU Assay
2.6. RNA Extraction
2.7. RNA-Sequencing
2.8. Quantitative Real-Time PCR
2.9. Statistical Analyses
3. Results
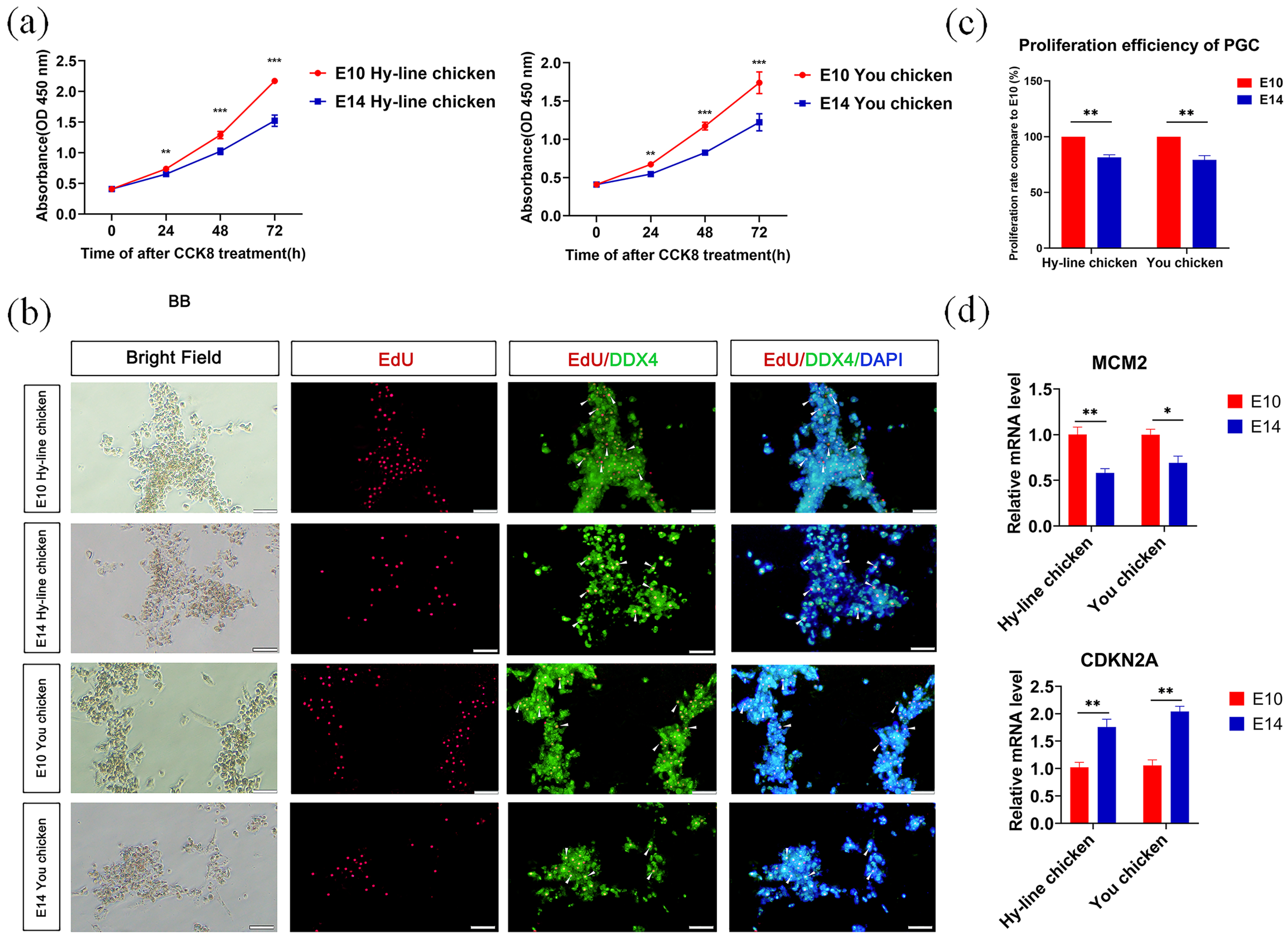
3.1. Cultivation and Characterization of PGCs
3.2. Comparison of the Proliferation Ability of PGCs from the Two Developmental Stages
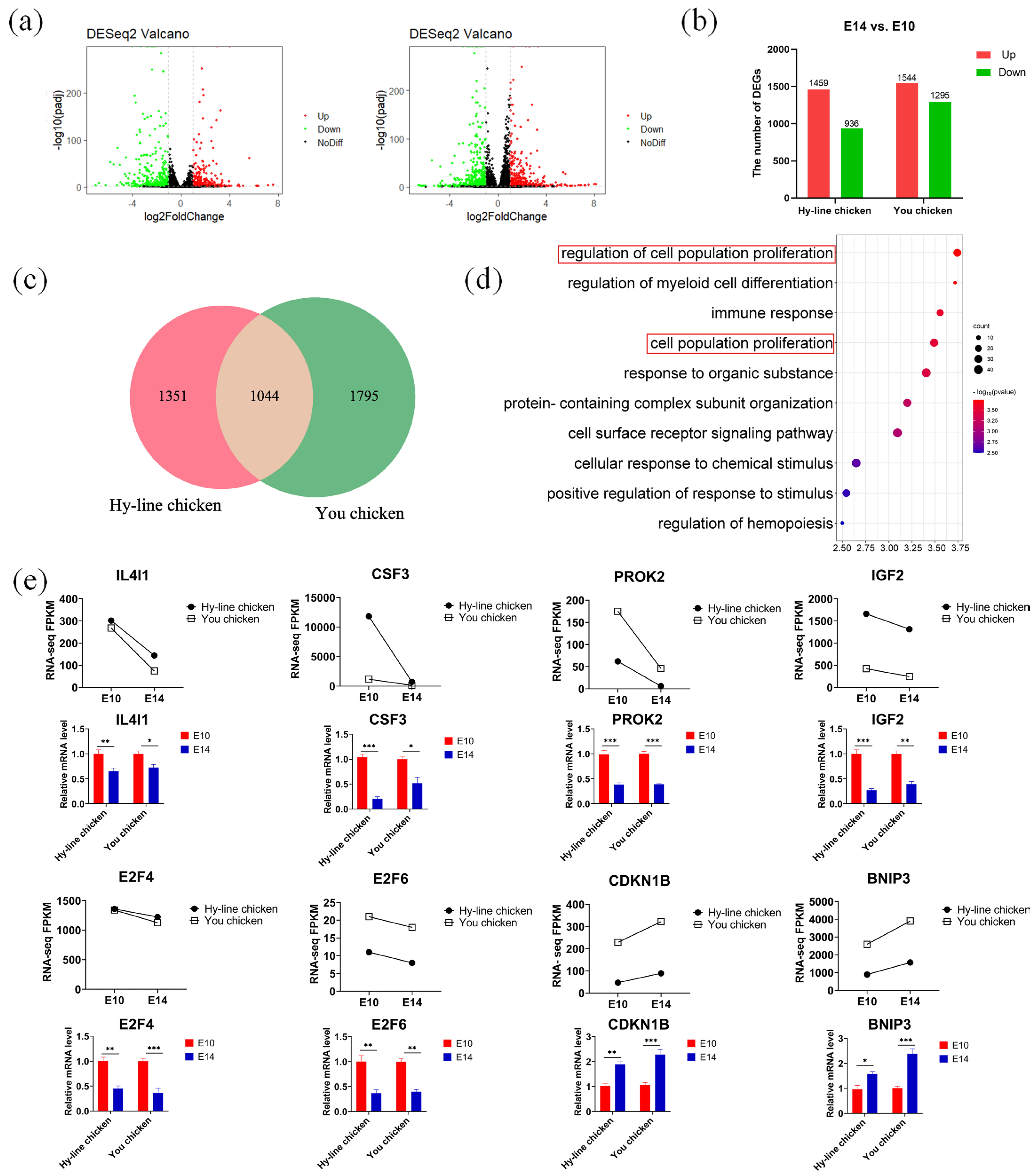
3.3. RNA Sequencing of E10 and E14 PGCs
3.4. Discovering Differential Co-Expression Genes Involved in Mitosis between You Chicken and Hy-Line Chicken
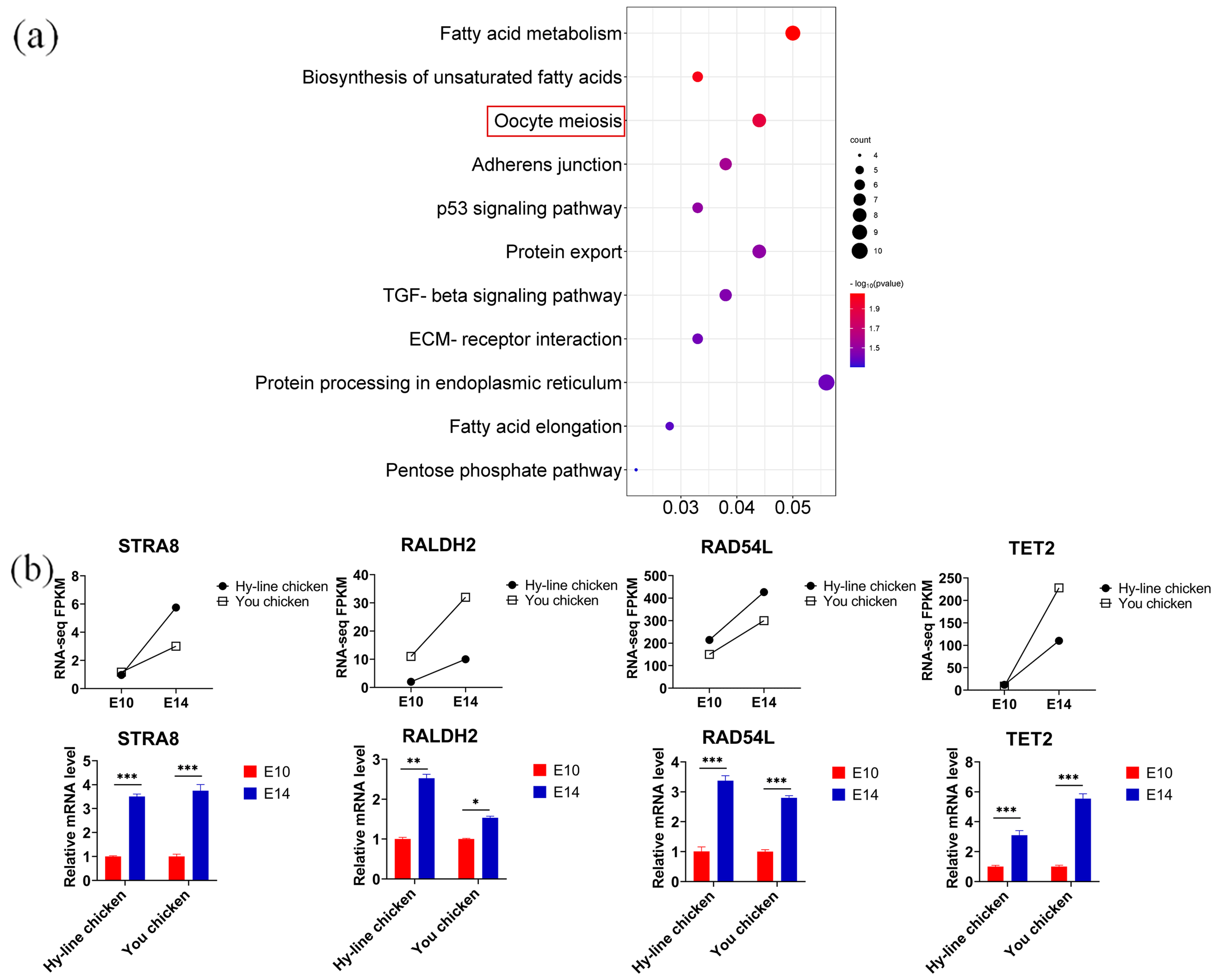
3.5. Discovering Differential Co-Expression Genes Involved in Meiosis between You Chicken and Hy-Line Chicken
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Khan, M.J.; Ullah, A.; Basit, S. Genetic Basis of Polycystic Ovary Syndrome (PCOS): Current Perspectives. Appl. Clin. Genet. 2019, 12, 249–260. [Google Scholar] [CrossRef] [Green Version]
- Ono, T.; Fang, Y.; Spector, D.L.; Hirano, T. Spatial and Temporal Regulation of Condensins I and II in Mitotic Chromosome Assembly in Human Cells. Mol. Biol. Cell 2004, 15, 3296–3308. [Google Scholar] [CrossRef] [PubMed]
- Cantú, A.V.; Laird, D.J. A pilgrim’s progress: Seeking meaning in primordial germ cell migration. Stem Cell Res. 2017, 24, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Hamburger, V. A series of normal stages in the development of the chick embryo. J. Morph 1951, 88, 49–92. [Google Scholar] [CrossRef]
- Pellegrini, S.; Chimienti, R.; Scotti, G.M.; Giannese, F.; Lazarevic, D.; Manenti, F.; Poggi, G.; Lombardo, M.T.; Cospito, A.; Nano, R.; et al. Transcriptional dynamics of induced pluripotent stem cell differentiation into β cells reveals full endodermal commitment and homology with human islets. Cytotherapy 2021, 23, 311–319. [Google Scholar] [CrossRef]
