
Whole-Genome Sequencing Data Reveal New Loci Affecting Milk Production in German Black Pied Cattle (DSN)
 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Population
2.2. Genotypes
2.3. Phenotypes
2.4. Genome-Wide Association Studies (GWASs)
2.5. Significance Threshold and QTL Definition
2.6. QTL Annotation
3. Results
3.1. Genomic Regions Associated with Milk Production Traits
3.2. Most Significant Locus Affecting Milk Fat and Protein Content on Chromosome 5
3.3. Loci on Chromosome 1 and 8 Affect Milk Yield as Well as Fat and Protein Yields
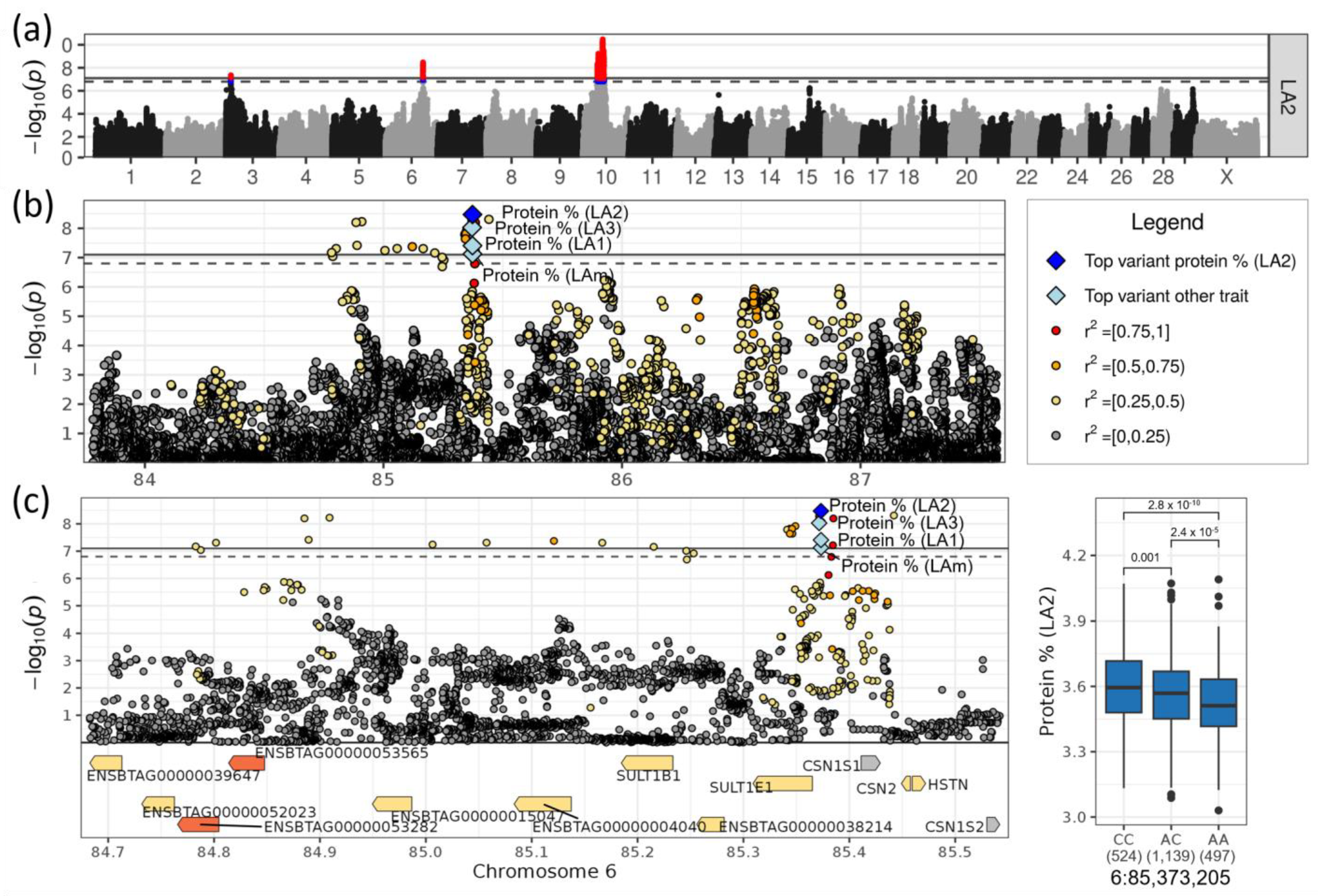
3.4. Loci on Chromosomes 6 and 10 Affected Milk Protein Content
3.5. A Locus on Chromosome 27 Affected Milk Fat Yield
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zentrale Dokumentation Tiergenetischer Ressourcen in Deutschland; B.L.E. Rind: Deutsches Schwarzbuntes Niederungsrind. Available online: https://tgrdeu.genres.de/ (accessed on 1 February 2023).
