
Appressoria Formation in Phytopathogenic Fungi Suppressed by Antimicrobial Peptides and Hybrid Peptides from Black Soldier Flies

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bioinformatics Analysis of AMPs from the BSF
2.2. Rearing of the BSF Larvae
2.3. Artificial Synthesis of the Screened CAD Peptides
2.4. Antifungal Activity Assays on M. oryzae and C. acutatum
2.5. Expression Pattern Analysis of the Selected Target Peptides by RT-qPCR
2.6. Cloning and Expression Pattern Analysis of the High-Antifungal-Activity Target Peptides
2.7. CAD-Con Chimeric Vector Construction and Expression in E. coli
2.8. Pathogenicity Assays on Pepper and Rice
2.9. Statistical Analysis
3. Results
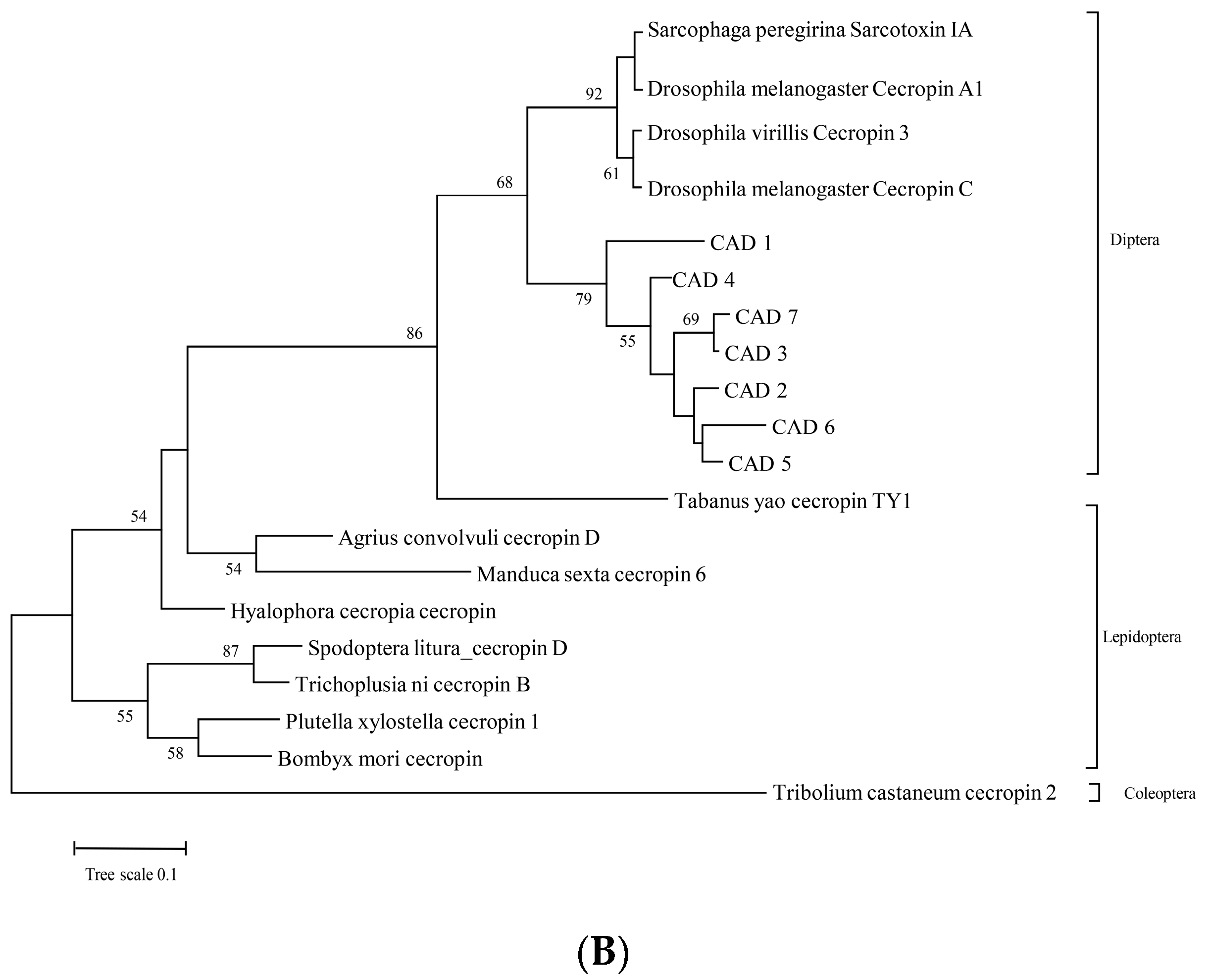
3.1. Amino Acid Sequence Analysis of Cecropin-like Proteins as AMPs from BSF
3.2. CAD1, CAD5, and CAD7 with Antifungal Activity by the Suppression of Appressorium Formation in C. acutatum and M. oryzae
3.3. Expression Patterns of the Selected Peptides from BSF Larvae at Different Instar Ages and under Different Treatment Conditions
3.4. Prokaryotic Expression of the Hybrid Peptide CAD-Con
3.5. Hybrid Peptide CAD-Con Had Stronger Antifungal Activity
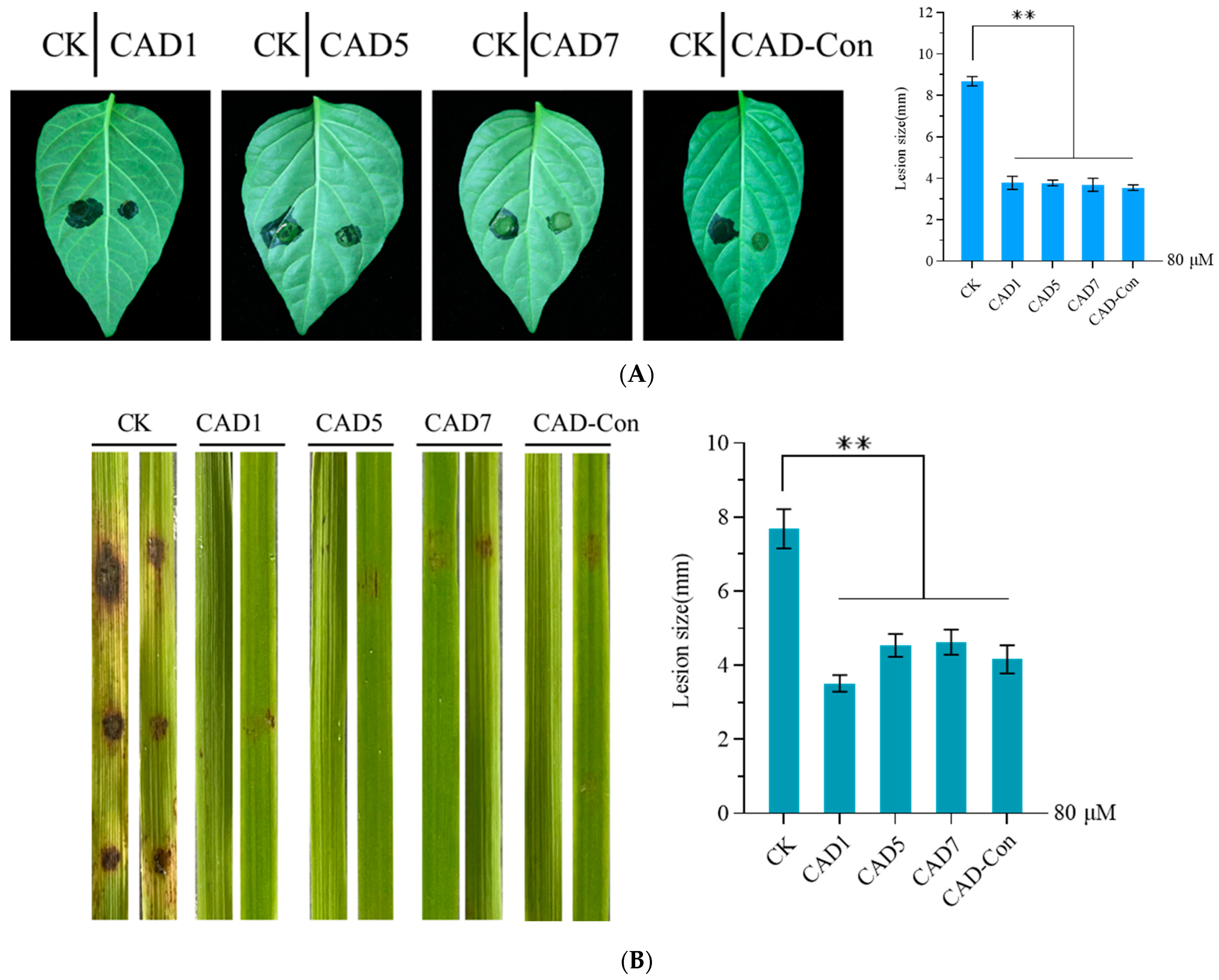
3.6. Pathogenicity Was Reduced by the Conidia of C. acutatum and M. oryzae Treated with CAD1, CAD5, CAD7, or CAD-Con
