
Plant Promoters: Their Identification, Characterization, and Role in Gene Regulation

 , ,
, ,
Abstract
:1. Introduction
1.1. Promoters and Their Importance in Genetically Modified Crops
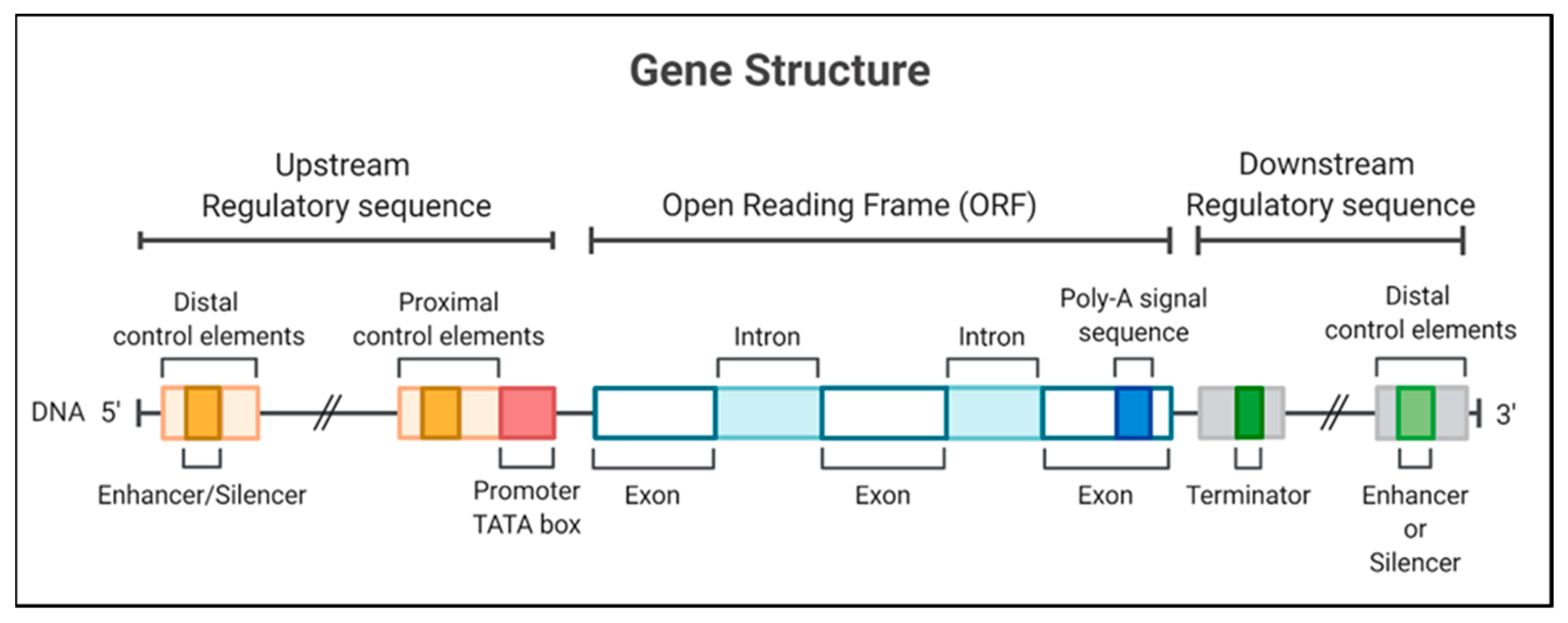
1.2. Types of Promoters
1.3. Constitutive Promoters
1.4. Tissue-Specific Promoters
1.5. Inducible Promoters
1.6. Synthetic Promoters
1.7. Use of Reporter Genes in Plant Promoter Characterization
2. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Potenza, C.; Aleman, L.; Sengupta-Gopalan, C. Targeting transgene expression in research, agricultural, and en-vironmental applications: Promoters used in plant transformation. Vitr. Cell. Dev. Biol. Plant 2004, 40, 1–22. [Google Scholar] [CrossRef]
- Danila, F.; Schreiber, T.; Ermakova, M.; Hua, L.; Vlad, D.; Lo, S.; Chen, Y.; Lambret-Frotte, J.; Hermanns, A.S.; Athmer, B.; et al. A single promoter-TALE system for tissue-specific and tuneable expression of multiple genes in rice. Plant Biotechnol. J. 2022, 20, 1786–1806. [Google Scholar] [CrossRef] [PubMed]
- Dafny-Yelin, M.; Tzfira, T. Delivery of Multiple Transgenes to Plant Cells. Plant Physiol. 2007, 145, 1118–1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Southern, M.M.; Brown, P.E.; Hall, A. Luciferases as Reporter Genes. Methods Mol. Biol. 2006, 323, 293–306. [Google Scholar] [CrossRef] [PubMed]
- Kor, S.D.; Chowdhury, N.; Keot, A.K.; Yogendra, K.; Chikkaputtaiah, C.; Reddy, P.S. RNA Pol III promoters—Key players in precisely targeted plant genome editing. Front. Genet. 2023, 13, 989199. [Google Scholar] [CrossRef] [PubMed]
- Dutt, M.; Dhekney, S.A.; Soriano, L.; Kandel, R.; Grosser, J.W. Temporal and spatial control of gene expression in horticultural crops. Hortic. Res. 2014, 1, 14047. [Google Scholar] [CrossRef] [Green Version]
- Cai, M.; Wei, J.; Li, X.; Xu, C.; Wang, S. A rice promoter containing both novel positive and negative cis-elements for regulation of green tissue-specific gene expression in transgenic plants. Plant Biotechnol. J. 2007, 5, 664–674. [Google Scholar] [CrossRef]
- Porto, M.S.; Pinheiro, M.P.N.; Batista, V.G.L.; Dos Santos, R.C.; De Albuquerque Melo Filho, P.; De Lima, L.M. Plant promoters: An approach of structure and function. Mol. Biotechnol. 2014, 56, 38–49. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Garcia, C.M.; Finer, J.J. Identification and validation of promoters and cis-acting regulatory elements. Plant Sci. 2014, 217–218, 109–119. [Google Scholar] [CrossRef] [Green Version]
- Damaj, M.B.; Beremand, P.D.; Buenrostro-Nava, M.T.; Ivy, J.; Kumpatla, S.P.; Jifon, J.; Beyene, G.; Yu, Q.; Thomas, T.L.; Mirkov, T.E. Isolating promoters of multigene family members from the polyploid sugarcane genome by PCR-based walking in BAC DNA. Genome 2010, 53, 840–847. [Google Scholar] [CrossRef]
- Devic, M.; Albert, S.; Delseny, M.; Roscoe, T.J. Efficient PCR walking on plant genomic DNA. Plant Physiol. Biochem. 1997, 35, 331–339. [Google Scholar]