- Brade, W.; Brade, E. Breeding History of German Holstein Cattle. Ber. Über Landwirtsch. 2013, 91. [Google Scholar]
- Grothe, P.O. Holstein-Friesian: Eine Rasse Geht Um Die Welt; Landwirtschaftsverlag: Münster, Germany, 1993; ISBN 3-7843-2535-1. [Google Scholar]
- Neumann, G.B.; Korkuć, P.; Arends, D.; Wolf, M.J.; May, K.; König, S.; Brockmann, G.A. Genomic Diversity and Relationship Analyses of Endangered German Black Pied Cattle (DSN) to 68 Other Taurine Breeds Based on Whole-Genome Sequencing. Front. Genet. 2023, 13, 993959. [Google Scholar] [CrossRef] [PubMed]
- Korkuć, P.; Arends, D.; May, K.; König, S.; Brockmann, G.A. Genomic Loci Affecting Milk Production in German Black Pied Cattle (DSN). Front. Genet. 2021, 12, 640039. [Google Scholar] [CrossRef]
- Meier, S.; Arends, D.; Korkuć, P.; Neumann, G.B.; Brockmann, G.A. A Genome-Wide Association Study for Clinical Mastitis in the Dual-Purpose German Black Pied Cattle Breed. J. Dairy Sci. 2020, 103, 10289–10298. [Google Scholar] [CrossRef] [PubMed]
- May, K.; Scheper, C.; Brügemann, K.; Yin, T.; Strube, C.; Korkuć, P.; Brockmann, G.A.; König, S. Genome-Wide Associations and Functional Gene Analyses for Endoparasite Resistance in an Endangered Population of Native German Black Pied Cattle. BMC Genom. 2019, 20, 277. [Google Scholar] [CrossRef]
- Matukumalli, L.K.; Lawley, C.T.; Schnabel, R.D.; Taylor, J.F.; Allan, M.F.; Heaton, M.P.; O’Connell, J.; Moore, S.S.; Smith, T.P.L.; Sonstegard, T.S.; et al. Development and Characterization of a High Density SNP Genotyping Assay for Cattle. PLoS ONE 2009, 4, e5350. [Google Scholar] [CrossRef] [Green Version]
- Neumann, G.B.; Korkuć, P.; Arends, D.; Wolf, M.J.; May, K.; Reißmann, M.; Elzaki, S.; König, S.; Brockmann, G.A. Design and Performance of a Bovine 200 k SNP Chip Developed for Endangered German Black Pied Cattle (DSN). BMC Genom. 2021, 22, 905. [Google Scholar] [CrossRef]
- Howard, N.P.; Troggio, M.; Durel, C.-E.; Muranty, H.; Denancé, C.; Bianco, L.; Tillman, J.; van de Weg, E. Integration of Infinium and Axiom SNP Array Data in the Outcrossing Species Malus × Domestica and Causes for Seemingly Incompatible Calls. BMC Genom. 2021, 22, 246. [Google Scholar] [CrossRef] [PubMed]
- Wolf, M.J.; Yin, T.; Neumann, G.B.; Korkuć, P.; Brockmann, G.A.; König, S.; May, K. Genome-wide Association Study Using Whole-genome Sequence Data for Fertility, Health Indicator, and Endoparasite Infection Traits in German Black Pied Cattle. Genes 2021, 12, 1163. [Google Scholar] [CrossRef]
- Wolf, M.J.; Neumann, G.B.; Korkuć, P.; Yin, T.; Brockmann, G.A.; König, S.; May, K. Genetic Evaluations for Endangered Dual-Purpose German Black Pied Cattle Using 50K SNPs, a Breed-Specific 200K Chip, and Whole-Genome Sequencing. J. Dairy Sci. 2023, submitted.