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, J.; Dou, X.; Song, J.; Lyu, Y.; Zhu, X.; Xu, L.; Li, W.; Shan, A. Antimicrobial Peptides: Promising Alternatives in the Post Feeding Antibiotic Era. Med. Res. Rev. 2019, 39, 831–859. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Haney, E.F.; Vogel, H.J. The Expanding Scope of Antimicrobial Peptide Structures and Their Modes of Action. Trends Biotechnol. 2011, 29, 464–472. [Google Scholar] [CrossRef]
- Boman, H.G. Antibacterial Peptides: Key Components Needed in Immunity. Cell 1991, 65, 205–207. [Google Scholar] [CrossRef] [PubMed]
- Tam, J.P.; Wang, S.; Wong, K.H.; Tan, W.L. Antimicrobial Peptides from Plants. Pharmaceuticals 2015, 8, 711–757. [Google Scholar] [CrossRef] [PubMed]
- Egorov, T.A.; Odintsova, T.I. Defense peptides of plant immune system. Bioorg. Khim. 2012, 38, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Gao, A.G.; Hakimi, S.M.; Mittanck, C.A.; Wu, Y.; Woerner, B.M.; Stark, D.M.; Shah, D.M.; Liang, J.; Rommens, C.M. Fungal Pathogen Protection in Potato by Expression of a Plant Defensin Peptide. Nat. Biotechnol. 2000, 18, 1307–1310. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, S.; Kato, K.; Koba, A.; Minami, Y.; Watanabe, K.; Yagi, F. Purification, Characterization, and Sequencing of Antimicrobial Peptides, Cy-AMP1, Cy-AMP2, and Cy-AMP3, from the Cycad (Cycas Revoluta) Seeds. Peptides 2008, 29, 2110–2117. [Google Scholar] [CrossRef]