- De Melo Bernardo, A.; Sprenkels, K.; Rodrigues, G.; Noce, T.; Chuva De Sousa Lopes, S.M. Chicken primordial germ cells use the anterior vitelline veins to enter the embryonic circulation. Biol. Open 2012, 1, 1146–1152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Meng, L.; Wang, S.; Man, Q.; Jiang, Y.; Zhu, G. Transcriptional dynamics of the circulating chicken primordial germ cells revealing key genes in cell adhesion and proliferation prior to gonad colonization. Mol. Reprod. Dev. 2022, 89, 214–226. [Google Scholar] [CrossRef] [PubMed]
- Estermann, M.A.; Williams, S.; Hirst, C.E.; Roly, Z.Y.; Serralbo, O.; Adhikari, D.; Powell, D.; Major, A.T.; Smith, C.A. Insights into Gonadal Sex Differentiation Provided by Single-Cell Transcriptomics in the Chicken Embryo. Cell Rep. 2020, 31, 107491. [Google Scholar] [CrossRef]
- Wartenberg, H.; Lenz, E.; Schweikert, H.-U. Sexual differentiation and the germ cell in sex reversed gonads after aromatase inhibition in the chicken embryo. Andrologia 1992, 24, 1–6. [Google Scholar] [CrossRef]
- Jiang, Y.; Peng, Z.; Man, Q.; Wang, S.; Huang, X.; Meng, L.; Wang, H.; Zhu, G. H3K27ac chromatin acetylation and gene expression analysis reveal sex- and situs-related differences in developing chicken gonads. Biol. Sex Differ. 2022, 13, 6. [Google Scholar] [CrossRef]
- Hughes, G.C. The Population of Germ Cells in the Developing Female Chick. Development 1963, 11, 513–536. [Google Scholar] [CrossRef]
- Rengaraj, D.; Cha, D.G.; Lee, H.J.; Lee, K.Y.; Choi, Y.H.; Jung, K.M.; Kim, Y.M.; Choi, H.J.; Choi, H.J.; Yoo, E.; et al. Dissecting chicken germ cell dynamics by combining a germ cell tracing transgenic chicken model with single-cell RNA sequencing. Comput. Struct. Biotechnol. J. 2022, 20, 1654–1669. [Google Scholar] [CrossRef]