- Korkuć, P.; Arends, D.; Brockmann, G.A. Finding the Optimal Imputation Strategy for Small Cattle Populations. Front. Genet. 2019, 10, 52. [Google Scholar] [CrossRef] [Green Version]
- Browning, B.L.; Zhou, Y.; Browning, S.R. A One-Penny Imputed Genome from Next-Generation Reference Panels. Am. J. Hum. Genet. 2018, 103, 338–348. [Google Scholar] [CrossRef] [Green Version]
- Danecek, P.; Auton, A.; Abecasis, G.; Albers, C.A.; Banks, E.; DePristo, M.A.; Handsaker, R.E.; Lunter, G.; Marth, G.T.; Sherry, S.T.; et al. The Variant Call Format and VCFtools. Bioinformatics 2011, 27, 2156–2158. [Google Scholar] [CrossRef] [PubMed]
- Rosen, B.D.; Bickhart, D.M.; Schnabel, R.D.; Koren, S.; Elsik, C.G.; Zimin, A.; Dreischer, C.; Schultheiss, S.; Hall, R.; Schroeder, S.G.; et al. Modernizing the Bovine Reference Genome Assembly. In Proceedings of the World Congress on Genetics Applied to Livestock Production, Auckland, New Zealand, 11–16 February 2018; p. 802. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.r-project.org/ (accessed on 1 February 2023).
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; De Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A Tool Set for Whole-Genome Association and Population-Based Linkage Analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Lee, S.H.; Goddard, M.E.; Visscher, P.M. GCTA: A Tool for Genome-Wide Complex Trait Analysis. Am. J. Hum. Genet. 2011, 88, 76–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howe, K.L.; Achuthan, P.; Allen, J.; Allen, J.; Alvarez-Jarreta, J.; Ridwan Amode, M.; Armean, I.M.; Azov, A.G.; Bennett, R.; Bhai, J.; et al. Ensembl 2021. Nucleic Acids Res. 2021, 49, D884–D891. [Google Scholar] [CrossRef] [PubMed]
- Raudvere, U.; Kolberg, L.; Kuzmin, I.; Arak, T.; Adler, P.; Peterson, H.; Vilo, J. G:Profiler: A Web Server for Functional Enrichment Analysis and Conversions of Gene Lists (2019 Update). Nucleic Acids Res. 2019, 47, W191–W198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLaren, W.; Gil, L.; Hunt, S.E.; Riat, H.S.; Ritchie, G.R.S.S.; Thormann, A.; Flicek, P.; Cunningham, F. The Ensembl Variant Effect Predictor. Genome Biol. 2016, 17, 122. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.; Henikoff, S.; Ng, P.C. Predicting the Effects of Coding Non-Synonymous Variants on Protein Function Using the SIFT Algorithm. Nat. Protoc. 2009, 4, 1073–1081. [Google Scholar] [CrossRef]
- Castro-Mondragon, J.A.; Riudavets-Puig, R.; Rauluseviciute, I.; Berhanu Lemma, R.; Turchi, L.; Blanc-Mathieu, R.; Lucas, J.; Boddie, P.; Khan, A.; Manosalva Pérez, N.; et al. JASPAR 2022: The 9th Release of the Open-Access Database of Transcription Factor Binding Profiles. Nucleic Acids Res. 2022, 50, D165–D173. [Google Scholar] [CrossRef]
- Tan, G.; Lenhard, B. TFBSTools: An R/Bioconductor Package for Transcription Factor Binding Site Analysis. Bioinformatics 2016, 32, 1555–1556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Z.L.; Park, C.A.; Reecy, J.M. Building a Livestock Genetic and Genomic Information Knowledgebase through Integrative Developments of Animal QTLdb and CorrDB. Nucleic Acids Res. 2019, 47, D701–D710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Wilkins, D. Gggenes: Draw Gene Arrow Maps in “Ggplot2”; R Package Version 0.4.1; Springer: New York, NY, USA, 2020; ISBN 978-3-319-24277-4. [Google Scholar]
- Slowikowski, K. Ggrepel: Automatically Position Non-Overlapping Text Labels with “Ggplot2”; R Package Version 0.9.1; 2021; Available online: https://CRAN.R-project.org/package=gggenes, (accessed on 1 February 2023).