- Terras, F.R.; Eggermont, K.; Kovaleva, V.; Raikhel, N.V.; Osborn, R.W.; Kester, A.; Rees, S.B.; Torrekens, S.; Van Leuven, F.; Vanderleyden, J. Small Cysteine-Rich Antifungal Proteins from Radish: Their Role in Host Defense. Plant Cell 1995, 7, 573–588. [Google Scholar] [CrossRef]
- Ohshima, M.; Mitsuhara, I.; Okamoto, M.; Sawano, S.; Nishiyama, K.; Kaku, H.; Natori, S.; Ohashi, Y. Enhanced Resistance to Bacterial Diseases of Transgenic Tobacco Plants Overexpressing Sarcotoxin IA, a Bactericidal Peptide of Insect. J. Biochem. 1999, 125, 431–435. [Google Scholar] [CrossRef]
- Coca, M.; Peñas, G.; Gómez, J.; Campo, S.; Bortolotti, C.; Messeguer, J.; Segundo, B.S. Enhanced Resistance to the Rice Blast Fungus Magnaporthe Grisea Conferred by Expression of a Cecropin A Gene in Transgenic Rice. Planta 2006, 223, 392–406. [Google Scholar] [CrossRef] [PubMed]
- DeGray, G.; Rajasekaran, K.; Smith, F.; Sanford, J.; Daniell, H. Expression of an Antimicrobial Peptide via the Chloroplast Genome to Control Phytopathogenic Bacteria and Fungi. Plant Physiol. 2001, 127, 852–862. [Google Scholar] [CrossRef]
- Mitsuhara, I.; Matsufuru, H.; Ohshima, M.; Kaku, H.; Nakajima, Y.; Murai, N.; Natori, S.; Ohashi, Y. Induced Expression of Sarcotoxin IA Enhanced Host Resistance against Both Bacterial and Fungal Pathogens in Transgenic Tobacco. Mol. Plant Microbe Interact 2000, 13, 860–868. [Google Scholar] [CrossRef] [PubMed]
- Insect Antimicrobial Peptides and Their Applications—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/24811407/ (accessed on 6 March 2023).