- Rishi, A.S.; Nelson, N.D.; Goyal, A. Genome walking of large fragments: An improved method. J. Biotechnol. 2004, 111, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Santos, E.; Pacheco, R.; Villao, L.; Galarza, L.; Ochoa, D.; Jordán, C.; Flores, J. Promoter analysis in banana. In Banana: Genomics and Transgenic Approaches for Genetic Improvement; Springer: Singapore, 2016; pp. 157–179. [Google Scholar] [CrossRef]
- Shahmuradov, I.A.; Gammerman, A.J.; Hancock, J.M.; Bramley, P.M.; Solovyev, V.V. PlantProm: A database of plant promoter sequences. Nucleic Acids Res. 2003, 31, 114–117. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Shujaat, M.; Tayara, H. iProm-Zea: A two-layer model to identify plant promoters and their types using convolutional neural network. Genomics 2022, 114, 110384. [Google Scholar] [CrossRef] [PubMed]
- Shahmuradov, I.A.; Umarov, R.; Solovyev, V.V. TSSPlant: A new tool for prediction of plant Pol II promoters. Nucleic Acids Res. 2017, 45, e65. [Google Scholar] [CrossRef] [PubMed]
- Zheng, N.; Li, T.; Dittman, J.D.; Su, J.; Li, R.; Gassmann, W.; Peng, D.; Whitham, S.A.; Liu, S.; Yang, B. CRISPR/Cas9-Based Gene Editing Using Egg Cell-Specific Promoters in Arabidopsis and Soybean. Front. Plant Sci. 2020, 11, 800. [Google Scholar] [CrossRef]
- Jiang, F.; Doudna, J.A. CRISPR–Cas9 Structures and Mechanisms. Annu. Rev. Biophys. 2017, 46, 505–529. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Wang, J.; Ye, M.X.; Li, H.; Ji, L.X.; Li, Y.; Cui, D.Q.; Liu, J.M.; An, X.M. A Novel Moderate Constitutive Promoter Derived from Poplar (Populus tomentosa Carrière). Int. J. Mol. Sci. 2013, 14, 6187. [Google Scholar] [CrossRef] [Green Version]
- McElroy, D.; Zhang, W.; Cao, J.; Wu, R. Isolation of an Efficient Actin Promoter for Use in Rice Transformation. Plant Cell 1990, 2, 163. [Google Scholar]
- Wang, Y.; Zhang, W.; Cao, J.; McElroy, D.; Wu, R. Characterization of cis-acting elements regulating transcription from the promoter of a constitutively active rice actin gene. Mol. Cell Biol. 1992, 12, 3399–3406. [Google Scholar]
- Kyozuka, J.; Fujimoto, H.; Izawa, T.; Shimamoto, K. Anaerobic induction and tissue-specific expression of maize Adh1 promoter in transgenic rice plants and their progeny. Mol. Genet. Genom. 1991, 228, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Onnela, M.-L.; Suihko, M.-L.; Penttilä, M.; Keränen, S. Use of a modified alcohol dehydrogenase, ADH1, promoter in construction of diacetyl non-producing brewer’s yeast. J. Biotechnol. 1996, 49, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Naito, S.; Komeda, Y. The Arabidopsis HSP18.2 promoter/GUS gene fusion in transgenic Arabidopsis plants: A powerful tool for the isolation of regulatory mutants of the heat-shock response. Plant J. 1992, 2, 751–761. [Google Scholar] [CrossRef]
- Tzafrir, I.; Torbert, K.A.; Lockhart, B.E.; Somers, D.A.; Olszewski, N.E. The sugarcane bacilliform badnavirus promoter is active in both monocots and dicots. Plant Mol. Biol. 1998, 38, 347–356. [Google Scholar] [CrossRef]