- Kassambara, A. Ggpubr: “ggplot2” Based Publication Ready Plots; R Package Version 0.4.0; 2020; Available online: https://CRAN.R-project.org/package=ggpubr, (accessed on 1 February 2023).
- Lidauer, M.H.; Pösö, J.; Pedersen, J.; Lassen, J.; Madsen, P.; Mäntysaari, E.A.; Nielsen, U.S.; Eriksson, J.-Å.; Johansson, K.; Pitkänen, T.; et al. Across-Country Test-Day Model Evaluations for Holstein, Nordic Red Cattle, and Jersey. J. Dairy Sci. 2015, 98, 1296–1309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muir, B.L.; Kistemaker, G.; Jamrozik, J.; Canavesi, F. Genetic Parameters for a Multiple-Trait Multiple-Lactation Random Regression Test-Day Model in Italian Holsteins. J. Dairy Sci. 2007, 90, 1564–1574. [Google Scholar] [CrossRef] [Green Version]
- Miglior, F.; Sewalem, A.; Jamrozik, J.; Bohmanova, J.; Lefebvre, D.M.; Moore, R.K. Genetic Analysis of Milk Urea Nitrogen and Lactose and Their Relationships with Other Production Traits in Canadian Holstein Cattle. J. Dairy Sci. 2007, 90, 2468–2479. [Google Scholar] [CrossRef] [Green Version]
- Pausch, H.; Emmerling, R.; Schwarzenbacher, H.; Fries, R. A Multi-Trait Meta-Analysis with Imputed Sequence Variants Reveals Twelve QTL for Mammary Gland Morphology in Fleckvieh Cattle. Genet. Sel. Evol. 2016, 48, 14. [Google Scholar] [CrossRef] [Green Version]
- Tribout, T.; Croiseau, P.; Lefebvre, R.; Barbat, A.; Boussaha, M.; Fritz, S.; Boichard, D.; Hoze, C.; Sanchez, M.P. Confirmed Effects of Candidate Variants for Milk Production, Udder Health, and Udder Morphology in Dairy Cattle. Genet. Sel. Evol. 2020, 52, 55. [Google Scholar] [CrossRef]
- Ning, C.; Kang, H.; Zhou, L.; Wang, D.; Wang, H.; Wang, A.; Fu, J.; Zhang, S.; Liu, J. Performance Gains in Genome-Wide Association Studies for Longitudinal Traits via Modeling Time-Varied Effects. Sci. Rep. 2017, 7, 590. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Wurmser, C.; Pausch, H.; Jung, S.; Reinhardt, F.; Tetens, J.; Thaller, G.; Fries, R. Identification and Dissection of Four Major QTL Affecting Milk Fat Content in the German Holstein-Friesian Population. PLoS ONE 2012, 7, e40711. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.; Ma, L.; Prakapenka, D.; VanRaden, P.M.; Cole, J.B.; Da, Y. A Large-Scale Genome-Wide Association Study in U.S. Holstein Cattle. Front. Genet. 2019, 10, 412. [Google Scholar] [CrossRef]
- Sanchez, M.P.; Govignon-Gion, A.; Croiseau, P.; Fritz, S.; Hozé, C.; Miranda, G.; Martin, P.; Barbat-Leterrier, A.; Letaïef, R.; Rocha, D.; et al. Within-Breed and Multi-Breed GWAS on Imputed Whole-Genome Sequence Variants Reveal Candidate Mutations Affecting Milk Protein Composition in Dairy Cattle. Genet. Sel. Evol. 2017, 49, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nayeri, S.; Sargolzaei, M.; Abo-Ismail, M.K.; May, N.; Miller, S.P.; Schenkel, F.; Moore, S.S.; Stothard, P. Genome-Wide Association for Milk Production and Female Fertility Traits in Canadian Dairy Holstein Cattle. BMC Genet. 2016, 17, 75. [Google Scholar] [CrossRef] [Green Version]
