
Genetic Diversity and Population Differentiation of a Chinese Endangered Plant Ammopiptanthus nanus (M. Pop.) Cheng f.
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Genetic Diversity Analysis
3.2. Gene Flow
3.3. Genetic Distance and Genetic Similarity among Populations
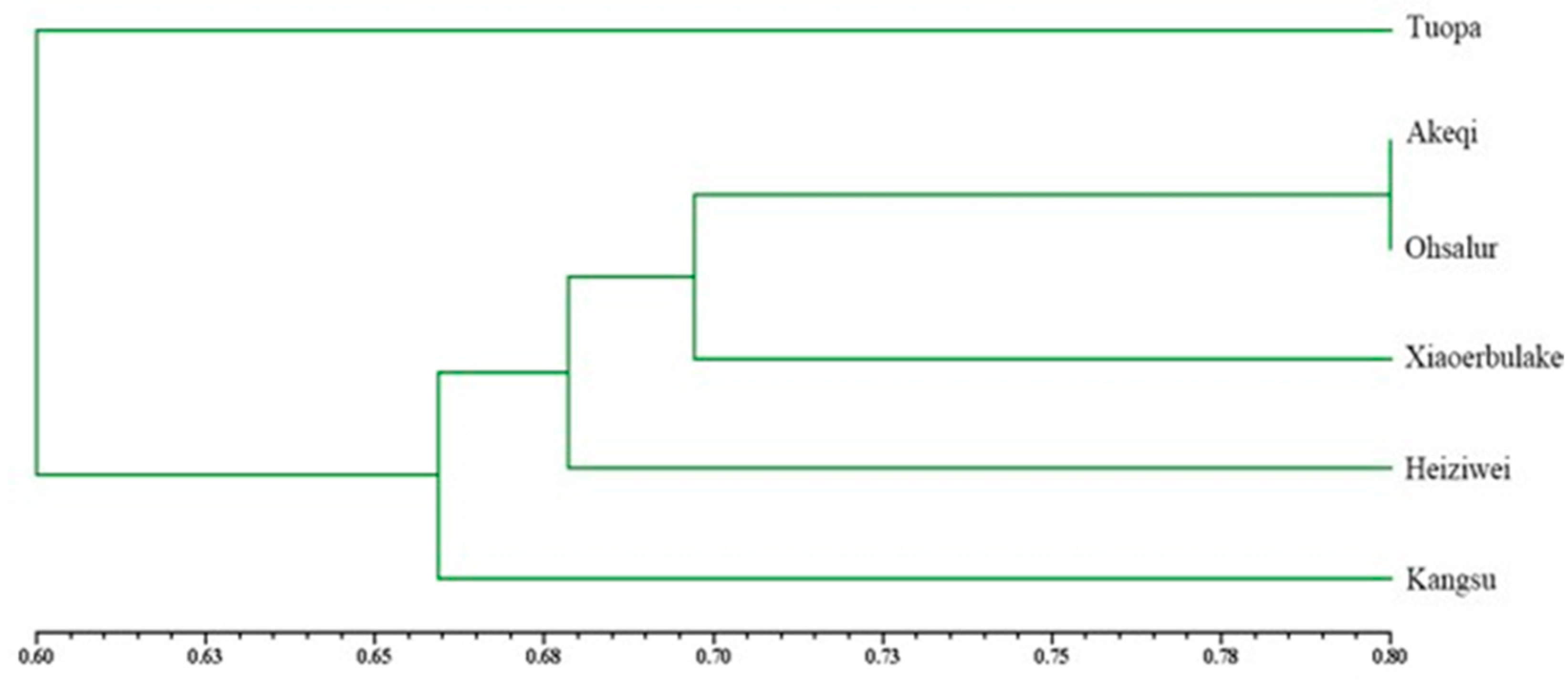
3.4. UPGMA Clustering
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Brown, W.L. Genetic diversity and genetic vulnerability—An appraisal. Econ. Bot. 1983, 37, 4–12. [Google Scholar] [CrossRef]
- Salgotra, R.K.; Chauhan, B.S. Genetic Diversity, Conservation, and Utilization of Plant Genetic Resources. Genes 2023, 14, 174. [Google Scholar] [CrossRef]
- Scribner, K.T.; Uhrig, G.; Kanefsky, J.; Sard, N.M.; Holtgren, M.; Jerome, C.; Ogren, S. Pedigree-based decadal estimates of lake sturgeon adult spawning numbers and genetic diversity of stream-side hatchery produced offspring. J. Great Lakes Res. 2022, 48, 551–564. [Google Scholar] [CrossRef]
- Soltis, P.; Soltis, U. Genetic Variation in Endemic and Widespread Plant Species. Aliso J. Syst. Florist. Bot. 1991, 13, 215–223. [Google Scholar] [CrossRef]
- Xiao, Y.; Jiang, X.; Lu, C.; Liu, J.; Diao, S.; Jiang, J. Genetic Diversity and Population Structure Analysis in the Chinese Endemic Species Michelia crassipes Based on SSR Markers. Forests 2023, 14, 508. [Google Scholar] [CrossRef]
- Kim, M.J.; Jeong, S.Y.; Kim, S.-S.; Jeong, J.S.; Kim, J.S.; Jeong, H.C.; Kim, K.-G.; Kim, I. Population genetic characterization of the endangered dung beetle Copris tripartitus (Coleoptera: Scarabaeidae) using novel microsatellite markers. J. Asia Pac. Èntomol. 2022, 25, 101899. [Google Scholar] [CrossRef]
- Borah, R.; Bhattacharjee, A.; Rao, S.R.; Kumar, V.; Sharma, P.; Upadhaya, K.; Choudhury, H. Genetic diversity and population structure assessment using molecular markers and SPAR approach in Illicium griffithii, a medicinally important endangered species of Northeast India. J. Genet. Eng. Biotechnol. 2021, 19, 118. [Google Scholar] [CrossRef]
- Sandamal, S.; Tennakoon, A.; Meng, Q.; Marambe, B.; Ratnasekera, D.; Melo, A.; Ge, S. Population genetics and evolutionary history of the wild rice species Oryza rufipogon and O. nivara in Sri Lanka. Ecol. Evol. 2018, 8, 12056–12065. [Google Scholar] [CrossRef]
- Wang, Y.; Cao, S.; Sui, X.; Wang, J.; Geng, Y.; Gao, F.; Zhou, Y. Genome-Wide Characterization, Evolution, and Expression Analysis of the Ascorbate Peroxidase and Glutathione Peroxidase Gene Families in Response to Cold and Osmotic Stress in Ammopiptanthus nanus. J. Plant Growth Regul. 2022, 42, 502–522. [Google Scholar] [CrossRef]
- Schaal, B.A.; Hayworth, D.A.; Olsen, K.M.; Rauscher, J.T.; Smith, W.A. Phylogeographic studies in plants: Problems and prospects. Mol. Ecol. 1998, 7, 465–474. [Google Scholar] [CrossRef]