- Zhang, Q.; Chen, G.; Shao, L.; Gu, X.; Huang, J.; Peng, C. The Hybridization between Sphagneticola Trilobata (L.) Pruski and Sphagneticola Calendulacea (L.) Pruski Improved the Tolerance of Hybrid to Cadmium Stress. Chemosphere 2020, 249, 126540. [Google Scholar] [CrossRef] [PubMed]
- Hongbiao, W.; Baolong, N.; Mengkui, X.; Lihua, H.; Weifeng, S.; Zhiqi, M. Biological Activities of Cecropin B-Thanatin Hybrid Peptides. J. Pept. Res. 2005, 66, 382–386. [Google Scholar] [CrossRef]
- Boman, H.G.; Nilsson-Faye, I.; Paul, K.; Rasmuson, T. Insect immunity. I. Characteristics of an inducible cell-free antibacterial reaction in hemolymph of Samia cynthia pupae. Infect. Immun. 1974, 10, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Hanson, M.A.; Dostálová, A.; Ceroni, C.; Poidevin, M.; Kondo, S.; Lemaitre, B. Synergy and Remarkable Specificity of Antimicrobial Peptides in Vivo Using a Systematic Knockout Approach. Elife 2019, 8, e44341. [Google Scholar] [CrossRef]
- Vizioli, J.; Richman, A.M.; Uttenweiler-Joseph, S.; Blass, C.; Bulet, P. The Defensin Peptide of the Malaria Vector Mosquito Anopheles Gambiae: Antimicrobial Activities and Expression in Adult Mosquitoes. Insect. Biochem. Mol. Biol. 2001, 31, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Casteels, P.; Ampe, C.; Riviere, L.; Van Damme, J.; Elicone, C.; Fleming, M.; Jacobs, F.; Tempst, P. Isolation and Characterization of Abaecin, a Major Antibacterial Response Peptide in the Honeybee (Apis Mellifera). Eur. J. Biochem. 1990, 187, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, J.; Jiang, L.; Yu, X.; Zhu, H.; Zhang, J.; Feng, Z.; Zhang, X.; Chen, G.; Zhang, Z. Black Soldier Fly (Hermetia Illucens) Larvae Significantly Change the Microbial Community in Chicken Manure. Curr. Microbiol. 2021, 78, 303–315. [Google Scholar] [CrossRef]
- Xia, J.; Ge, C.; Yao, H. Antimicrobial Peptides from Black Soldier Fly (Hermetia Illucens) as Potential Antimicrobial Factors Representing an Alternative to Antibiotics in Livestock Farming. Animals 2021, 11, 1937. [Google Scholar] [CrossRef] [PubMed]
- Almeida, C.; Rijo, P.; Rosado, C. Bioactive Compounds from Hermetia Illucens Larvae as Natural Ingredients for Cosmetic Application. Biomolecules 2020, 10, 976. [Google Scholar] [CrossRef]
- Scala, A.; Cammack, J.A.; Salvia, R.; Scieuzo, C.; Franco, A.; Bufo, S.A.; Tomberlin, J.K.; Falabella, P. Rearing Substrate Impacts Growth and Macronutrient Composition of Hermetia Illucens (L.) (Diptera: Stratiomyidae) Larvae Produced at an Industrial Scale. Sci. Rep. 2020, 10, 19448. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Luo, X.; Fang, G.; Zhan, S.; Wu, J.; Wang, D.; Huang, Y. Transgenic Expression of Antimicrobial Peptides from Black Soldier Fly Enhance Resistance against Entomopathogenic Bacteria in the Silkworm, Bombyx Mori. Insect Biochem. Mol. Biol. 2020, 127, 103487. [Google Scholar] [CrossRef] [PubMed]
- Dean, R.; Van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 Fungal Pathogens in Molecular Plant Pathology: Top 10 Fungal Pathogens. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef]
- Dagdas, Y.F.; Yoshino, K.; Dagdas, G.; Ryder, L.S.; Bielska, E.; Steinberg, G.; Talbot, N.J. Septin-Mediated Plant Cell Invasion by the Rice Blast Fungus, Magnaporthe Oryzae. Science 2012, 336, 1590–1595. [Google Scholar] [CrossRef]