- Meredith, B.K.; Kearney, F.J.; Finlay, E.K.; Bradley, D.G.; Fahey, A.G.; Berry, D.P.; Lynn, D.J. Genome-Wide Associations for Milk Production and Somatic Cell Score in Holstein-Friesian Cattle in Ireland. BMC Genet. 2012, 13, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cole, J.B.; Wiggans, G.R.; Ma, L.; Sonstegard, T.S.; Lawlor, T.J.; Crooker, B.A.; Van Tassell, C.P.; Yang, J.; Wang, S.; Matukumalli, L.K.; et al. Genome-Wide Association Analysis of Thirty One Production, Health, Reproduction and Body Conformation Traits in Contemporary U.S. Holstein Cows. BMC Genom. 2011, 12, 408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grisart, B.; Coppieters, W.; Farnir, F.; Karim, L.; Ford, C.; Berzi, P.; Cambisano, N.; Mni, M.; Reid, S.; Simon, P.; et al. Positional Candidate Cloning of a QTL in Dairy Cattle: Identification of a Missense Mutation in the Bovine DGAT1 Gene with Major Effect on Milk Yield and Composition. Genome Res. 2002, 12, 222–231. [Google Scholar] [CrossRef] [Green Version]
- Korkuć, P.; Neumann, G.B.; Arends, D.; Wolf, M.J.; May, K.; König, S.; Brockmann, G.A. Improved Genome-Wide Associations Using a Breed-Specific 200K SNP Chip for German Black Pied (DSN) Cattle. In Proceedings of the World Congress on Genetics Applied to Livestock Production, Rotterdam, The Netherlands, 3–8 July 2022. [Google Scholar]
- Littlejohn, M.D.; Tiplady, K.; Fink, T.A.; Lehnert, K.; Lopdell, T.; Johnson, T.; Couldrey, C.; Keehan, M.; Sherlock, R.G.; Harland, C.; et al. Sequence-Based Association Analysis Reveals an MGST1 EQTL with Pleiotropic Effects on Bovine Milk Composition. Sci. Rep. 2016, 6, 25376. [Google Scholar] [CrossRef]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Tissue-Based Map of the Human Proteome. Science (80-) 2015, 347, 1260419. [Google Scholar] [CrossRef]
- Groza, T.; Gomez, F.L.; Mashhadi, H.H.; Muñoz-Fuentes, V.; Gunes, O.; Wilson, R.; Cacheiro, P.; Frost, A.; Keskivali-Bond, P.; Vardal, B.; et al. The International Mouse Phenotyping Consortium: Comprehensive Knockout Phenotyping Underpinning the Study of Human Disease. Nucleic Acids Res. 2023, 51, D1038–D1045. [Google Scholar] [CrossRef]
- Gross, J.J.; Schwinn, A.C.; Müller, E.; Münger, A.; Dohme-Meier, F.; Bruckmaier, R.M. Plasma Cholesterol Levels and Short-Term Adaptations of Metabolism and Milk Production during Feed Restriction in Early Lactating Dairy Cows on Pasture. J. Anim. Physiol. Anim. Nutr. 2021, 105, 1024–1033. [Google Scholar] [CrossRef]