- Schenk, P.M.; Sagi, L.; Remans, T.; Dietzgen, R.G.; Bernard, M.J.; Graham, M.W.; Manners, J.M. A promoter from sugarcane bacilliform badnavirus drives transgene expression in banana and other monocot and dicot plants. Plant Mol. Biol. 1999, 39, 1221–1230. [Google Scholar] [CrossRef]
- Christensen, A.H.; Sharrock, R.A.; Quail, P.H. Maize polyubiquitin genes: Structure, thermal perturbation of expression and transcript splicing, and promoter activity following transfer to protoplasts by electroporation. Plant Mol. Biol. 1992, 18, 675–689. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Oard, J.H. Rice ubiquitin promoters: Deletion analysis and potential usefulness in plant transformation systems. Plant Cell Rep. 2003, 22, 129–134. [Google Scholar] [CrossRef]
- Hernandez-Garcia, C.M.; Martinelli, A.P.; Bouchard, R.A.; Finer, J.J. A soybean (Glycine max) polyubiquitin promoter gives strong constitutive expression in transgenic soybean. Plant Cell Rep. 2009, 28, 837–849. [Google Scholar] [CrossRef]
- Odell, J.T.; Nagy, F.; Chua, N.-H. Identification of DNA sequences required for activity of the cauliflower mosaic virus 35S promoter. Nature 1985, 313, 810–812. [Google Scholar] [CrossRef]
- An, G.; Costa, M.A.; Ha, S.B. Nopaline synthase promoter is wound inducible and auxin inducible. Plant Cell 1990, 2, 225–233. [Google Scholar] [CrossRef] [Green Version]
- Bevan, M.; Barnes, W.M.; Chilton, M.-D. Strcuture and transcription of the nopaline synthase gene region of T-DNA. Nucleic Acids Res. 1983, 11, 369–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahoo, D.K.; Sarkar, S.; Maiti, I.B.; Dey, N. Novel Synthetic Promoters from the Cestrum Yellow Leaf Curling Virus. Methods Mol. Biol. 2016, 1482, 111–138. [Google Scholar] [PubMed]
- Kelly, G.; Lugassi, N.; Belausov, E.; Wolf, D.; Khamaisi, B.; Brandsma, D.; Kottapalli, J.; Fidel, L.; Ben-Zvi, B.; Egbaria, A.; et al. The Solanum tuberosum KST1 partial promoter as a tool for guard cell expression in multiple plant species. J. Exp. Bot. 2017, 68, 2885–2897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, S.; Ding, W.; Bai, W.; Lu, J.; Zhou, L.; Ma, X.; Zhu, Q. Application of a novel strong promoter from Chinese fir (Cunninghamia lanceolate) in the CRISPR/Cas mediated genome editing of its protoplasts and transgenesis of rice and poplar. Front Plant Sci. 2023, 14, 1289. [Google Scholar] [CrossRef]
- Li, J.; Xu, R.F.; Qin, R.Y.; Ma, H.; Li, H.; Zhang, Y.P.; Li, L.; Wei, P.C.; Yang, J.B. Isolation and functional characterization of a novel rice constitutive promoter. Plant Cell Rep. 2014, 33, 1651–1660. [Google Scholar] [CrossRef] [PubMed]
- Masura, S.S.; Parveez, G.K.A.; Ti, L.L.E. Isolation and characterization of an oil palm constitutive promoter derived from a translationally control tumor protein (TCTP) gene. Plant Physiol. Biochem. 2011, 49, 701–708. [Google Scholar] [CrossRef]
- Santos, E.; Remy, S.; Thiry, E.; Windelinckx, S.; Swennen, R.; Sági, L. Characterization and isolation of a T-DNA tagged banana promoter active during in vitro culture and low temperature stress. BMC Plant Biol. 2009, 9, 100187. [Google Scholar] [CrossRef] [Green Version]