- Eseola, A.B.; Ryder, L.S.; Osés-Ruiz, M.; Findlay, K.; Yan, X.; Cruz-Mireles, N.; Molinari, C.; Garduño-Rosales, M.; Talbot, N.J. Investigating the Cell and Developmental Biology of Plant Infection by the Rice Blast Fungus Magnaporthe Oryzae. Fungal Genet. Biol. 2021, 154, 103562. [Google Scholar] [CrossRef] [PubMed]
- Foster, A.J.; Ryder, L.S.; Kershaw, M.J.; Talbot, N.J. The Role of Glycerol in the Pathogenic Lifestyle of the Rice Blast Fungus Magnaporthe Oryzae. Environ. Microbiol. 2017, 19, 1008–1016. [Google Scholar] [CrossRef]
- Gisi, U.; Sierotzki, H.; Cook, A.; McCaffery, A. Mechanisms Influencing the Evolution of Resistance to Qo Inhibitor Fungicides. Pest. Manag. Sci. 2002, 58, 859–867. [Google Scholar] [CrossRef]
- Canavan, K.; Canavan, S.; Harms, N.E.; Lambertini, C.; Paterson, I.D.; Thum, R. The Potential for Biological Control on Cryptic Plant Invasions. Biological. Control 2020, 144, 104243. [Google Scholar] [CrossRef]
- Mourgues, F.; Brisset, M.N.; Chevreau, E. Strategies to Improve Plant Resistance to Bacterial Diseases through Genetic Engineering. Trends Biotechnol. 1998, 16, 203–210. [Google Scholar] [CrossRef]
- Heath, M.C. Nonhost Resistance and Nonspecific Plant Defenses. Curr. Opin. Plant Biol. 2000, 3, 315–319. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Wang, Y.; Fu, F.; Li, W. Transgenic Improvement for Biotic Resistance of Crops. Int. J. Mol. Sci. 2022, 23, 14370. [Google Scholar] [CrossRef] [PubMed]
- Milanović, V.; Roncolini, A.; Cardinali, F.; Garofalo, C.; Aquilanti, L.; Riolo, P.; Ruschioni, S.; Corsi, L.; Isidoro, N.; Zarantoniello, M.; et al. Occurrence of Antibiotic Resistance Genes in Hermetia Illucens Larvae Fed Coffee Silverskin Enriched with Schizochytrium Limacinum or Isochrysis Galbana Microalgae. Genes 2021, 12, 213. [Google Scholar] [CrossRef] [PubMed]
- Elhag, O.; Zhang, Y.; Xiao, X.; Cai, M.; Zheng, L.; Jordan, H.R.; Tomberlin, J.K.; Huang, F.; Yu, Z.; Zhang, J. Inhibition of Zoonotic Pathogens Naturally Found in Pig Manure by Black Soldier Fly Larvae and Their Intestine Bacteria. Insects 2022, 13, 66. [Google Scholar] [CrossRef]
- Ditsawanon, T.; Roytrakul, S.; Phaonakrop, N.; Charoenlappanit, S.; Thaisakun, S.; Parinthawong, N. Novel Small Antimicrobial Peptides Extracted from Agricultural Wastes Act against Phytopathogens but Not Rhizobacteria. Agronomy 2022, 12, 1841. [Google Scholar] [CrossRef]
- Chen, Y.; Zeng, Q.; Man, Y.; Liu, S.; Ouyang, C.; Li, C.; Wu, X.; Zhang, D.; Liu, Y.; Tan, X. Simple Sequence Repeat Markers Reflect the Biological Phenotype Differentiation and Genetic Diversity of Colletotrichum Gloeosporioides Strains from Capsicum Annuum L. in China. J. Phytopathol. 2021, 169, 701–709. [Google Scholar] [CrossRef]
- Zhu, Z.; Yu, J.; Tang, X.; Xiong, A.; Sun, M. Selection and Validation of Reference Genes in Different Tissues of Okra (Abelmoschus Esculentus L.) under Different Abiotic Stresses. Genes 2023, 14, 603. [Google Scholar] [CrossRef]
- Park, S.-I.; Chang, B.S.; Yoe, S.M. Detection of Antimicrobial Substances from Larvae of the Black Soldier Fly, Hermetia Illucens (Diptera: Stratiomyidae). Entomol. Res. 2014, 44, 58–64. [Google Scholar] [CrossRef]
- Wu, C.-L.; Chih, Y.-H.; Hsieh, H.-Y.; Peng, K.-L.; Lee, Y.-Z.; Yip, B.-S.; Sue, S.-C.; Cheng, J.-W. High Level Expression and Purification of Cecropin-like Antimicrobial Peptides in Escherichia coli. Biomedicines 2022, 10, 1351. [Google Scholar] [CrossRef]