- Wen, L. The effects of genetic diversity from habitat fragment. Sci. Econ. Soc. 2006, 24, 70–72. [Google Scholar]
- Wang, Z.; Peng, S.; Ren, H. Genetic variation and inbreeding depression in small populations. J. Plant Genet. Resour. 2005, 6, 101–107. [Google Scholar] [CrossRef]
- Ruping, W.; Lin, L. The extinction vortex of small population. J. Biol. 2008, 25, 14–16–24. [Google Scholar]
- Primack, B.R. Essentials of Conservation Biology, 5th ed.; Sinauer Associates, Inc.: Sunderland, MA, USA, 2010. [Google Scholar]
- Cheng, S.H. Ammopiptanthus Cheng f. A new genus of Leguminosae from central Asia. Bot. Zhurnal 1959, 44, 1381–1386. [Google Scholar]
- Li, Y. Evergreen shrubs Ammopiptanthus nanus (M. Pop.) Cheng f. research report. Xinjiang For. Sci. Technol. 1989, 1, 55–58. [Google Scholar]
- Pan, B.; Yu, Q.; Yna, C. Study for the ecological environment and vulnerable reasons of the Ammopiptanthus nanus. Acta Phytoecol. Geobot. Sin. 1992, 16, 276–282. [Google Scholar]
- Sun, Y.; Liu, L.; Sun, S.; Han, W.; Irfan, M.; Zhang, X.; Zhang, L.; Chen, L. AnDHN, a Dehydrin Protein from Ammopiptanthus nanus, Mitigates the Negative Effects of Drought Stress in Plants. Front. Plant Sci. 2021, 12, 788938. [Google Scholar] [CrossRef]
- Yu, H.; Zheng, H.; Liu, Y.; Yang, Q.; Li, W.; Zhang, Y.; Fu, F. Antifreeze protein from Ammopiptanthus nanus functions in temperature-stress through domain A. Sci. Rep. 2021, 11, 8458. [Google Scholar] [CrossRef]
- Zhu, M.; Liu, Q.; Liu, F.; Zheng, L.; Bing, J.; Zhou, Y.; Gao, F. Gene Profiling of the Ascorbate Oxidase Family Genes under Osmotic and Cold Stress Reveals the Role of AnAO5 in Cold Adaptation in Ammopiptanthus nanus. Plants 2023, 12, 677. [Google Scholar] [CrossRef]
- Liu, Q.; Sui, X.; Wang, Y.; Zhu, M.; Zhou, Y.; Gao, F. Genome-Wide Analyses of Thaumatin-like Protein Family Genes Reveal the Involvement in the Response to Low-Temperature Stress in Ammopiptanthus nanus. Int. J. Mol. Sci. 2023, 24, 2209. [Google Scholar] [CrossRef]
- Ji, T.-F.; Li, J.; Liang, C.-H. The Chemical constituents of the twigs of Ammopiptanthus nanus. J. Asian Nat. Prod. Res. 2013, 15, 332–336. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Wu, S.; Pan, B.; Wang, D. Geographical distribution and habitat characteristic of [Ammopiptanthus (Maxim.) Cheng f.] (Fabaceae) in China. Arid. Land Geogr. 2017, 40, 380–387. [Google Scholar] [CrossRef]
- Wang, Z. Resource survey of rare plant dwarf sand holly and its conservation. Chin. Wild Plant Resour. 2005, 6, 44–45. [Google Scholar]
- Zhang, Y.Z.; Pan, B.R.; Yin, L.K.; Duan, S.M. Study on the floristic genera and structure of the community of Ammopiptanthus nanus. Arid. Zone Res. 2006, 23, 320–326. [Google Scholar] [CrossRef]
- Ge, X.-J.; Yu, Y.; Yuan, Y.-M.; Huang, H.-W.; Yan, C. Genetic Diversity and Geographic Differentiation in Endangered Ammopiptanthus (Leguminosae) Populations in Desert Regions of Northwest China as Revealed by ISSR Analysis. Ann. Bot. 2005, 95, 843–851. [Google Scholar] [CrossRef]
