
How to Choose? Comparing Different Methods to Count Wolf Packs in a Protected Area of the Northern Apennines

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
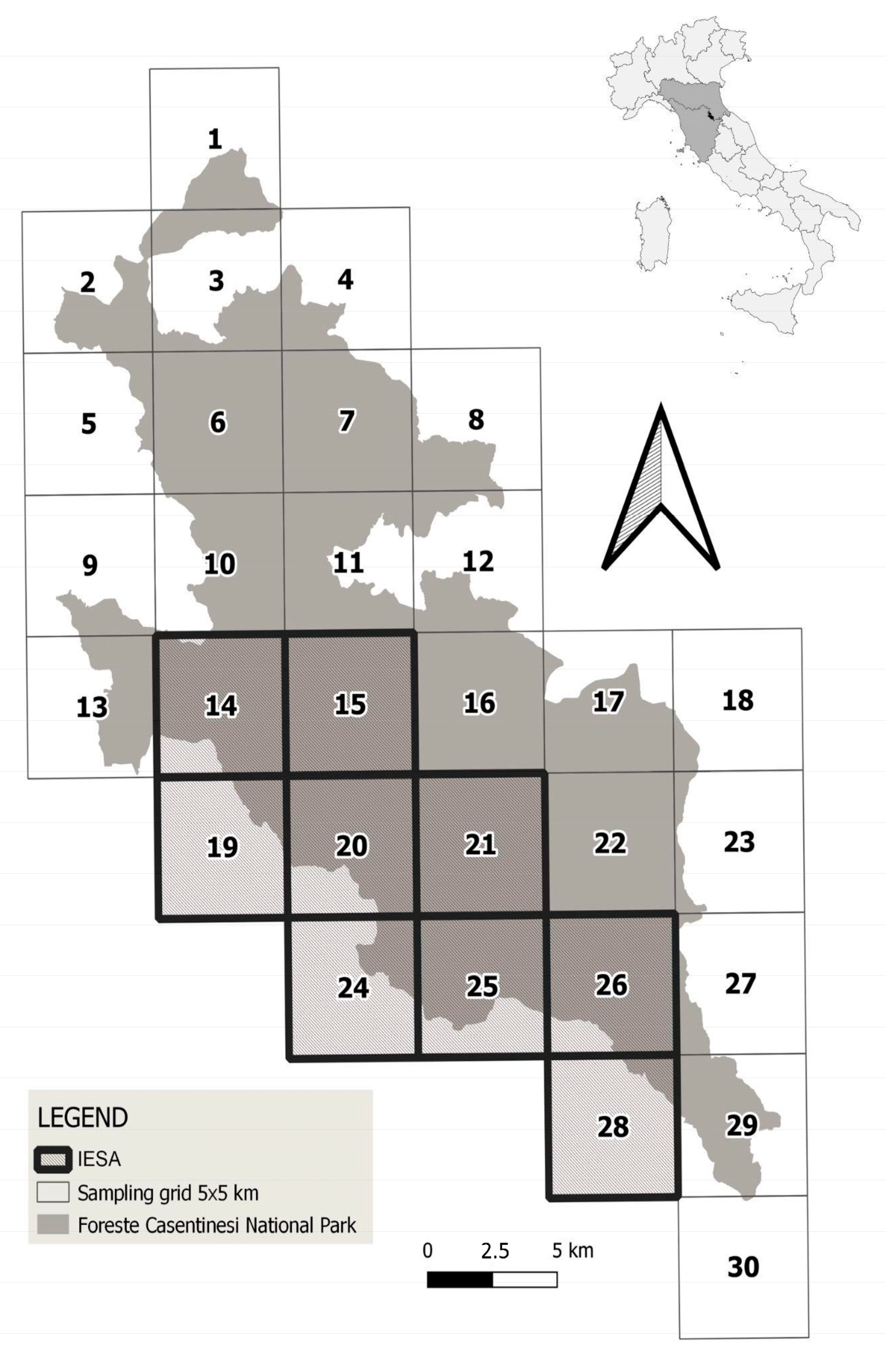
2.1. Study Area
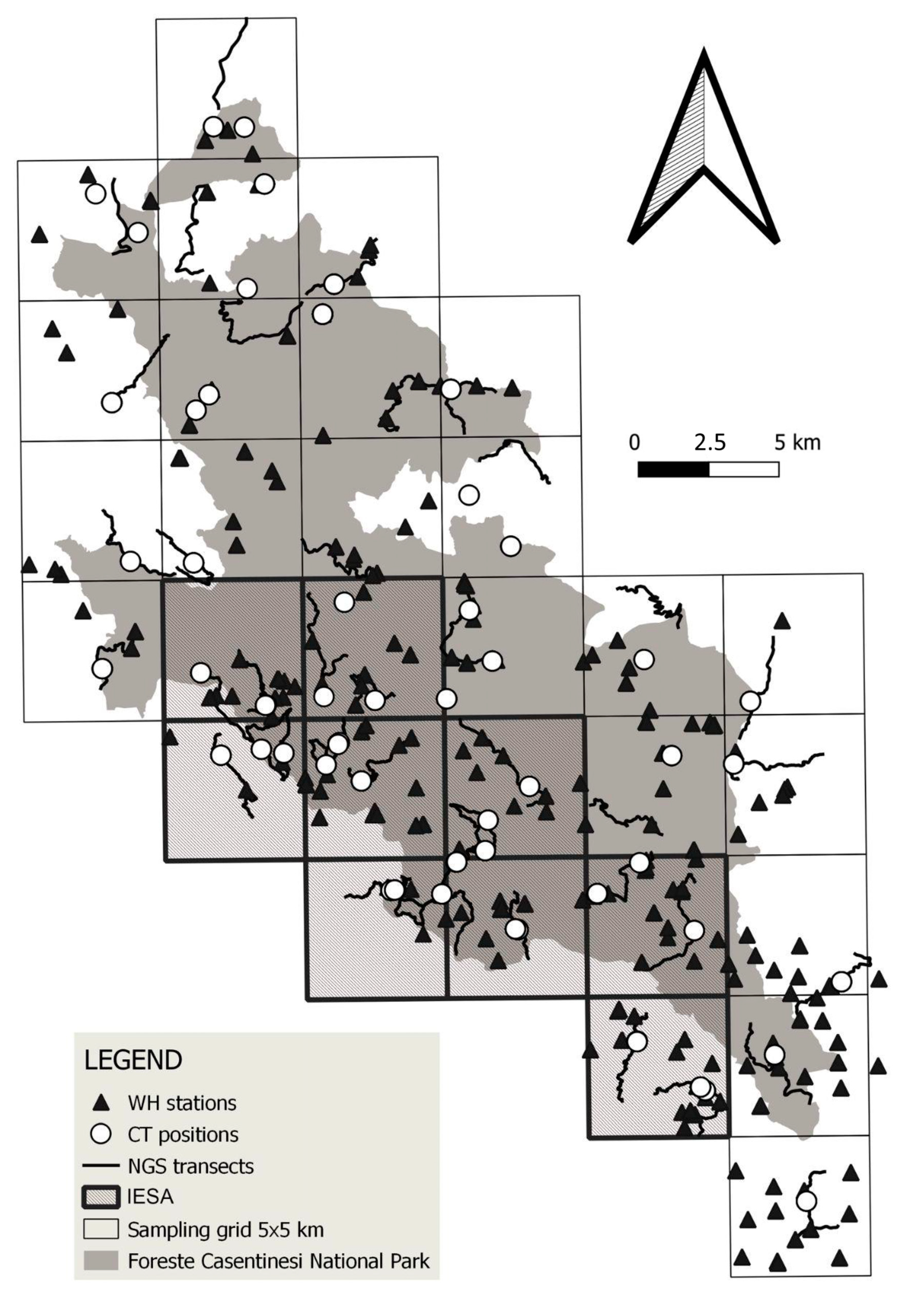
2.2. Survey Design
2.3. Wolf Howling
2.4. Camera Trapping
2.5. Non-Invasive Genetic Sampling
2.6. Comparing and Integrating Methods
2.7. Costs and Working Time
3. Results
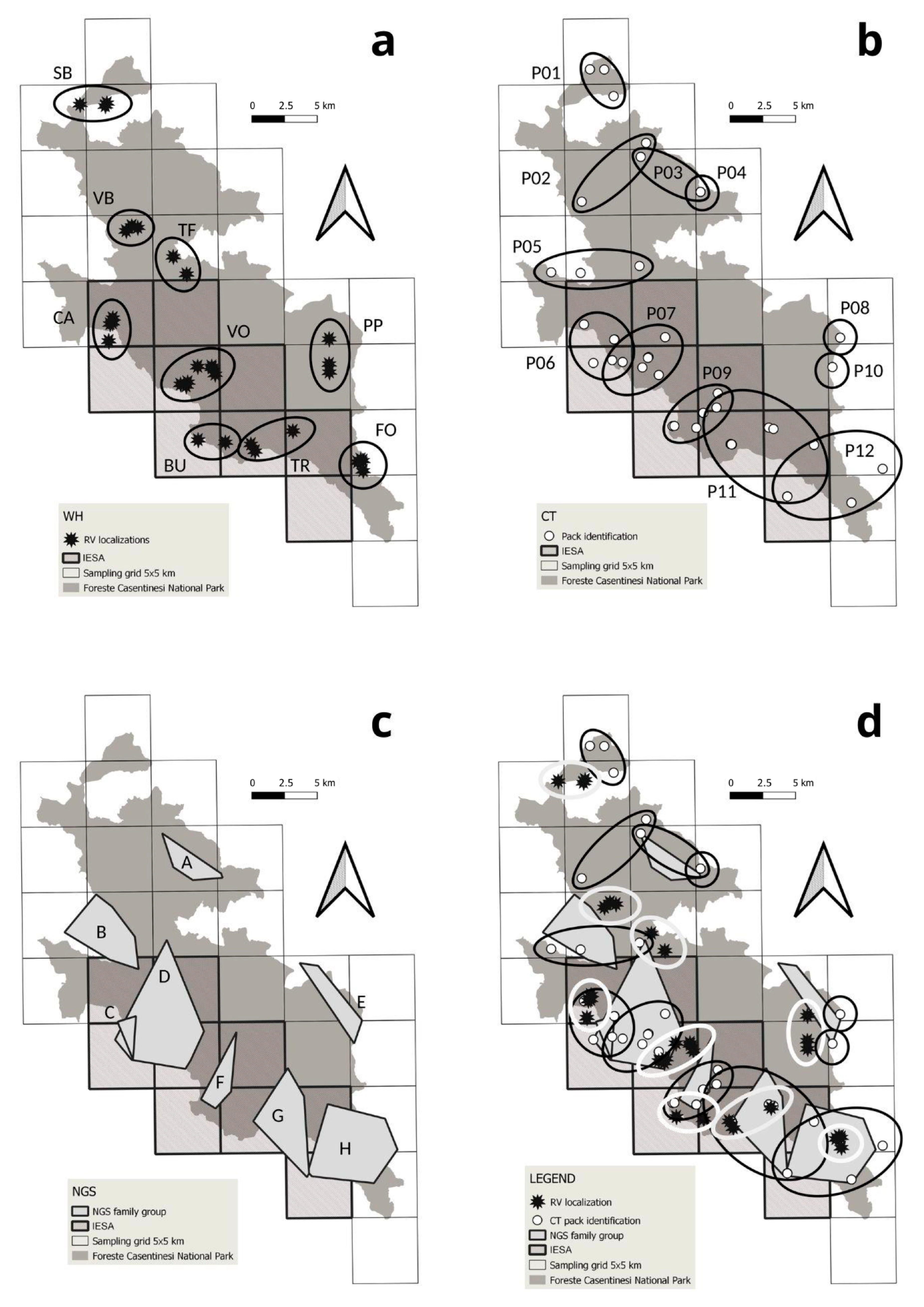
3.1. Wolf Howling
3.2. Camera Trapping
3.3. Non-Invasive Genetic Sampling
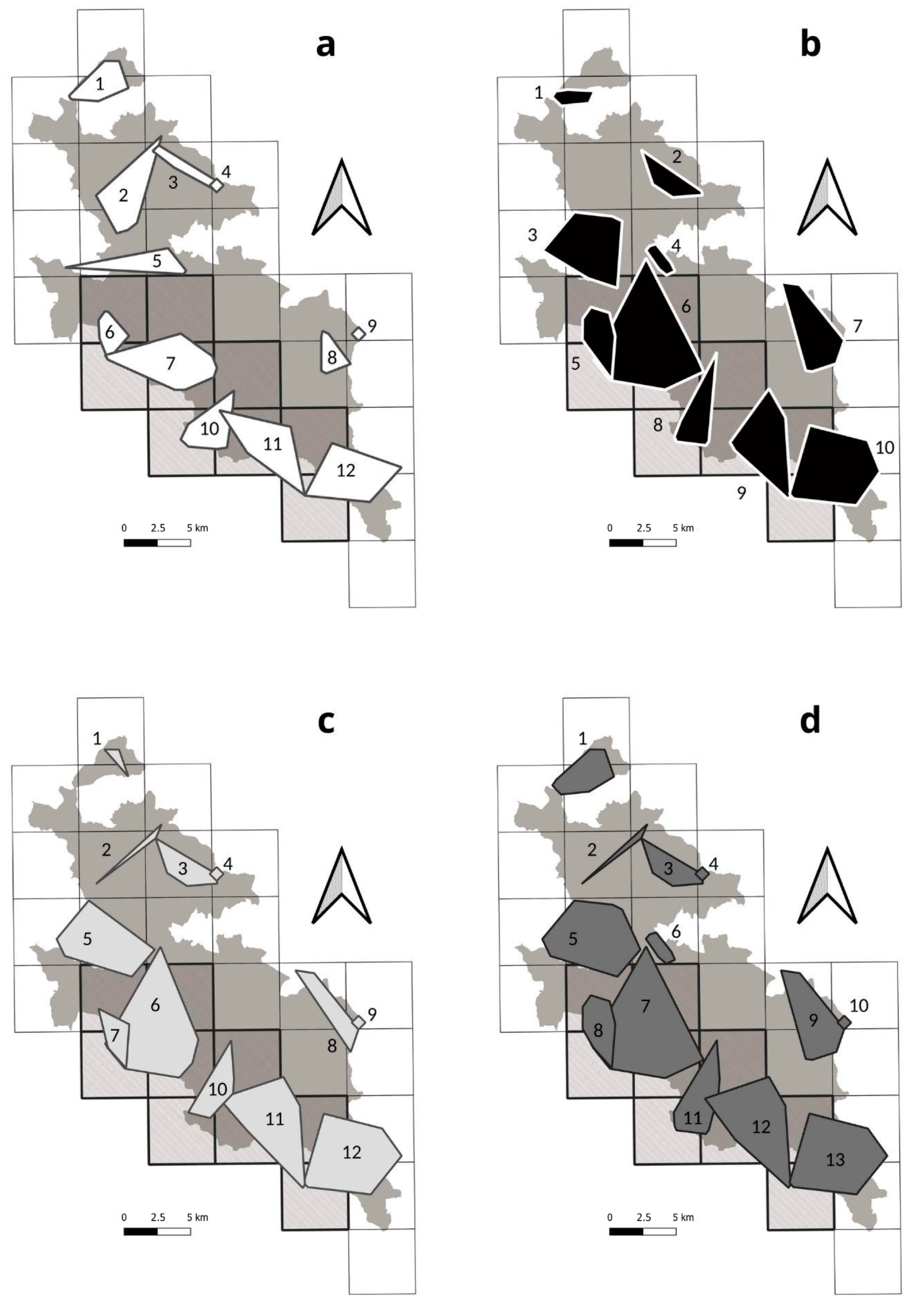
3.4. Integrated Methods
3.5. Costs and Working Time
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Chapron, G.; Kaczensky, P.; von Arx, M.; Huber, D.; Linnell, J.; Boitani, L. Recovery of large carnivores in Europe’s modern human-dominated landscapes. Science 2014, 346, 1517. [Google Scholar] [CrossRef]
- Kusak, J.; Fabbri, E.; Galov, A.; Gomerčić, T.; Arbanasić, H.; Caniglia, R.; Galaverni, M.; Reljić, S.; Huber, D.; Randi, E. Wolf-dog hybridization in Croatia. Vet. Arh. 2018, 88, 375–395. [Google Scholar] [CrossRef]
- Musto, C.; Cerri, J.; Galaverni, M.; Caniglia, R.; Fabbri, E.; Apollonio, M.; Mucci, N.; Bonilauri, P.; Maioli, G.; Fontana, M.C.; et al. Men and wolves: Anthropogenic causes are an important driver of wolf mortality in human-dominated landscapes in Italy. GECCO 2021, 32, 01892. [Google Scholar] [CrossRef]
- Stronen, A.V.; Mattucci, F.; Fabbri, E.; Galaverni, M.; Cocchiararo, B.; Nowak, C.; Godinho, R.; Ruiz-González, A.; Kusak, J.; Skrbinšek, T.; et al. A reduced SNP panel to trace gene flow across southern European wolf populations and detect hybridization with other Canis taxa. Sci. Rep. 2022, 12, 4195. [Google Scholar] [CrossRef]
- Kaczensky, P.; Chapron, G.; von Arx, M.; Huber, D.; Andrén, H.; Linnell, J. Status, Management and Distribution of Large Carnivores-Bear, Lynx, Wolf and Wolverine-in Europe. In A Large Carnivore Initiative for to the Europe Report prepared for the European Commission; Part 1—Europe summaries, report: 1–72; Istituto di Ecologia Applicata: Rome, Italy, 2013. [Google Scholar] [CrossRef]