- Chen, Y.; Le, X.; Sun, Y.; Li, M.; Zhang, H.; Tan, X.; Zhang, D.; Liu, Y.; Zhang, Z. MoYcp4 Is Required for Growth, Conidiogenesis and Pathogenicity in Magnaporthe Oryzae. Mol. Plant Pathol. 2017, 18, 1001–1011. [Google Scholar] [CrossRef]
- Zhang, C.; Qi, Z.; Jiao, P.; Jiang, Z.; Liu, S.; Fan, S.; Zhao, C.; Ma, Y.; Guan, S. Cloning and Disease Resistance Analysis of the Maize ZmBON3 Gene. Agronomy 2023, 13, 152. [Google Scholar] [CrossRef]
- Park, S.-I.; Yoe, S.M. A Novel Cecropin-like Peptide from Black Soldier Fly, Hermetia Illucens: Isolation, Structural and Functional Characterization. Entomol. Res. 2017, 47, 115–124. [Google Scholar] [CrossRef]
- De Smet, J.; Wynants, E.; Cos, P.; Van Campenhout, L. Microbial Community Dynamics during Rearing of Black Soldier Fly Larvae (Hermetia Illucens) and Impact on Exploitation Potential. Appl. Environ. Microbiol. 2018, 84, e02722-17. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Vasil, A.I.; Hale, J.D.; Hancock, R.E.W.; Vasil, M.L.; Hodges, R.S. Effects of Net Charge and the Number of Positively Charged Residues on the Biological Activity of Amphipathic α-Helical Cationic Antimicrobial Peptides. Biopolymers 2008, 90, 369–383. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.G.; Hahm, K.-S.; Shin, S.Y. Structure and Fungicidal Activity of a Synthetic Antimicrobial Peptide, P18, and Its Truncated Peptides. Biotechnol. Lett. 2004, 26, 337–341. [Google Scholar] [CrossRef]
- Lee, D.G.; Kim, H.K.; Kim, S.A.; Park, Y.; Park, S.-C.; Jang, S.-H.; Hahm, K.-S. Fungicidal Effect of Indolicidin and Its Interaction with Phospholipid Membranes. Biochem. Biophys. Res. Commun. 2003, 305, 305–310. [Google Scholar] [CrossRef]
- Barbault, F.; Landon, C.; Guenneugues, M.; Meyer, J.-P.; Schott, V.; Dimarcq, J.-L.; Vovelle, F. Solution Structure of Alo-3: A New Knottin-Type Antifungal Peptide from the Insect Acrocinus Longimanus. Biochemistry 2003, 42, 14434–14442. [Google Scholar] [CrossRef]
- Nakajima, Y.; Qu, X.M.; Natori, S. Interaction between Liposomes and Sarcotoxin IA, a Potent Antibacterial Protein of Sarcophaga Peregrina (Flesh Fly). J. Biol. Chem. 1987, 262, 1665–1669. [Google Scholar] [CrossRef]
- Wu, J.; Mu, L.; Zhuang, L.; Han, Y.; Liu, T.; Li, J.; Yang, Y.; Yang, H.; Wei, L. A Cecropin-like Antimicrobial Peptide with Anti-Inflammatory Activity from the Black Fly Salivary Glands. Parasit. Vectors 2015, 8, 561. [Google Scholar] [CrossRef]
- Natori, S. Function of Antimicrobial Proteins in Insects. Ciba Found. Symp. 1994, 186, 123–132, discussion 132–134. [Google Scholar] [CrossRef]
- Nunes, C.; Sucena, É.; Koyama, T. Endocrine Regulation of Immunity in Insects. FEBS J. 2021, 288, 3928–3947. [Google Scholar] [CrossRef] [PubMed]
- Feng, M.; Fei, S.; Xia, J.; Labropoulou, V.; Swevers, L.; Sun, J. Antimicrobial Peptides as Potential Antiviral Factors in Insect Antiviral Immune Response. Front. Immunol. 2020, 11, 2030. [Google Scholar] [CrossRef]
- Hultmark, D. Drosophila Immunity: Paths and Patterns. Curr. Opin. Immunol. 2003, 15, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Bettencourt, R.; Ip, Y.T. Learning the Codes of Fly Immunity. Mol. Cell 2004, 13, 1–2. [Google Scholar] [CrossRef]