- Chen, G.; Huang, H.; Ge, X. Allozyme diversity and population differentiation in an endangered plant, Ammopiptanthus nanus (Leguminosae). J. Wuhan Bot. Res. 2005, 23, 131–137. [Google Scholar]
- Zhao, P.; Yong, X.; Hu, G.; Lv, T.; Jiao, P. RAPD analysis on the genetic diversity of endangered plant Ammopiptanthus nanus (M. Pop.) Cheng f. J. Arid. Land Resour. Environ. 2016, 30, 74–79. [Google Scholar] [CrossRef]
- Prevost, A.; Wilkinson, M.J. A new system of comparing PCR primers applied to ISSR fingerprinting of potato cultivars. Theor. Appl. Genet. 1999, 98, 107–112. [Google Scholar] [CrossRef]
- Ali, Z.; Xu, Z.; Zhang, D.; He, X.; Bahadur, S.; Yi, J. Molecular diversity analysis of eggplant (Solanum melongena) genetic resources. Genet. Mol. Res. 2011, 10, 1141–1155. [Google Scholar] [CrossRef]
- Tantasawat, P.A.; Poolsawat, O.; Arsakit, K.; Papan, P. Identification of ISSR, ISSR-RGA and SSR Markers Associated with Cercospora Leaf Spot Resistance Gene in Mungbean. Int. J. Agric. Biol. 2020, 23, 447–453. [Google Scholar] [CrossRef]
- Vaishnav, K.; Tiwari, V.; Durgapal, A.; Meena, B.; Rana, T.S. Estimation of genetic diversity and population genetic structure in Gymnema sylvestre (Retz.) R. Br. ex Schult. populations using DAMD and ISSR markers. J. Genet. Eng. Biotechnol. 2023, 21, 42. [Google Scholar] [CrossRef]
- Slatkin, M. Gene Flow and the Geographic Structure of Natural Populations. Science 1987, 236, 787–792. [Google Scholar] [CrossRef]
- Boydak, M.; Teker, T.; Gazdağli, A.; Thanos, C.A.; Çalişkan, S.; Kaltsis, A.; Tozlu, E.; Fournaraki, C.; Albayrak, G. ISSR genotyping of Phoenix theophrasti natural populations in Turkey and Crete (Greece) and P. dactylifera. Nord. J. Bot. 2021, 39, 1–11. [Google Scholar] [CrossRef]
- Ghomi, K.; Rabiei, B.; Sabouri, H.; Alamdari, E.G. Association analysis, genetic diversity and population structure of barley (Hordeum vulgare L.) under heat stress conditions using SSR and ISSR markers linked to primary and secondary metabolites. Mol. Biol. Rep. 2021, 48, 6673–6694. [Google Scholar] [CrossRef] [PubMed]
- Sheikh, Z.N.; Sharma, V.; Shah, R.A.; Sharma, N.; Summuna, B.; Al-Misned, F.A.; Serehy, H.A.E.; Mir, J.I. Genetic diversity analysis and population structure in apricot (Prunus armeniaca L.) grown under north-western himalayas using ISSR markers. Saudi J. Biol. Sci. 2021, 28, 5986–5992. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Pan, B.; Zhang, Y.; Duan, S. Analysis on the characteristics of communities of Ammopiptanthus nanus and A. mongolicus. Arid. Zone Res. 2007, 24, 487–494. [Google Scholar] [CrossRef]
- Govindaraju, D.R. Relationship between Dispersal Ability and Levels of Gene Flow in Plants. Oikos 1988, 52, 31. [Google Scholar] [CrossRef]