- Villao, L.; Sánchez, E.; Romero, C.; Galarza, L.; Flores, J.; Santos-Ordóñez, E. Activity characterization of the plantain promoter from the heavy metal-associated isoprenylated plant gene (MabHIPP) using the luciferase reporter gene. Plant Gene 2019, 19, 100187. [Google Scholar] [CrossRef]
- Villao, L.; Flores, J.; Santos-Ordóñez, E. Genetic transformation of apical meristematic shoots in the banana cultivar ‘Williams’. Bionatura 2021, 6, 1462–1465. [Google Scholar] [CrossRef]
- Park, S.H.; Yi, N.; Kim, Y.S.; Jeong, M.H.; Bang, S.W.; Choi YDo Kim, J.K. Analysis of five novel putative constitutive gene promoters in transgenic rice plants. J. Exp. Bot. 2010, 61, 2459. [Google Scholar] [CrossRef] [Green Version]
- Cornejo, M.J.; Luth, D.; Blankenship, K.M.; Anderson, O.D.; Blechl, A.E. Activity of a maize ubiquitin promoter in transgenic rice. Plant Mol. Biol. 1993, 23, 567–581. [Google Scholar] [CrossRef] [PubMed]
- Marín-Rodríguez, M.C.; Smith, D.L.; Manning, K.; Orchard, J.; Seymour, G.B. Pectate lyase gene expression and enzyme activity in ripening banana fruit. Plant Mol. Biol. 2003, 51, 851–857. [Google Scholar] [CrossRef] [PubMed]
- Nazir, S.; Iqbal, M.Z.; Rahman, S.U. Molecular Identification of Genetically Modified Crops for Biosafety and Legitimacy of Transgenes. In Gene Editing-Technologies and Applications; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef] [Green Version]
- Manavella, P.A.; Chan, R.L. Development of Tissue-Specific Promoters for Plant Transformation. Plant Physiol. 2006, 2, 21–25. [Google Scholar]
- Paul, J.Y.; Khanna, H.; Kleidon, J.; Hoang, P.; Geijskes, J.; Daniells, J.; Zaplin, E.; Rosenberg, Y.; James, A.; Mlalazi, B.; et al. Golden bananas in the field: Elevated fruit pro-vitamin A from the expression of a single banana transgene. Plant Biotechnol. J. 2017, 15, 520–532. [Google Scholar] [CrossRef] [Green Version]
- Ye, R.; Zhou, F.; Lin, Y. Two novel positive cis-regulatory elements involved in green tissue-specific promoter activity in rice (Oryza sativa L. ssp.). Plant Cell Rep. 2012, 31, 1159–1172. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, P.K.; Nath, P. MaExp1, an ethylene-induced expansin from ripening banana fruit. Plant Sci. 2004, 167, 1351–1358. [Google Scholar] [CrossRef]
- Sane, V.A.; Sane, A.P.; Nath, P. Multiple forms of α-expansin genes are expressed during banana fruit ripening and development. Postharvest Biol. Technol. 2007, 45, 184–192. [Google Scholar]
- Singh, K.B.; Foley, R.C.; Oñate-Sánchez, L. Transcription factors in plant defense and stress responses. Curr. Opin. Plant Biol. 2002, 5, 430–436. [Google Scholar] [CrossRef]
- Mazarei, M.; Al-Ahmad, H.; Rudis, M.R.; Stewart, C.N., Jr. Protoplast isolation and transient gene expression in switchgrass, Panicum virgatum L. Biotechnol. J. 2008, 3, 354–359. [Google Scholar] [CrossRef]
- Gurr, S.J.; Rushton, P.J. Engineering plants with increased disease resistance: What are we going to express? Trends Biotechnol. 2005, 23, 275–282. [Google Scholar] [CrossRef]