- Gese, E.M.; Sillero-Zubiri, C.; Hoffmann, M.; Macdonald, D.W. Survey and Census Techniques for Canids. In Canids: Foxes, Wolves, Jackals and Dogs-Status Survey and Conservation Action Plan, IUCN/SSC Canid Specialist Group; IUCN Publications Services Unit: Cambridge, UK, 2004; pp. 273–279. [Google Scholar]
- Kunkel, K.; Mack, C.M.; Melquist, W.E. An Assessment of Current Methods for Surveying and Monitoring Wolves; Nez Perce Tribe: Lapwai, ID, USA, 2005. [Google Scholar]
- Blanco, J.C.; Cortéz, Y. Surveying wolves without snow: A critical review of the methods used in Spain. Hystrix It J. Mamm. 2012, 23, 35–48. [Google Scholar] [CrossRef]
- Llaneza, L.; García, E.J.; López-Bao, J.V. Intensity of Territorial Marking Predicts Wolf Reproduction: Implications for Wolf Monitoring. PLoS ONE 2014, 9, e93015. [Google Scholar] [CrossRef]
- Galaverni, M.; Caniglia, R.; Fabbri, E.; Milanesi, P.; Randi, E. One, no one, or one hundred thousand: How many wolves are there currently in Italy? Mammal Res. 2015, 61, 13–24. [Google Scholar] [CrossRef]
- Palacios, V.; Font, E.; García, E.J.; Svensson, L.; Llaneza, L.; Frank, J.; López-Bao, J.V. Reliability of human estimates of the presence of pups and the number of wolves vocalizing in chorus howls: Implications for decision-making processes. Eur. J. Wildl. Res. 2017, 63, 59. [Google Scholar] [CrossRef]
- Gable, T.D.; Windels, S.K.; Bump, J.K. Finding wolf homesites: Improving the efficacy of howl surveys to study wolves. Peer J. 2018, 6, 5629. [Google Scholar] [CrossRef] [PubMed]
- Canu, A.; Mattioli, L.; Santini, A.; Apollonio, M.; Scandura, M. ‘Video-scats’: Combining camera trapping and non-invasive genotyping to assess individual identity and hybrid status in gray wolf. Wildl. Biol. 2017, 2017, 1–9. [Google Scholar] [CrossRef]