- Lei, L.; Spradling, A.C. Mouse primordial germ cells produce cysts that partially fragment prior to meiosis. Development 2013, 140, 2075–2081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kagiwada, S.; Kurimoto, K.; Hirota, T.; Yamaji, M.; Saitou, M. Replication-coupled passive DNA demethylation for the erasure of genome imprints in mice. EMBO J. 2013, 32, 340–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diril, M.K.; Ratnacaram, C.K.; Padmakumar, V.C.; Du, T.; Wasser, M.; Coppola, V.; Tessarollo, L.; Kaldis, P. Cyclin-dependent kinase 1 (Cdk1) is essential for cell division and suppression of DNA re-replication but not for liver regeneration. Proc. Natl. Acad. Sci. USA 2012, 109, 3826–3831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Risal, S.; Zhang, J.; Adhikari, D.; Liu, X.; Shao, J.; Hu, M.; Busayavalasa, K.; Tu, Z.; Chen, Z.; Kaldis, P.; et al. MASTL is essential for anaphase entry of proliferating primordial germ cells and establishment of female germ cells in mice. Cell Discov. 2017, 3, 16052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Messiaen, S.; Le Bras, A.; Duquenne, C.; Barroca, V.; Moison, D.; Déchamps, N.; Doussau, M.; Bauchet, A.L.; Guerquin, M.J.; Livera, G.; et al. Rad54 is required for the normal development of male and female germ cells and contributes to the maintainance of their genome integrity after genotoxic stress. Cell Death Dis. 2013, 4, e774. [Google Scholar] [CrossRef] [Green Version]
- Li, X.H.; Zhang, J.R.; Shi, G.; Shi, C.; Ji, Z.T. Study on the Quality and Related Volatile Compounds of Beijing You- Chicken at Different Temperatures. Indian J. Anim. Res. 2022, 56, 1176–1181. [Google Scholar] [CrossRef]
- Shi, L.; Sun, Y.Y.; Xu, H.; Liu, Y.F.; Li, Y.L.; Huang, Z.Y.; Ni, A.X.; Chen, C.; Wang, P.L.; Ye, J.H.; et al. Effect of age at photostimulation on reproductive performance of Beijing-You Chicken breeders. Poult. Sci. 2019, 98, 4522–4529. [Google Scholar] [CrossRef]
- Van de Lavoir, M.-C.; Diamond, J.H.; Leighton, P.A.; Mather-Love, C.; Heyer, B.S.; Bradshaw, R.; Kerchner, A.; Hooi, L.T.; Gessaro, T.M.; Swanberg, S.E.; et al. Germline transmission of genetically modified primordial germ cells. Nature 2006, 441, 766–769. [Google Scholar] [CrossRef]