- Moonsap, P.; Laksanavilat, N.; Sinumporn, S.; Tasanasuwan, P.; Kate-Ngam, S.; Jantasuriyarat, C. Genetic diversity of Indo-China rice varieties using ISSR, SRAP and InDel markers. J. Genet. 2019, 98, 80. [Google Scholar] [CrossRef]
- Parab, A.R.; Lynn, C.B.; Subramaniam, S. Assessment of Genetic Stability on In Vitro And Ex Vitro Plants of Ficus Carica Var. Black Jack Using Issr and Damd Markers. Mol. Biol. Rep. 2021, 48, 7223–7231. [Google Scholar] [CrossRef]
- Li, X. The Reproductive Biology of Ammopiptanthus Cheng f. (Fabaceae); Xinjiang Agricultural University: Urumqi, China, 2006. [Google Scholar]
- Jiao, P.; Li, Z. Study on poopulation biology of endangered species Ammmopiptanthus nanus (M. Pop.) Cheng f. J. Anhui Agric. Sci. 2010, 38, 355–360. [Google Scholar] [CrossRef]
- Jixian, W. Save rare plants Ammopiptanthus nanus (M. Pop.) Cheng f. For. People 1995, 1, 27. [Google Scholar]
- Liu, H.; Niu, T.; Yu, Q.; Yang, L.; Ma, J.; Qiu, S.; Wang, R.; Liu, W.; Li, J. Spatial and temporal variations in the relationship between the topological structure of eco-spatial network and biodiversity maintenance function in China. Ecol. Indic. 2022, 139, 108919. [Google Scholar] [CrossRef]
- Sun, X.; Xiao, Y. Vegetation Growth Trends of Grasslands and Impact Factors in the Three Rivers Headwater Region. Land 2022, 11, 2201. [Google Scholar] [CrossRef]
- Chaiyarat, R.; Wettasin, M.; Youngpoy, N.; Cheachean, N. Use of Human Dominated Landscape as Connectivity Corridors among Fragmented Habitats for Wild Asian Elephants (Elephas maximus) in the Eastern Part of Thailand. Diversity 2023, 15, 6. [Google Scholar] [CrossRef]
- Xiao, Y.; Xiao, Q.; Zhang, J. Balancing the international benefits and risks associated with implementation of ecological policy on the Qinghai-Tibet Plateau, China. Gondwana Res. 2023, 115, 183–190. [Google Scholar] [CrossRef]
- Karstens, S.; Dorow, M.; Bochert, R.; Stybel, N.; Schernewski, G.; Mühl, M. Stepping Stones Along Urban Coastlines—Improving Habitat Connectivity for Aquatic Fauna with Constructed Floating Wetlands. Wetlands 2022, 42, 76. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Collection Site | Longitude | Latitude | Altitude (m) | Number of Samples |
|---|---|---|---|---|
| Tuopa | 75°34′54.62″ | 39°46′26.11″ | 2018 | 20 |
| Akeqi | 75°35′24.26″ | 39°49′54.45″ | 2059 | 20 |
| Heiziwei | 75°18′58.45″ | 39°48′9.34″ | 2402 | 20 |
| Kangsu | 75°03′22.00″ | 39°42′12.00″ | 2161 | 20 |
| Ohsalur | 74°45′18.00″ | 39°39′30.00″ | 2202 | 20 |
| Xiaoerbulak | 75°00′53.41″ | 39°41′47.35″ | 2217 | 20 |
| Primers | Strios Number | Polymorphic Band | Percentage of Polymorphic Bands | Primer Sequences (From 5′ End to 3′ End) | Fire Retardant Temperature |
|---|---|---|---|---|---|
| UBC807 | 16 | 9 | 56.25% | AGA GAG AGA GAG AGA GT | 53 °C |