- Marucco, F.; Avanzinelli, E.; Boitani, L. Non-invasive Integrated Sampling Design to Monitor the Wolf Population in Piemonte, Italian Alps. Hystrix It. J. Mamm. 2012, 23, 5–13. [Google Scholar] [CrossRef]
- Balestrieri, A.; Remonti, L.; Prigioni, C. Habitat selection in a low-density badger Meles meles population: A comparison of radio-tracking and latrine surveys. Wildl. Biol. 2009, 15, 442–448. [Google Scholar] [CrossRef]
- Espartosa, K.D.; Trevizan Pinotti, B.; Pardini, R. Performance of camera trapping and track counts for surveying large mammals in rainforest remnants. Biodivers. Conserv. 2011, 20, 2815–2829. [Google Scholar] [CrossRef]
- Monterroso, P.; Rich, L.N.; Serronha, A.; Ferreras, P.; Alves, P.C. Efficiency of hair snares and camera traps to survey mesocarnivore populations. Eur. J. Wildl. Res. 2014, 60, 279–289. [Google Scholar] [CrossRef]
- Velli, E.; Bologna, M.A.; Castelli, S.; Ragni, B.; Randi, E. Non-invasive monitoring of the European wildcat (Felis silvestris silvestris Schreber, 1777): Comparative analysis of three different monitoring techniques and evaluation of their integration. Eur. J. Wildl. Res. 2015, 61, 657–668. [Google Scholar] [CrossRef]
- Torrents-Ticó, M.; Rich, L.; McNutt, J.W.; Nthomiwa, M.; Mothala, M.; Motsamai, G.; Jordan, N.R. On the right track? Comparing concurrent spoor and camera-trap surveys in Botswana. Afr. J. Wildl. Res. 2017, 47, 128–137. [Google Scholar] [CrossRef]
- Cozzi, G.; Hollerbach, L.; Suter, S.M.; Reiners, T.E.; Kunz, F.; Tettamanti, F.; Ozgul, A. Eyes, ears, or nose? Comparison of three non-invasive methods to survey wolf recolonization. Mamm. Biol. 2021, 101, 881–893. [Google Scholar] [CrossRef]
- Apollonio, M.; Mattioli, L.; Scandura, M.; Mauri, L.; Gazzola, A.; Avanzinelli, E. Wolves in the Casentinesi Forests: Insights for wolf conservation in Italy from a protected area with a rich wild prey community. Biol. Conserv. 2004, 120, 249–260. [Google Scholar] [CrossRef]