- Meng, L.; Wang, S.; Jiang, H.; Hua, Y.; Yin, B.; Huang, X.; Man, Q.; Wang, H.; Zhu, G. Oct4 dependent chromatin activation is required for chicken primordial germ cell migration. Stem Cell Rev. Rep. 2022, 18, 2535–2546. [Google Scholar] [CrossRef] [PubMed]
- Yin, B.; Zhang, K.; Du, X.; Cai, H.; Ye, T.; Wang, H. Developmental switch from morphological replication to compensatory growth for salamander lung regeneration. Cell Prolif. 2023, 56, e13369. [Google Scholar] [CrossRef] [PubMed]
- Lei, M.; Tye, B.K. Initiating DNA synthesis: From recruiting to activating the MCM complex. J. Cell Sci. 2001, 114, 1447–1454. [Google Scholar] [CrossRef] [PubMed]
- Duronio, R.J.; Xiong, Y. Signaling Pathways That Control Cell Proliferation. Cold Spring Harb. Perspect. Biol. 2013, 5, 12. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.A.; Roeszler, K.N.; Bowles, J.; Koopman, P.; Sinclair, A.H. Onset of meiosis in the chicken embryo; evidence of a role for retinoic acid. BMC Dev. Biol. 2008, 8, 85. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.; Yu, P.; Leghari, I.H.; Ge, C.; Mi, Y.; Zhang, C. RALDH2, the enzyme for retinoic acid synthesis, mediates meiosis initiation in germ cells of the female embryonic chickens. Amino Acids 2013, 44, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Yamaya, K.; Wang, B.; Memar, N.; Odiba, A.S.; Woglar, A.; Gartner, A.; Villeneuve, A.M. Disparate roles for C. elegans DNA translocase paralogs RAD-54.L and RAD-54.B in meiotic prophase germ cells. bioRxiv 2022, 2022.2012.2012.520157. [Google Scholar] [CrossRef]