- Bustos, M.M.; Kalkan, F.A.; VandenBosch, K.A.; Hall, T.C. Differential accumulation of four phaseolin glycoforms in transgenic tobacco. Plant Mol. Biol. 1991, 16, 381–395. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekharan, M.B.; Bishop, K.J.; Hall, T.C. Module-specific regulation of the β-phaseolin promoter during embryogenesis. Plant J. 2003, 33, 853–866. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Dahal, P.; Bradford, K.J. Two Tomato Expansin Genes Show Divergent Expression and Localization in Embryos during Seed Development and Germination. Plant Physiol. 2001, 127, 928–936. [Google Scholar] [CrossRef] [PubMed]
- Zubaidah, R.; Nurniwalis, A.W.; Siti Nor, A.A.; Ghulam Kadir, A.P. The use of Arabidopsis thaliana model system for testing oil palm promoter: Case study on oil palm mt3-a promoter. J. Oil Palm Res. 2017, 29, 189–196. [Google Scholar] [CrossRef] [Green Version]
- Chan, P.L.; Akmar, A.S.N.; Roohaida, O. Light-harvesting chlorophyll A/B binding protein (LHCB) promoter for targeting specific expression in oil palm leaves. J. Oil Palm. Res. 2008, 2, 21–29. [Google Scholar]
- Mohan, C.; Jayanarayanan, A.N.; Narayanan, S. Construction of a novel synthetic root-specific promoter and its characterization in transgenic tobacco plants. 3 Biotech 2017, 7, 234. [Google Scholar] [CrossRef]
- Li, M.; Song, B.; Zhang, Q.; Liu, X.; Lin, Y.; Ou, Y.; Zhang, H.; Liu, J. A synthetic tuber-specific and cold-induced promoter is applicable in controlling potato cold-induced sweetening. Plant Physiol. Biochem. 2013, 67, 41–47. [Google Scholar] [CrossRef]
- Hiwasa-Tanase, K.; Kuroda, H.; Hirai, T.; Aoki, K.; Takane, K.; Ezura, H. Novel promoters that induce specific transgene expression during the green to ripening stages of tomato fruit development. Plant Cell Rep. 2012, 31, 1415–1424. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.-Y.; Kwon, S.-Y.; Lee, H.-S.; Hur, Y.; Bang, J.-W.; Kwak, S.-S. A novel oxidative stress-inducible peroxidase promoter from sweetpotato: Molecular cloning and characterization in transgenic tobacco plants and cultured cells. Plant Mol. Biol. 2003, 51, 831–838. [Google Scholar] [CrossRef]
- Sasaki, K.; Yuichi, O.; Hiraga, S.; Gotoh, Y.; Seo, S.; Mitsuhara, I.; Ito, H.; Matsui, H.; Ohashi, Y. Characterization of two rice peroxidase promoters that respond to blast fungus-infection. Mol. Genet. Genom. 2007, 278, 709–722. [Google Scholar] [CrossRef]
- Uknes, S.; Dincher, S.; Friedrich, L.; Negrotto, D.; Williams, S.; Thompson-Taylor, H.; Potter, S.; Ward, E.; Ryals, J. Regulation of pathogenesis-related protein-1a gene expression in tobacco. Plant Cell 1993, 5, 159–169. [Google Scholar] [CrossRef]
- Malnoy, M.; Reynoird, J.P.; Borejsza-Wysocka, E.E.; Aldwinckle, H.S. Activation of the Pathogen-Inducible Gst1 Promoter of Potato after Elicitation by Venturia inaequalis and Erwinia amylovora in Transgenic Apple (Malus × Domestica). Transgenic Res. 2006, 15, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Malnoy, M.; Venisse, J.-S.; Reynoird, J.; Chevreau, E. Activation of three pathogen-inducible promoters of tobacco in transgenic pear (Pyrus communis L.) after abiotic and biotic elicitation. Planta 2003, 216, 802–814. [Google Scholar] [CrossRef] [PubMed]