- Bassi, E.; Willis, S.G.; Passilongo, D.; Mattioli, L.; Apollonio, M. Predicting the Spatial Distribution of Wolf (Canis lupus) Breeding Areas in a Mountainous Region of Central Italy. PLoS ONE 2015, 10, e0124698. [Google Scholar] [CrossRef]
- Mattioli, L.; Canu, A.; Passilongo, D.; Scandura, M.; Apollonio, M. Estimation of pack density in grey wolf (Canis lupus) by applying spatially explicit capture-recapture models to camera trap data supported by genetic monitoring. Front. Zool. 2018, 15, 38. [Google Scholar] [CrossRef] [PubMed]
- Caniglia, R.; Fabbri, E.; Galaverni, M.; Milanesi, P.; Randi, E. Noninvasive sampling and genetic variability, pack structure, and dynamics in an expanding wolf population. J. Mammal. 2014, 95, 41–59. [Google Scholar] [CrossRef]
- Fabbri, E.; Velli, E.; D’Amico, F.; Galaverni, M.; Mastrogiuseppe, M.; Mattucci, F.; Caniglia, R. From predation to management: Monitoring wolf distribution and understanding depredation patterns from attacks on livestock. Hystrix It J. Mamm. 2018, 29, 101–110. [Google Scholar] [CrossRef]
- Galaverni, M.; Palumbo, D.; Fabbri, E.; Caniglia, R.; Greco, C.; Randi, E. Monitoring wolves (Canis lupus) by noninvasive genetics and camera trapping: A small-scale pilot study. Eur. J. Wildl. Res. 2012, 58, 47–58. [Google Scholar] [CrossRef]
- Duchamp, C.; Boyer, J.; Briaudet, P.E.; Leonard, Y.; Moris, P.; Bataille, A.; Dahier, T.; Delacour, G.; Millisher, G.; Miquel, C.; et al. A dual frame survey to assess time- and space-related changes of the colonizing wolf population in France. Hystrix It J. Mamm. 2012, 23, 14–28. [Google Scholar] [CrossRef]
- Chapron, G.; Wikenros, C.; Liberg, O.; Wabakken, P.; Flagstad, Ø.; Milleret, C.; Månsson, J.; Svensson, L.; Zimmermann, B.; Åkesson, M.; et al. Estimating wolf (Canis lupus) population size from number of packs and an individual based model. Ecol. Model. 2016, 339, 33–44. [Google Scholar] [CrossRef]
- Wolf Alpine Group. Wolf Population Status in the Alps: Pack Distribution and Trend Up to 2012. 6p. Available online: http://www.lcie.org (accessed on 16 June 2014).