- Wang, H.; Liu, L.; Gou, M.; Huang, G.; Tian, C.; Yang, J.; Wang, H.; Xu, Q.; Xu, G.L.; Liu, L. Roles of Tet2 in meiosis, fertility and reproductive aging. Protein Cell 2020, 12, 578–585. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, S.; Zhou, Q.; Liao, Y.; Luo, W.; Peng, Z.; Ren, R.; Wang, H. Disturbance of calcium homeostasis and myogenesis caused by TET2 deletion in muscle stem cells. Cell Death Discov. 2022, 8, 236. [Google Scholar] [CrossRef]
- Yamaguchi, S.; Hong, K.; Liu, R.; Shen, L.; Inoue, A.; Diep, D.; Zhang, K.; Zhang, Y. Tet1 controls meiosis by regulating meiotic gene expression. Nature 2012, 492, 443–447. [Google Scholar] [CrossRef] [Green Version]
- Sélénou, C.; Brioude, F.; Giabicani, E.; Sobrier, M.-L.; Netchine, I. IGF2: Development, Genetic and Epigenetic Abnormalities. Cells 2022, 11, 1886. [Google Scholar] [CrossRef] [PubMed]
- Moreira, D.P.; Melo, R.M.C.; Weber, A.A.; Rizzo, E. Insulin-like growth factors 1 and 2 are associated with testicular germ cell proliferation and apoptosis during fish reproduction. Reprod. Fertil. Dev. 2020, 32, 988–998. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.D.; Huang, Y.Y.; Liu, W.Q.; Zhang, X.S. Up-Regulation of microRNA-210 Is Associated with Spermatogenesis by Targeting IGF2 in Male Infertility. Med. Sci. Monit. 2016, 22, 6. [Google Scholar] [CrossRef] [Green Version]
- Tang, S.; Sun, D.; Ou, J.; Zhang, Y.; Xu, G.; Zhang, Y. Evaluation of the IGFs (IGF1 and IGF2) Genes as Candidates for Growth, Body Measurement, Carcass, and Reproduction Traits in Beijing You and Silkie Chickens. Anim. Biotechnol. 2010, 21, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Helin, K. Regulation of cell proliferation by the E2F transcription factors. Curr. Opin. Genet. Dev. 1998, 8, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Zou, Z.; Lin, S.; Chen, X.; Wang, F.; Li, X.; Dai, J. Identification and functional analysis of spermatogenesis-associated gene modules in azoospermia by weighted gene coexpression network analysis. J. Cell. Biochem. 2019, 120, 3934–3944. [Google Scholar] [CrossRef]