- Wagner, G.; Fenzl, A.; Lindroos-Christensen, J.; Einwallner, E.; Husa, J.; Witzeneder, N.; Rauscher, S.; Gröger, M.; Derdak, S.; Mohr, T.; et al. LMO3 Reprograms Visceral Adipocyte Metabolism during Obesity. J. Mol. Med. 2021, 99, 1151–1171. [Google Scholar] [CrossRef]
- Sochacka, M.; Karelus, R.; Opalinski, L.; Krowarsch, D.; Biadun, M.; Otlewski, J.; Zakrzewska, M. FGF12 Is a Novel Component of the Nucleolar NOLC1/TCOF1 Ribosome Biogenesis Complex. Cell Commun. Signal. 2022, 20, 182. [Google Scholar] [CrossRef] [PubMed]
- Cyranka, M.; Veprik, A.; McKay, E.J.; van Loon, N.; Thijsse, A.; Cotter, L.; Hare, N.; Saibudeen, A.; Lingam, S.; Pires, E.; et al. Abcc5 Knockout Mice Have Lower Fat Mass and Increased Levels of Circulating GLP-1. Obesity 2019, 27, 1292–1304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powell, D.R.; Revelli, J.P.; Doree, D.D.; Dacosta, C.M.; Desai, U.; Shadoan, M.K.; Rodriguez, L.; Mullens, M.; Yang, Q.M.; Ding, Z.M.; et al. High-Throughput Screening of Mouse Gene Knockouts Identifies Established and Novel High Body Fat Phenotypes. Diabetes Metab. Syndr. Obes. Targets Ther. 2021, 14, 3753–3785. [Google Scholar] [CrossRef] [PubMed]
- Farrell, H.M.; Jimenez-Flores, R.; Bleck, G.T.; Brown, E.M.; Butler, J.E.; Creamer, L.K.; Hicks, C.L.; Hollar, C.M.; Ng-Kwai-Hang, K.F.; Swaisgood, H.E. Nomenclature of the Proteins of Cows’ Milk—Sixth Revision. J. Dairy Sci. 2004, 87, 1641–1674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuss, A.W.; Gogol, J.; Bartenschlager, H.; Geldermann, H. Polymorphic AP-1 Binding Site in Bovine CSN1S1 Shows Quantitative Differences in Protein Binding Associated with Milk Protein Expression. J. Dairy Sci. 2005, 88, 2246–2252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kucerova, J.; Matejicek, A.; Jandurová, O.M.; Sorensen, P.; Nemcova, E.; Stipkova, M.; Kott, T.; Bouska, J.; Frelich, J. Milk Protein Genes CSN1S1, CSN2, CSN3, LGB and Their Relation to Genetic Values of Milk Production Parameters in Czech Fleckvieh. Czech J. Anim. Sci. 2006, 51, 241. [Google Scholar] [CrossRef] [Green Version]
- Caroli, A.M.; Chessa, S.; Erhardt, G.J. Invited Review: Milk Protein Polymorphisms in Cattle: Effect on Animal Breeding and Human Nutrition. J. Dairy Sci. 2009, 92, 5335–5352. [Google Scholar] [CrossRef] [Green Version]
- Meier, S.; Korkuć, P.; Arends, D.; Brockmann, G.A. DNA Sequence Variants and Protein Haplotypes of Casein Genes in German Black Pied Cattle (DSN). Front. Genet. 2019, 10, 1129. [Google Scholar] [CrossRef] [Green Version]