- Harrington, F.H.; Mech, L.D. An Analysis of howling response parameters useful for wolf pack censusing. J. Wildl. Manag. 1982, 46, 686–693. [Google Scholar] [CrossRef]
- Mech, D.L.; Boitani, L. Wolves: Behavior, Ecology and Conservation; The University of Chicago Press: Chicago, IL, USA, 2003; pp. 31–78. [Google Scholar]
- Noss, A.J.; Cuéllar, R.L.; Barrientos, J.; Maffei, L.; Cuéllar, E.; Arispe, R.; Rúmiz, D.; Rivero, K. A Camera Trapping and Radio Telemetry Study of Lowland Tapir (Tapirus terrestris) in Bolivian Dry Forests. In Tapir Conservation-Newsletter of the IUCN/SSC Tapir Specialist Group; Houston Zoo Inc.: Houston, TX, USA, 2003; Volume 12, pp. 24–32. [Google Scholar]
- Trolle, M.; Noss, A.J.; De Sousa Lima, E.; Dalponte, G.C. Camera-trap studies of maned wolf density in the Cerrado and the Pantanal of Brazil. Biodivers. Conserv. 2007, 16, 1197–1204. [Google Scholar] [CrossRef]
- Se Quin Larrucea, E.; Brussard, P.F.; Jaeger, M.M.; Barrett, R.H. Cameras, Coyotes, and the Assumption of Equal Detectability. J. Wildl. Manag. 2007, 71, 1682–1689. [Google Scholar] [CrossRef]
- Kelly, M.J.; Noss, A.J.; Di Bitetti, M.S.; Maffei, L.; Arispe, R.L.; Paviolo, A.; De Angelo, C.D.; Di Blanco, Y.E. Estimating Puma densities from camera trapping across three study sites: Bolivia, Argentina, and Belize. J. Mammal. 2008, 89, 408–418. [Google Scholar] [CrossRef]
- Sarmento, P.; Cruz, J.; Eira, C.; Fonseca, C. Evaluation of Camera Trapping for Estimating Red Fox Abundance. J. Wildl. Manag. 2009, 73, 1207–1212. [Google Scholar] [CrossRef]
- Linnell, J.D.C.; Swenson, J.E.; Landa, A.; Kvam, T. Methods for monitoring European large carnivores-A worldwide review of relevant experience. NINA Oppdragsmeld. 1998, 549, 1–38. [Google Scholar]
- Marucco, F.; Mattei, L.; Papitto, G.; Bionda, R.; Ramassa, E.; Avanzinelli, E.; Pedrini, P.; Bragalanti, N.; Martinelli, L.; Canavese, G.; et al. Strategia, Metodi e Criteri Per il Monitoraggio Dello Stato di Conservazione Della Popolazione di Lupo Sulle Alpi Italiane. LIFE 12 NAT/IT/00080 WOLFALPS–Action A2. pp. 15–22. 2014. Available online: http://ex.lifewolfalps.eu/download/ (accessed on 5 June 2018).
- Velli, E.; Fabbri, E.; Galaverni, M.; Mattucci, F.; Mattioli, L.; Molinari, L.; Caniglia, R. Ethanol versus swabs: What is a better tool to preserve faecal samples for non-invasive genetic analyses? Hystrix It J. Mamm. 2019, 30, 24–29. [Google Scholar] [CrossRef]
- Taberlet, P.; Waits, L.P.; Luikart, G. Noninvasive genetic sampling: Look before you leap. TREE 1999, 14, 323–327. [Google Scholar] [CrossRef]
- Randi, E.; Hulva, P.; Fabbri, E.; Galaverni, M.; Galov, A.; Kusak, J.; Bigi, D.; Bolfínová, B.C.; Smetanová, M.; Caniglia, R. Multilocus Detection of Wolf x Dog Hybridization in Italy, and Guidelines for Marker Selection. PLoS ONE 2014, 9, e86409. [Google Scholar] [CrossRef]
- Caniglia, R.; Galaverni, M.; Velli, E.; Mattucci, F.; Canu, A.; Appollonio, M.; Mucci, N.; Scandura, M.; Fabbri, E. A standardized approach to empirically define reliable assignment thresholds and appropriate management categories in deeply introgressed populations. Sci. Rep. 2020, 10, 2862. [Google Scholar] [CrossRef] [PubMed]
- Sundqvist, A.K.; Ellegren, H.; Olivier, M.; Vilà, C. Y chromosome haplotyping in Scandinavian wolves (Canis lupus) based on microsatellite markers. Mol. Ecol. 2001, 10, 1959–1966. [Google Scholar] [CrossRef] [PubMed]
- Candille, S.I.; Kaelin, C.B.; Cattanach, B.M.; Yu, B.; Thompson, D.A.; Nix, M.A.; Kerns, J.A.; Schmutz, S.M.; Millhauser, G.L.; Barsh, G.S. A b-defensin mutation causes black coat color in domestic dogs. Science 2007, 318, 1418–1423. [Google Scholar] [CrossRef]
- Caniglia, R.; Fabbri, E.; Greco, C.; Galaverni, M.; Manghi, L.; Boitani, L.; Sforzi, A.; Randi, E. Black coats in an admixed wolf × dog pack is melanism an indicator of hybridization in wolves? Eur. J. Wildl. Res. 2013, 59, 543–555. [Google Scholar] [CrossRef]
- Valière, N. GIMLET: A computer program for analysing genetic individual identification data. Mol. Ecol. Notes 2002, 2, 377–379. [Google Scholar] [CrossRef]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Linked loci and correlated allele frequencies. Genetics 2003, 164, 1567–1587. [Google Scholar] [CrossRef]