- Kehoe, S.M.; Oka, M.; Hankowski, K.E.; Reichert, N.; Garcia, S.; McCarrey, J.R.; Gaubatz, S.; Terada, N. A Conserved E2F6-Binding Element in Murine Meiosis-Specific Gene Promoters1. Biol. Reprod. 2008, 79, 921–930. [Google Scholar] [CrossRef] [Green Version]
- Cui, C.; Han, S.; Yin, H.; Luo, B.; Shen, X.; Yang, F.; Liu, Z.; Zhu, Q.; Li, D.; Wang, Y. FOXO3 Is Expressed in Ovarian Tissues and Acts as an Apoptosis Initiator in Granulosa Cells of Chickens. BioMed Res. Int. 2019, 2019, 6902906. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, T.; Abe, Y.; Wakai, T.; Hoshino, Y.; Miyamoto, A.; Sato, E. Recent Patents of TGF-Beta; Family and VEGF Associated with Ovarian Follicular Development in Mammals. Recent Pat. DNA Gene Seq. 2007, 1, 195–199. [Google Scholar] [CrossRef]
- Anderson, E.L.; Baltus, A.E.; Roepers-Gajadien, H.L.; Hassold, T.J.; de Rooij, D.G.; van Pelt, A.M.M.; Page, D.C. Stra8 and its inducer, retinoic acid, regulate meiotic initiation in both spermatogenesis and oogenesis in mice. Proc. Natl. Acad. Sci. USA 2008, 105, 14976–14980. [Google Scholar] [CrossRef] [Green Version]
- Le Bouffant, R.; Guerquin, M.J.; Duquenne, C.; Frydman, N.; Coffigny, H.; Rouiller-Fabre, V.; Frydman, R.; Habert, R.; Livera, G. Meiosis initiation in the human ovary requires intrinsic retinoic acid synthesis. Hum. Reprod. 2010, 25, 2579–2590. [Google Scholar] [CrossRef] [Green Version]
- Meng, L.; Zhang, Y.; Hua, Y.; Ma, Y.; Wang, H.; Li, X.; Jiang, Y.; Zhu, G. Identification of oogonial stem cells in chicken ovary. Cell Prolif. 2023, e13371. [Google Scholar] [CrossRef] [PubMed]
- Swindell, E.C.; Thaller, C.; Sockanathan, S.; Petkovich, M.; Jessell, T.M.; Eichele, G. Complementary Domains of Retinoic Acid Production and Degradation in the Early Chick Embryo. Dev. Biol. 1999, 216, 282–296. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.; Xu, Y.; Yu, D.; Yu, D.; Du, W. Comparative analysis of temporal gene expression patterns in the developing ovary of the embryonic chicken. J. Reprod. Dev. 2015, 61, 123–133. [Google Scholar] [CrossRef] [Green Version]