- Chi, N.W.; Lodish, H.F. Tankyrase Is a Golgi-Associated Mitogen-Activated Protein Kinase Substrate That Interacts with IRAP in GLUT4 Vesicles. J. Biol. Chem. 2000, 275, 38437–38444. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait (Lactation) | rsID | Chr | Position | MA | MAF | βMA | SE(βMA) | −log10(p) |
|---|---|---|---|---|---|---|---|---|
| Protein kg (LA3) | rs379781684 | 1 | 75,187,853 | C | 0.15 | −16.8 | 2.7 | 7.96 |
| Fat kg (LA3) | rs379781684 | 1 | 75,187,853 | C | 0.15 | −19.4 | 3.4 | 7.03 |
| Milk kg (LA3) | rs209578598 | 1 | 83,272,783 | A | 0.36 | −349 | 77 | 7.82 |
| Protein % (LA2) | rs876040025 | 3 | 11,592,760 | C | 0.05 | −0.098 | 0.015 | 7.34 |
| Protein % (LA3) | rs41586418 | 3 | 13,915,936 | G | 0.18 | −0.068 | 0.012 | 7.18 |
| Fat % (LAm) | rs134606936 | 5 | 90,406,099 | T | 0.44 | −0.113 | 0.024 | 7.78 |
| Fat % (LA1) | rs211210569 | 5 | 93,516,066 | T | 0.23 | −0.151 | 0.033 | 11.93 |
| Fat % (LA3) | rs209372883 | 5 | 93,518,685 | C | 0.21 | −0.186 | 0.051 | 8.26 |
| Fat % (LA2) | rs207994397 | 5 | 93,525,076 | T | 0.23 | −0.226 | 0.043 | 10.91 |
| Protein % (LA3) | rs41604619 | 5 | 95,098,733 | T | 0.29 | 0.061 | 0.011 | 8.26 |
| Protein % (LA3) | rs382685419 | 6 | 85,371,484 | T | 0.50 | −0.054 | 0.013 | 8.03 |
| Protein % (LA2) | rs378558630 | 6 | 85,373,205 | A | 0.49 | −0.055 | 0.011 | 8.47 |
| Protein % (LA1) | rs378558630 | 6 | 85,373,205 | A | 0.49 | −0.035 | 0.01 | 7.41 |
| Protein % (LAm) | rs378558630 | 6 | 85,373,205 | A | 0.49 | −0.047 | 0.013 | 7.13 |
| Milk kg (LA3) | rs385677618 | 8 | 56534074 | T | 0.33 | −346 | 114 | 8.04 |
| Protein kg (LA3) | rs432948152 | 8 | 56568636 | G | 0.33 | −8.5 | 3.9 | 6.84 |
| Protein kg (LAm) | rs210911072 | 8 | 59917537 | G | 0.23 | −6.4 | 4.6 | 7.53 |
| Milk kg (LAm) | rs797297575 | 8 | 60079367 | G | 0.14 | −280 | 226 | 8.03 |
| Protein % (LA2) | rs211239920 | 10 | 44,746,907 | T | 0.34 | 0.054 | 0.015 | 10.48 |
| Protein % (LA3) | rs208655317 | 10 | 44,746,980 | G | 0.29 | 0.065 | 0.02 | 8.13 |
| Protein % (LAm) | rs208655317 | 10 | 44,746,980 | G | 0.29 | 0.075 | 0.021 | 7.95 |
| Protein % (LA1) | rs137281406 | 20 | 33,314,537 | C | 0.07 | −0.089 | 0.012 | 7.19 |
| Milk kg (LA2) | rs211525696 | 21 | 66,634,034 | A | 0.22 | 338 | 64 | 7.35 |
| Milk kg (LAm) | rs385070122 | 21 | 68,335,947 | A | 0.09 | 486 | 84 | 7.42 |
| Fat kg (LA1) | rs42120938 | 27 | 25,539,379 | A | 0.42 | −9.4 | 2.4 | 8.95 |
| Top SNP | QTL Start | QTL Stop | Length | Gene Names (No. of Genes) |
|---|---|---|---|---|
| 1:75,187,853 | 75,045,795 | 75,397,280 | 0.35 | FGF12 (1) |
| 1:83,272,783 | 83,066,360 | 83,334,688 | 0.27 | HTR3C, s, ABCC5, ENSBTAG00000052825, PARL, MAP6D1, YEATS2 (7) |
| 3:11,592,760 | 11,579,114 | 11,630,952 | 0.05 | CD1E (1) |