- Verardi, A.; Lucchin, V.; Randi, E. Detecting introgressive hybridization between free-ranging domestic dogs and wild wolves (Canis lupus) by admixture linkage disequilibrium analysis. Mol. Ecol. 2006, 15, 2845–2855. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Santure, A.W. Parentage and Sibship Inference from Multilocus Genotype Data under Polygamy. Genetics 2009, 181, 1579–1594. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef]
- Marucco, F.; Boitani, L.; Pletscher, D.H.; Schwartz, M.K. Bridging the gaps between non-invasive genetic sampling and population parameter estimation. Eur. J. Wildl. Res. 2011, 57, 1–13. [Google Scholar] [CrossRef]
- Gazzola, A.; Avanzinelli, E.; Mauri, L.; Scandura, M.; Apollonio, M. Temporal changes of howling in South European wolf packs. Ital. J. Zool. 2002, 69, 157–161. [Google Scholar] [CrossRef]
- Llaneza, L.; Ordiz, A.; Palacios, V.; Uzal, A. Monitoring Wolf Populations Using Howling Points Combined with Sign Survey Transects. Wildl. Biol. Pract. 2005, 1, 108–117. [Google Scholar] [CrossRef]
- Iliopoulos, Y.; Youlatos, D.; Sgardelis, S. Wolf pack rendezvous site selection in Greece is mainly affected by anthropogenic landscape features. Eur. J. Wildl. Res. 2013, 60, 23–24. [Google Scholar] [CrossRef]
- Meek, P.D.; Ballard, G.A.; Falzon, G. The higher you go the less you will know: Placing camera traps high to avoid theft will affect detection. Remote Sens. Ecol. Conserv. 2016, 2, 204–211. [Google Scholar] [CrossRef]
- Marucco, F.; Pletscher, D.H.; Boitani, L.; Schwartz, M.K.; Pilgrim, K.L.; Lebreton, J. Wolf survival and population trend using non-invasive capture–recapture techniques in the Western Alps. J. Appl. Ecol. 2009, 46, 1003–1010. [Google Scholar] [CrossRef]
- Caniglia, R.; Fabbri, E.; Cubaynes, S.; Gimenez, O.; Lebreton, J.; Randi, E. An improved procedure to estimate wolf abundance using non-invasive genetic sampling and capture–recapture mixture models. Conserv. Genet. 2012, 13, 53–64. [Google Scholar] [CrossRef]
- Rigg, R.; Skrbinšek, T.; Linnell, J. Engaging hunters and other stakeholders in a pilot study of wolves in Slovakia using non-invasive genetic sampling. In Final Report to the EU Commission; Istituto di Ecologia Applicata: Rome, Italy, 2014. [Google Scholar]
- Ausband, D.E.; Lukacs, P.M.; Hurley, M.; Roberts, S.; Strickfaden, K.; Moeller, A.K. Estimating wolf abundance from cameras. Ecosphere 2022, 13, 3933. [Google Scholar] [CrossRef]
- Granroth-Wilding, H.; Primmer, C.; Lindqvist, M.; Poutanen, J.; Thalmann, O.; Aspi, J.; Harmoinen, J.; Kojola, I.; Laaksonen, T. Non-invasive genetic monitoring involving citizen science enables reconstruction of current pack dynamics in a re-establishing wolf population. BMC Ecol. 2017, 17, 44. [Google Scholar] [CrossRef] [PubMed]
- Vonholdt, B.M.; Stahler, D.R.; Smith, D.W.; Earl, D.A.; Pollinger, J.P.; Wayne, R.K. The genealogy and genetic viability of reintroduced Yellowstone grey wolves. Mol. Ecol. 2007, 17, 252–274. [Google Scholar] [CrossRef]
- Jaraush, A.; Harms, V.; Kluth, J.; Reinhardt, I.; Nowak, C. How the west was won: Genetic reconstruction of rapid wolf recolonization into Germany’s anthropogenic landscapes. Heredity 2021, 127, 92–106. [Google Scholar] [CrossRef] [PubMed]
- Santostasi, N.L.; Gimenez, O.; Caniglia, R.; Fabbri, E.; Molinari, L.; Reggioni, W.; Ciucci, P. Estimating Admixture at the Population Scale: Taking Imperfect Detectability and Uncertainty in Hybrid Classification Seriously. J. Wildl. Manag. 2021, 85, 1031–1046. [Google Scholar] [CrossRef]
- Lorenzini, R.; Fanelli, R.; Grifoni, G.; Scholl, F.; Fico, R. Wolf–dog crossbreeding: “Smelling” a hybrid may not be easy. Mamm. Biol. 2014, 79, 149–156. [Google Scholar] [CrossRef]
- Galaverni, M.; Caniglia, R.; Pagani, L.; Fabbri, E.; Boattini, A.; Randi, E. Disentangling Timing of Admixture, Patterns of Introgression, and Phenotypic Indicators in a Hybridizing Wolf Population. Mol. Biol. Evol. 2017, 34, 2324–2339. [Google Scholar] [CrossRef]
- Godinho, R.; Lòpez-Bao, J.V.; Castro, D.; Llaneza, L.; Lopes, S.; Silva, P.; Ferrand, N. Real-time assessment of hybridization between wolves and dogs: Combining noninvasive samples with ancestry informative markers. Mol. Ecol. Resour. 2014, 15, 317–328. [Google Scholar] [CrossRef]
- Hindrikson, M.; Männil, P.; Ozolins, J.; Krzywinski, A.; Saarma, U. Bucking the Trend in Wolf-Dog Hybridization: First Evidence from Europe of Hybridization between Female Dogs and Male Wolves. PLoS ONE 2012, 7, e46465. [Google Scholar] [CrossRef]