- Castillo, J.C.; Reynolds, S.E.; Eleftherianos, I. Insect Immune Responses to Nematode Parasites. Trends Parasitol. 2011, 27, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.-N.; Tong, Y.-C.; Wang, H.-L.; Zhang, J.; Li, Z.-X.; Abbas, Z.; Yang, T.-T.; Liu, M.-Y.; Chen, P.-Y.; Hua, Z.-C.; et al. Novel Hybrid Peptide Cathelicidin 2 (1-13)-Thymopentin (TP5) and Its Derived Peptides with Effective Antibacterial, Antibiofilm, and Anti-Adhesion Activities. Int. J. Mol. Sci. 2021, 22, 11681. [Google Scholar] [CrossRef]
- Hoelscher, M.P.; Forner, J.; Calderone, S.; Krämer, C.; Taylor, Z.; Loiacono, F.V.; Agrawal, S.; Karcher, D.; Moratti, F.; Kroop, X.; et al. Expression Strategies for the Efficient Synthesis of Antimicrobial Peptides in Plastids. Nat. Commun. 2022, 13, 5856. [Google Scholar] [CrossRef]
- Wei, X.; Wu, R.; Zhang, L.; Ahmad, B.; Si, D.; Zhang, R. Expression, Purification, and Characterization of a Novel Hybrid Peptide with Potent Antibacterial Activity. Molecules 2018, 23, 1491. [Google Scholar] [CrossRef]
- Cheng, J.; Ahmat, M.; Guo, H.; Wei, X.; Zhang, L.; Cheng, Q.; Zhang, J.; Wang, J.; Si, D.; Zhang, Y.; et al. Expression, Purification and Characterization of a Novel Hybrid Peptide CLP with Excellent Antibacterial Activity. Molecules 2021, 26, 7142. [Google Scholar] [CrossRef]
- Wu, R.; Wang, Q.; Zheng, Z.; Zhao, L.; Shang, Y.; Wei, X.; Liao, X.; Zhang, R. Design, Characterization and Expression of a Novel Hybrid Peptides Melittin (1-13)-LL37 (17-30). Mol. Biol. Rep. 2014, 41, 4163–4169. [Google Scholar] [CrossRef]
- Qi, L.; Kim, Y.; Jiang, C.; Li, Y.; Peng, Y.; Xu, J.-R. Activation of Mst11 and Feedback Inhibition of Germ Tube Growth in Magnaporthe Oryzae. Mol. Plant Microbe Interact 2015, 28, 881–891. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, X.; Hu, S.; Liu, H.; Xu, J.-R. PKA Activity Is Essential for Relieving the Suppression of Hyphal Growth and Appressorium Formation by MoSfl1 in Magnaporthe Oryzae. PLoS Genet. 2017, 13, e1006954. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.; Liu, Y.; Shui, L.Y.; Zhao, Z.Y.; Mao, X.F.; Liu, Z.Y. Mechanisms of Action of the Antimicrobial Peptide Cecropin in the Killing of Candida albicans. Life 2022, 12, 1581. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.-Q.; Peng, J.; Yang, L.-B.; Jiao, Z.-L.; Zhou, L.-X.; Tao, R.-Y.; Zhu, L.-J.; Tian, Z.-Q.; Huang, M.-J.; Guo, G. A Cecropin-4 Derived Peptide C18 Inhibits Candida Albicans by Disturbing Mitochondrial Function. Front. Microbiol. 2022, 13, 872322. [Google Scholar] [CrossRef] [PubMed]
- Ko, E.J.; Ho Shin, Y.; Hyun, H.N.; Song, H.S.; Hong, J.K.; Jeun, Y.C. Bio-Sulfur Pre-Treatment Suppresses Anthracnose on Cucumber Leaves Inoculated with Colletotrichum Orbiculare. Mycobiology 2019, 47, 308–318. [Google Scholar] [CrossRef]
- Lee, Y.; Ko, Y.; Jeun, Y. Illustration of Disease Suppression of Anthracnose on Cucumber Leaves by Treatment with Chlorella Fusca. Res. Plant Dis. 2016, 22, 257–263. [Google Scholar] [CrossRef]
- Chumley, F.; Valent, B. Genetic Analysis of Melanin-Deficient, Nonpathogenic Mutants of Magnaporthe Grisea. Mol. Plant-Microbe Interact 1990, 3, 135–143. [Google Scholar] [CrossRef]
- Fan, Y.; Miguez-Macho, G.; Jobbágy, E.G.; Jackson, R.B.; Otero-Casal, C. Hydrologic Regulation of Plant Rooting Depth. Proc. Natl. Acad. Sci. USA 2017, 114, 10572–10577. [Google Scholar] [CrossRef]
- Zheng, M.; Tian, S.Z.; Capurso, D.; Kim, M.; Maurya, R.; Lee, B.; Piecuch, E.; Gong, L.; Zhu, J.J.; Li, Z.; et al. Multiplex Chromatin Interactions with Single-Molecule Precision. Nature 2019, 566, 558–562. [Google Scholar] [CrossRef]