- Larose, H.; Shami, A.N.; Abbott, H.; Manske, G.; Lei, L.; Hammoud, S.S. Chapter Eight-Gametogenesis: A journey from inception to conception. Curr. Top. Dev. Biol. 2019, 132, 257–310. [Google Scholar] [PubMed]
- Gu, T.-P.; Guo, F.; Yang, H.; Wu, H.-P.; Xu, G.-F.; Liu, W.; Xie, Z.-G.; Shi, L.; He, X.; Jin, S.-g.; et al. The role of Tet3 DNA dioxygenase in epigenetic reprogramming by oocytes. Nature 2011, 477, 606–610. [Google Scholar] [CrossRef] [PubMed]
- Hackett, J.A.; Sengupta, R.; Zylicz, J.J.; Murakami, K.; Lee, C.; Down, T.A.; Surani, M.A. Germline DNA Demethylation Dynamics and Imprint Erasure Through 5-Hydroxymethylcytosine. Science 2013, 339, 448–452. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, H.; Sakurai, T.; Miura, F.; Imai, M.; Mochiduki, K.; Yanagisawa, E.; Sakashita, A.; Wakai, T.; Suzuki, Y.; Ito, T. High-resolution DNA methylome analysis of primordial germ cells identifies gender-specific reprogramming in mice. Genome Res. 2013, 23, 616–627. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.-Y.; Liang, X.-W.; Schatten, H.; Sun, Q.-Y. Active DNA demethylation in mammalian preimplantation embryos: New insights and new perspectives. Mol. Hum. Reprod. 2012, 18, 333–340. [Google Scholar] [CrossRef] [Green Version]
- Ge, Z.-J.; Schatten, H.; Zhang, C.-L.; Sun, Q.-Y. Oocyte ageing and epigenetics. Reprod. Off. J. Soc. Study Fertil. 2015, 149, R103–R114. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Gene ID | log2 Fold Change (You Chicken) | log2 Fold Change (Hy-Line Chicken) | Gene Name |
|---|---|---|---|
| ENSGALG00010009474 | 4.637687 | 0.33777 | ACVR2A |
| ENSGALG00010018099 | 0.640534 | 0.92462 | SMAD3 |
| ENSGALG00010015624 | 1.571854 | 0.709248 | ITGA1 |
| ENSGALG00010024658 | −4.02396 | −3.205 | CSF3 |