| UBC810 | 15 | 8 | 53.33% | GAG AGA GAG AGA GAG AT | 50 °C |
| UBC811 | 16 | 10 | 62.50% | GAG AGA GAG AGA GAG AC | 45 °C |
| UBC822 | 6 | 3 | 50.00% | TCT CTC TCT CTC TCT CA | 50 °C |
| UBC834 | 13 | 7 | 53.85% | AGAGAGAGAGAGAGAGYT | 49 °C |
| UBC835 | 13 | 6 | 46.15% | AGAGAGAGAGAGAGAGYC | 52 °C |
| UBC840 | 14 | 8 | 57.14% | GAGAGA GAG AGA GAG AYT | 50 °C |
| UBC844 | 15 | 8 | 53.33% | CTC TCT CTC TCT CTC TRC | 49 °C |
| UBC888 | 14 | 5 | 35.71% | BDB CAC ACA CAC ACA CA | 41 °C |
| UBC891 | 14 | 6 | 42.86% | HVH TGT GTG TGT GTG TG | 55 °C |
| Total | 136 | 70 | 51.47% |
| Population/Species | P (%) | NA | NE | H | I | |
|---|---|---|---|---|---|---|
| Population | Tuopa | 22.79 | 1.23 ± 0.42 ab | 1.16 ± 0.32 a | 0.09 ± 0.17 ab | 0.13 ± 0.25 ab |
| Akeqi | 43.38 | 1.43 ± 0.50 b | 1.26 ± 0.35 b | 0.15 ± 0.20 c | 0.23 ± 0.28 c | |
| Heiziwei | 31.62 | 1.32 ± 0.47 b | 1.17 ± 0.29 a | 0.11 ± 0.17 b | 0.16 ± 0.25 b | |
| Kangsu | 28.68 | 1.29 ± 0.45 b | 1.16 ± 0.30 a | 0.09 ± 0.17 ab | 0.14 ± 0.24 ab | |
| Ohsalur | 17.65 | 1.18 ± 0.38 a | 1.10 ± 0.24 a | 0.06 ± 0.14 a | 0.09 ± 0.21 a | |
| Xiaoerbulak | 16.91 | 1.17 ± 0.38 a | 1.12 ± 0.30 a | 0.07 ± 0.16 ab | 0.10 ± 0.23 ab | |
| Mean | 26.84 | 1.27 | 1.16 | 0.10 | 0.14 | |
| Species | 51.70 | 1.99 | 1.60 | 0.35 | 0.52 | |
| Ht | Hs | Gst | Nm |
|---|---|---|---|
| 0.35 ± 0.02 | 0.10 ± 0.01 | 0.73 | 0.19 |
| Populations | Tuopa | Akeqi | Heiziwei | Kangsu | Ohsalur | Xiaoerbulak |
|---|---|---|---|---|---|---|
| Tuopa | ****** | 0.610 | 0.622 | 0.613 | 0.561 | 0.618 |
| Akeqi | 0.495 | ****** | 0.669 | 0.674 | 0.804 | 0.729 |
| Heiziwei | 0.475 | 0.402 | ****** | 0.647 | 0.681 | 0.701 |
| Kangsu | 0.489 | 0.395 | 0.435 | ****** | 0.672 | 0.663 |
| Ohsalur | 0.578 | 0.218 | 0.385 | 0.397 | ****** | 0.675 |
| Xiaoerbulak | 0.481 | 0.316 | 0.356 | 0.411 | 0.393 | ****** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, A.; Ma, M.; Li, H.; He, S.; Wang, S. Genetic Diversity and Population Differentiation of a Chinese Endangered Plant Ammopiptanthus nanus (M. Pop.) Cheng f. Genes 2023, 14, 1020. https://doi.org/10.3390/genes14051020
Li A, Ma M, Li H, He S, Wang S. Genetic Diversity and Population Differentiation of a Chinese Endangered Plant Ammopiptanthus nanus (M. Pop.) Cheng f. Genes. 2023; 14(5):1020. https://doi.org/10.3390/genes14051020
Chicago/Turabian StyleLi, Aoran, Miao Ma, Haotian Li, Songfeng He, and Shugao Wang. 2023. "Genetic Diversity and Population Differentiation of a Chinese Endangered Plant Ammopiptanthus nanus (M. Pop.) Cheng f." Genes 14, no. 5: 1020. https://doi.org/10.3390/genes14051020
APA StyleLi, A., Ma, M., Li, H., He, S., & Wang, S. (2023). Genetic Diversity and Population Differentiation of a Chinese Endangered Plant Ammopiptanthus nanus (M. Pop.) Cheng f. Genes, 14(5), 1020. https://doi.org/10.3390/genes14051020