- Benson, J.F.; Patterson, B.R. Inter-specific territoriality in a Canis hybrid zone: Spatial segregation between wolves, coyotes, and hybrids. Oecologia 2013, 173, 1539–1550. [Google Scholar] [CrossRef]
- Ciucci, P.; Boitani, L.; Falco, M.; Maiorano, L. Hierarchical, Multi-Grain Rendezvous Site Selection by Wolves in Southern Italy. J. Wildl. Manag. 2018, 82, 1049–1061. [Google Scholar] [CrossRef]
- Corcoran, E.; Winsen, M.; Sudholz, A.; Hamilton, G. Automated detection of wildlife using drones: Synthesis, opportunities and constraints. Methods Ecol. Evol. 2021, 12, 1103–1114. [Google Scholar] [CrossRef]
- Harmoinen, J.; von Thaden, A.; Aspi, J.; Kvist, L.; Cocchiararo, B.; Jaraush, A.; Gazzola, A.; Sin, T.; Lohi, H.; Hytönen, M.K.; et al. Reliable wolf-dog hybrid detection in Europe using a reduced SNP panel developed for non-invasively collected samples. BMC Genom. 2021, 22, 473. [Google Scholar] [CrossRef]
- Stronen, A.V.; Aspi, J.; Caniglia, R.; Fabbri, E.; Galaverni, M.; Godinho, R.; Kvist, L.; Mattucci, F.; Nowak, C.; von Thaden, A.; et al. Wolf-dog admixture highlights the need for methodological standards and multidisciplinary cooperation for effective governance of wild x domestic hybrids. Biol. Conserv. 2022, 266, 109467. [Google Scholar] [CrossRef]
- Hassan, S.; Bhat, S.; Poczai, P.; Ganai, B.A.; Almalki, W.H.; Gafur, A.; Sayyed, R.Z. Environmental DNA Metabarcoding: A Novel Contrivance for Documenting Terrestrial Biodiversity. Biology 2022, 11, 1297. [Google Scholar] [CrossRef] [PubMed]
- Mills, L.S.; Citta, J.J.; Lair, K.P.; Schwartz, M.K.; Tallmon, D.A. Estimating animal abundance using non-invasive DNA sampling: Promise and pitfalls. Ecol. Appl. 2000, 10, 283–294. [Google Scholar] [CrossRef]
- Waits, L.P.; Luikart, G.; Taberlet, P. Estimating the probability of identity among genotypes in natural populations: Cautions and guidelines. Mol. Ecol. 2001, 10, 249–256. [Google Scholar] [CrossRef]
- Randi, E.; Lucchini, V.; Fjeldsø Christensen, M.; Mucci, N.; Funk, S.M.; Dolf, G.; Loeschcke, V. Mitochondrial DNA variability in Italian and east European wolf: Detecting the consequence of small population size and hybridization. Conserv. Biol. 2000, 14, 464–473. [Google Scholar] [CrossRef]
- Iacolina, L.; Scandura, M.; Gazzola, A.; Cappai, N.; Capitani, C.; Mattioli, L.; Vercillo, F.; Apollonio, M. Y-chromosome microsatellite variation in Italian wolves: A contribution to the study of wolf–dog hybridization patterns. Mamm. Biol. 2010, 75, 341–347. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software Structure: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef]
- Randi, E. Detecting hybridization between wild species and their domesticated relatives. Mol. Ecol. 2008, 17, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, E.E.; Bach, L.A.; Kotlicki, P. Hybridlab (version 1.0): A program for generating simulated hybrids from population samples. Mol. Ecol. Notes 2006, 6, 971–973. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2019 | 2020 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| May | Jun | Jul | Aug | Sep | Oct | Nov | Dec | Jan | Feb | Mar | Apr |
| WH | WH | WH | WH | ||||||||
| CT | CT | CT | CT | CT | CT * | CT * | CT * | CT * | CT | ||
| NGS | NGS | NGS | NGS | NGS | NGS | NGS | NGS * | NGS * | NGS * | NGS * | NGS |
| Area Coverage | Packs | Effort | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Technique | % Cells with Sampling Results * | % Cells with Packs Identification | N° Packs | Packs Rated as A | Costs (€) | Total Work Days | ||||
| WH | 36.7 | 55.6 | 40 | 55.6 | 9 | 4 | - | - | 720.96 | 72 |
| CT | 93.3 | 100 | 73.3 | 100 | 12 | 5 | - | - | 12,168.80 | 412 |
| NGS | 100 | 100 | 66.7 | 100 | 8 | 5 | - | - | 29,400.00 | 506 |
| WH + CT | 93.3 | 100 | 83.3 | 100 | 12 | 5 | 4 | 4 | 12,889.76 | 484 |
| WH + NGS | 100 | 100 | 73.3 | 100 | 10 | 5 | 3 | 3 | 30,120.96 | 578 |
| CT + NGS | 100 | 100 | 80 | 100 | 12 | 5 | 6 | 5 | 41,568.80 | 606 ** |
| WH + CT + NGS | 100 | 100 | 93.3 | 100 | 13 | 5 | 6 | 5 | 42,289.76 | 678 ** |
| Wolf Howling (WH) | Camera Trapping (CT) | ||||||
|---|---|---|---|---|---|---|---|
| Pack ID | N° Localizations (WH) | Distance (km) | Detection Days | N° Localizations (CT) | Distance (km) | Detection Days | N° Puppies (CT) |
| SB | 2 | 2.06 | 24/07/2019 04/09/2019 | - | - | - | - |
| VB | 1 | - | 26/09/2019 | - | - | - | - |
| TF | 2 | 1.68 | 19/08/2019 09/09/2019 | - | - | - | - |