| 3:13,915,936 | 13,915,389 | 13,919,145 | 0.01 | ARHGEF11 (1) |
| 5:93,516,066 | 93,300,498 | 93,662,363 | 0.36 | LMO3, MGST1, SLC15A5 (3) |
| 6:85,373,205 | 84,782,659 | 85,442,084 | 0.66 | ENSBTAG00000053282, ENSBTAG00000053565, ENSBTAG00000015047, ENSBTAG00000004040, SULT1B1, ENSBTAG00000038214, SULT1E1, CSN1S1 (8) |
| 8:56,534,074 | 55,472,873 | 56,898,488 | 1.43 | TLE4 (1) |
| 8:60,079,367 | 59,912,308 | 60,102,002 | 0.19 | GBA2, RGP1, MSMP, NPR2, SPAG8, HINT2, FAM221B, TMEM8B, OR13E1, OR13E10, OR13J1C, OR13J1F (12) |
| 10:44,746,907 | 43,852,505 | 47,828,666 | 3.98 | TRIM9, ENSBTAG00000053552, TMX1, FRMD6, GNG2, RTRAF, NID2, ENSBTAG00000001423, PTGDR, PLEKHO2, PIF1, RBPMS2, OAZ2, ZNF609, TRIP4, ENSBTAG00000049725, PCLAF, CSNK1G1, ENSBTAG00000049412, PPIB, SNX22, SNX1, CIAO2A, DAPK2, HERC1, ENSBTAG00000052707, ENSBTAG00000051076, ENSBTAG00000054388, ENSBTAG00000050908, ENSBTAG00000019474, FBXL22, USP3, CA12, APH1B, RAB8B, RPS27L, LACTB, TPM1, ENSBTAG00000040590, TLN2 (40) |
| 20:33,314,537 | 33,313,093 | 33,329,684 | 0.02 | C6 (1) |
| 21:66,634,034 | 66,620,652 | 66,634,034 | 0.01 | - (0) |
| 21:68,335,947 | 68,328,315 | 69,356,368 | 1.03 | ZFYVE21, PPP1R13B, ATP5MJ, TDRD9, RD3L, ASPG, KIF26A, C21H14orf180, TMEM179, ENSBTAG00000054250, ENSBTAG00000007187, ADSS1, SIVA1, AKT1, ZBTB42, CEP170B, PLD4 (17) |
| 27:25,539,379 | 25,489,516 | 25,563,491 | 0.07 | TNKS (1) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Korkuć, P.; Neumann, G.B.; Hesse, D.; Arends, D.; Reißmann, M.; Rahmatalla, S.; May, K.; Wolf, M.J.; König, S.; Brockmann, G.A. Whole-Genome Sequencing Data Reveal New Loci Affecting Milk Production in German Black Pied Cattle (DSN). Genes 2023, 14, 581. https://doi.org/10.3390/genes14030581
Korkuć P, Neumann GB, Hesse D, Arends D, Reißmann M, Rahmatalla S, May K, Wolf MJ, König S, Brockmann GA. Whole-Genome Sequencing Data Reveal New Loci Affecting Milk Production in German Black Pied Cattle (DSN). Genes. 2023; 14(3):581. https://doi.org/10.3390/genes14030581
Chicago/Turabian StyleKorkuć, Paula, Guilherme B. Neumann, Deike Hesse, Danny Arends, Monika Reißmann, Siham Rahmatalla, Katharina May, Manuel J. Wolf, Sven König, and Gudrun A. Brockmann. 2023. "Whole-Genome Sequencing Data Reveal New Loci Affecting Milk Production in German Black Pied Cattle (DSN)" Genes 14, no. 3: 581. https://doi.org/10.3390/genes14030581
APA StyleKorkuć, P., Neumann, G. B., Hesse, D., Arends, D., Reißmann, M., Rahmatalla, S., May, K., Wolf, M. J., König, S., & Brockmann, G. A. (2023). Whole-Genome Sequencing Data Reveal New Loci Affecting Milk Production in German Black Pied Cattle (DSN). Genes, 14(3), 581. https://doi.org/10.3390/genes14030581