| ENSGALG00010023281 | −0.39614 | −0.84782 | B2M |
| ENSGALG00010005131 | −1.76463 | −0.72073 | IL8L2 |
| ENSGALG00010024580 | −0.33743 | −0.40021 | IGF2 |
| ENSGALG00010028080 | −0.84127 | −0.85296 | MUSTN1 |
| ENSGALG00010005913 | −1.23741 | −0.51199 | NFKBIA |
| ENSGALG00010020507 | −0.7204 | −2.24103 | ANKRD1 |
| Gene ID | log2 Fold Change (You Chicken) | log2 Fold Change (Hy-Line Chicken) | Gene Name |
|---|---|---|---|
| ENSGALG00010018388 | 0.331045 | 0.569627 | SMC3 |
| ENSGALG00010024756 | 1.115203 | 1.449926 | CDK2 |
| ENSGALG00000041923 | 0.730368 | 1.49584 | ESPL1 |
| ENSGALG00010017105 | 1.318177 | 3.994044 | PLK1 |
| ENSGALG00010022702 | 0.952322 | 3.161609 | CDC20 |
| ENSGALG00010017024 | 1.104456 | 3.314962 | BUB1 |
| ENSGALG00010018531 | 0.779517 | 0.531854 | ADCY6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Y.; Wu, W.; Zhang, Y.; Wang, X.; Wei, J.; Guo, X.; Xue, M.; Zhu, G. The Synchronized Progression from Mitosis to Meiosis in Female Primordial Germ Cells between Layers and Broilers. Genes 2023, 14, 781. https://doi.org/10.3390/genes14040781
Ma Y, Wu W, Zhang Y, Wang X, Wei J, Guo X, Xue M, Zhu G. The Synchronized Progression from Mitosis to Meiosis in Female Primordial Germ Cells between Layers and Broilers. Genes. 2023; 14(4):781. https://doi.org/10.3390/genes14040781
Chicago/Turabian StyleMa, Yuxiao, Wenhui Wu, Yun Zhang, Xuzhao Wang, Jiahui Wei, Xiaotong Guo, Man Xue, and Guiyu Zhu. 2023. "The Synchronized Progression from Mitosis to Meiosis in Female Primordial Germ Cells between Layers and Broilers" Genes 14, no. 4: 781. https://doi.org/10.3390/genes14040781
APA StyleMa, Y., Wu, W., Zhang, Y., Wang, X., Wei, J., Guo, X., Xue, M., & Zhu, G. (2023). The Synchronized Progression from Mitosis to Meiosis in Female Primordial Germ Cells between Layers and Broilers. Genes, 14(4), 781. https://doi.org/10.3390/genes14040781