- Saugar, J.M.; Rodríguez-Hernández, M.J.; de la Torre, B.G.; Pachón-Ibañez, M.E.; Fernández-Reyes, M.; Andreu, D.; Pachón, J.; Rivas, L. Activity of Cecropin A-Melittin Hybrid Peptides against Colistin-Resistant Clinical Strains of Acinetobacter Baumannii: Molecular Basis for the Differential Mechanisms of Action. Antimicrob. Agents Chemother. 2006, 50, 1251–1256. [Google Scholar] [CrossRef]
- Tan, T.; Wu, D.; Li, W.; Zheng, X.; Li, W.; Shan, A. High Specific Selectivity and Membrane-Active Mechanism of Synthetic Cationic Hybrid Antimicrobial Peptides Based on the Peptide FV7. Int. J. Mol. Sci. 2017, 18, 339. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Lin, Q.; Li, B.; Li, N.; Zhang, S. Expression and Purification the Antimicrobial Peptide CM4 in Escherichia coli. Biotechnol. Lett. 2009, 31, 437–441. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.Y.; Kang, J.H.; Hahm, K.S. Structure-Antibacterial, Antitumor and Hemolytic Activity Relationships of Cecropin A-Magainin 2 and Cecropin A-Melittin Hybrid Peptides. J. Pept. Res. 1999, 53, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Wiradharma, N.; Khoe, U.; Hauser, C.A.E.; Seow, S.V.; Zhang, S.; Yang, Y.-Y. Synthetic Cationic Amphiphilic α-Helical Peptides as Antimicrobial Agents. Biomaterials 2011, 32, 2204–2212. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | C. acutatum (HHDL02) MIC50 (μM) | M. oryzae (Guy11) MIC50 (μM) |
|---|---|---|
| CAD1 | 51 | 40 |
| CAD5 | 49 | 43 |
| CAD7 | 44 | 43 |
| CAD-Con | 22 | 15 |
| Strain | C. acutatum (HHDL02) Germ Tube (μm) | M. oryzae (Guy11) Germ Tube (μm) | ||
|---|---|---|---|---|
| 2 h | 4 h | 4 h | 8 h | |
| CK | 6.49 ± 0.50 a | 13.50 ± 1.10 a | 41.82 ± 8.37 a | 74.12 ± 9.44 a |
| CAD1 | 293.96 ± 11.40 b | 808.30 ± 18.28 b | 600.13 ± 16.25 b | 1309.88 ± 23.23 b |
| ACD5 | 294.81 ± 13.61 b | 815.70 ± 16.85 b | 633.77 ± 18.57 b | 1343.67 ± 15.84 b |
| ACD7 | 304.30 ± 14.90 b | 799.26 ± 25.03 b | 609.12 ± 16.56 b | 1322.07 ± 18.61 b |
| CAD-Con | 367.26 ± 19.74 b | 986.74 ± 23.73 b | 774.09 ± 22.28 b | 1613.34 ± 36.75 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Q.; Zhang, X.; Ouyang, Y.; Yu, P.; Man, Y.; Guo, S.; Liu, S.; Chen, Y.; Wang, Y.; Tan, X. Appressoria Formation in Phytopathogenic Fungi Suppressed by Antimicrobial Peptides and Hybrid Peptides from Black Soldier Flies. Genes 2023, 14, 1096. https://doi.org/10.3390/genes14051096
Sun Q, Zhang X, Ouyang Y, Yu P, Man Y, Guo S, Liu S, Chen Y, Wang Y, Tan X. Appressoria Formation in Phytopathogenic Fungi Suppressed by Antimicrobial Peptides and Hybrid Peptides from Black Soldier Flies. Genes. 2023; 14(5):1096. https://doi.org/10.3390/genes14051096
Chicago/Turabian StyleSun, Qianlong, Xin Zhang, Ying Ouyang, Pingzhong Yu, Yilong Man, Sheng Guo, Sizhen Liu, Yue Chen, Yunsheng Wang, and Xinqiu Tan. 2023. "Appressoria Formation in Phytopathogenic Fungi Suppressed by Antimicrobial Peptides and Hybrid Peptides from Black Soldier Flies" Genes 14, no. 5: 1096. https://doi.org/10.3390/genes14051096
APA StyleSun, Q., Zhang, X., Ouyang, Y., Yu, P., Man, Y., Guo, S., Liu, S., Chen, Y., Wang, Y., & Tan, X. (2023). Appressoria Formation in Phytopathogenic Fungi Suppressed by Antimicrobial Peptides and Hybrid Peptides from Black Soldier Flies. Genes, 14(5), 1096. https://doi.org/10.3390/genes14051096