| CA | 2 | 1.78 | 01/08/2019 07/08/2019 | 2 | 1.94 | From 25/09/2019 to 30/09/2019 | 4 |
| PP | 2 | 2.51 | 03/08/2019 27/08/2019 05/09/2019 | - | - | - | - |
| VO | 2 | 2.75 | 23/07/2019 19/08/2019 | - | - | - | - |
| BU | 2 | 2.09 | 29/07/2019 22/08/2019 | 1 | - | From 29/08/2019 to 18/09/2019 | 4 |
| TR | 2 | 3.36 | 31/07/2019 15/08/2019 23/08/2019 | 2 | 3.41 | From 02/08/2019 to 28/09/2019 | 4 |
| FO | 1 | - | 25/07/2019 | 1 | - | From 19/08/2019 to 28/09/2019 | 4 |
| Average | 1.8 | 2.32 | - | 1.5 | 2.67 | - | 4 |
| Camera Trapping (CT) | |||||||
|---|---|---|---|---|---|---|---|
| Pack ID | N° Videos | Pack Size | N° Females Detected | N° Males Detected | First Detection | Last Detection | N° Focal Individuals |
| P01 | 12 | 6 | 3 | 1 | 03/12/2019 | 20/02/2020 | 2 |
| P02 | 34 | 8 | 1 | 1 | 25/11/2019 | 02/04/2020 | 2 |
| P03 | 22 | 3 | 1 | 1 | 04/12/2019 | 19/03/2020 | 1 |
| P04 | 31 | 8 | 2 | 1 | 01/12/2019 | 12/03/2020 | 3 |
| P05 | 19 | 9 | - | - | 21/12/2019 | 05/03/2020 | 4 |
| P06 | 9 | 9 | 1 | 1 | 12/12/2019 | 29/03/2020 | 2 |
| P07 | 25 | 6 | 2 | 3 | 25/11/2019 | 02/04/2020 | 3 |
| P08 | 18 | 6 | 2 | 1 | 21/11/219 | 08/02/2020 | 5 |
| P09 | 70 | 7 | 2 | 1 | 07/12/2019 | 07/04/2020 | 1 |
| P10 | 13 | 3 | 1 | 1 | 21/01/2020 | 30/03/2020 | 0 |
| P11 | 76 | 12 | 3 | 2 | 03/12/2019 | 01/04/2020 | 1 |
| P12 | 68 | 9 | 2 | 2 | 24/11/2019 | 15/03/2020 | 0 |
| Average | 33.1 | 7.2 | 1.8 | 1.4 | - | - | 2 |
| Non-Invasive Genetic Sampling (NGS) | ||||||
|---|---|---|---|---|---|---|
| Pack ID | Pack Size | Reproductive Couple | N° Offspring | N° Female Offspring | N° Male Offspring | N° Dispersers |
| A | 4 | HFO3M + WFO313F | 2 | 1 | 1 | 0 |
| B | 5 | WFO229M + WFO239F | 4 | 1 | 3 | 1 |
| C | 3 | WFO204M + WFO315F | 3 | 3 | 0 | 2 |
| D | 6 | WFO288M + WFO249F | 4 | 1 | 3 | 0 |
| E | 4 | WFO278M + WFO282F | 2 | 1 | 1 | 0 |
| F | 5 | WFO251M + WFO205F | 3 | 3 | 0 | 0 |
| G | 8 | WFO232M + WFO206F | 6 | 4 | 2 | 0 |
| H | 7 | HFO1M + WFO256F | 6 | 5 | 1 | 1 |
| Average | 5.3 | - | 3.8 | 2.4 | 1.4 | 0.5 |
| N° Packs | WH + CT | Grade | WH + NGS | Grade | CT + NGS | Grade | WH + CT + NGS | Grade |
|---|---|---|---|---|---|---|---|---|
| 1 | SB + P01 | (B) | SB + -- | (C) | P01 + -- | (C) | SB + P01 + -- | (B) |
| 2 | VB + P02 | (B) | -- + A | (C) | P02 + -- | (C) | -- + P02 + -- | (C) |
| 3 | -- + P03 | (C) | VB + B | (B) | P03 + A | (B) | -- + P03 + A | (B) |
| 4 | -- + P04 | (C) | TF + -- | (C) | P04 + -- | (C) | -- + P04 + -- | (C) |
| 5 | TF + P05 | (B) | CA + C | (B) | P05 + B | (A) | VB + P05 + B | (A) |
| 6 | CA + P06 | (A) | VO + D | (A) | P06 + C | (A) | TF + -- + -- | (C) |
| 7 | VO + P07 | (A) | PP + E | (B) | P07 + D | (A) | VO + P07 + D | (A) |
| 8 | PP + P10 | (B) | BU + F | (B) | P10 + E | (B) | CA + P06 + C | (A) |
| 9 | -- + P08 | (C) | TR + G | (A) | P08 + -- | (C) | PP + P10 + E | (B) |
| 10 | BU + P09 | (B) | FO + H | (A) | P09 + F | (A) | -- + P08 + -- | (C) |
| 11 | TR + P11 | (A) | -- + -- | -- | P11 + G | (A) | BU + P09 + F | (A) |
| 12 | FO + P12 | (A) | -- + -- | -- | P12 + H | (A) | TR + P11 + G | (A) |
| 13 | -- + -- | -- | -- + -- | -- | -- + -- | -- | FO + P12 + H | (A) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dissegna, A.; Rota, M.; Basile, S.; Fusco, G.; Mencucci, M.; Cappai, N.; Galaverni, M.; Fabbri, E.; Velli, E.; Caniglia, R. How to Choose? Comparing Different Methods to Count Wolf Packs in a Protected Area of the Northern Apennines. Genes 2023, 14, 932. https://doi.org/10.3390/genes14040932
Dissegna A, Rota M, Basile S, Fusco G, Mencucci M, Cappai N, Galaverni M, Fabbri E, Velli E, Caniglia R. How to Choose? Comparing Different Methods to Count Wolf Packs in a Protected Area of the Northern Apennines. Genes. 2023; 14(4):932. https://doi.org/10.3390/genes14040932
Chicago/Turabian StyleDissegna, Arianna, Martino Rota, Simone Basile, Giuseppe Fusco, Marco Mencucci, Nadia Cappai, Marco Galaverni, Elena Fabbri, Edoardo Velli, and Romolo Caniglia. 2023. "How to Choose? Comparing Different Methods to Count Wolf Packs in a Protected Area of the Northern Apennines" Genes 14, no. 4: 932. https://doi.org/10.3390/genes14040932
APA StyleDissegna, A., Rota, M., Basile, S., Fusco, G., Mencucci, M., Cappai, N., Galaverni, M., Fabbri, E., Velli, E., & Caniglia, R. (2023). How to Choose? Comparing Different Methods to Count Wolf Packs in a Protected Area of the Northern Apennines. Genes, 14(4), 932. https://doi.org/10.3390/genes14040932