- Jin, B.; Sheng, Z.; Muhammad, I.; Chen, J.; Yang, H. Cloning and functional analysis of the promoter of a stress-inducible gene (Zmap) in maize. PLoS ONE 2019, 14, e0211941. [Google Scholar] [CrossRef] [PubMed]
- Peremarti, A.; Twyman, R.M.; Gómez-Galera, S.; Naqvi, S.; Farré, G.; Sabalza, M.; Miralpeix, B.; Dashevskaya, S.; Yuan, D.; Ramessar, K.; et al. Promoter diversity in multigene transformation. Plant Mol. Biol. 2010, 73, 363–378. [Google Scholar] [CrossRef] [PubMed]
- Jameel, A.; Noman, M.; Liu, W.; Ahmad, N.; Wang, F.; Li, X.; Li, H. Tinkering Cis Motifs Jigsaw Puzzle Led to Root-Specific Drought-Inducible Novel Synthetic Promoters. Int. J. Mol. Sci. 2020, 21, 1357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Does, D.; Leon-Reyes, A.; Koornneef, A.; Van Verk, M.C.; Rodenburg, N.; Pauwels, L.; Goossens, A.; Körbes, A.P.; Memelink, J.; Ritsema, T.; et al. Salicylic Acid Suppresses Jasmonic Acid Signaling Downstream of SCFCOI1-JAZ by Targeting GCC Promoter Motifs via Transcription Factor ORA59. Plant Cell 2013, 25, 744–761. [Google Scholar] [CrossRef] [Green Version]
- Barry, C.S.; Blume, B.; Bouzayen, M.; Cooper, W.; Hamilton, A.J.; Grierson, D. Differential expression of the 1-aminocyclopropane-1-carboxylate oxidase gene family of tomato. Plant J. 1996, 9, 525–535. [Google Scholar] [CrossRef]
- Santamaria, M.; Thomson, C.J.; Read, N.D.; Loake, G.J. The promoter of a basic PR1-like gene, AtPRB1, from Ara-bidopsis establishes an organ-specific expression pattern and responsiveness to ethylene and methyl jasmonate. Plant Mol. Biol. 2001, 47, 641–652. [Google Scholar] [CrossRef]
- Brandalise, M.; Severino, F.E.; Maluf, M.P.; Maia, I.G. The promoter of a gene encoding an isoflavone reductase-like protein in coffee (Coffea arabica) drives a stress-responsive expression in leaves. Plant Cell Rep. 2009, 28, 1699–1708. [Google Scholar] [CrossRef]
- Liu, W.; Stewart, C.N. Plant synthetic promoters and transcription factors. Curr. Opin. Biotechnol. 2016, 37, 36–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rushton, P.; Rushton, P.J.; Reinstädler, A.; Lipka, V.; Lippok, B.; Somssich, I.E. Synthetic plant promoters containing defined regulatory elements provide novel insights into pathogen- and wound-induced signaling Synthetic Plant Promoters Containing Defined Regulatory Elements Provide Novel Insights into Pathogen- and Wound-Induced signaling. The Plant Cell 2002, 14, 749–762. [Google Scholar]
- Khan, A.; Nasim, N.; Pudhuvai, B.; Koul, B.; Upadhyay, S.K.; Sethi, L.; Dey, N. Plant Synthetic Promoters: Advancement and Prospective. Agriculture 2023, 13, 298. [Google Scholar] [CrossRef]
- Das, S.; Bansal, M. Variation of gene expression in plants is influenced by gene architecture and structural properties of promoters. PLoS ONE 2019, 14, e0212678. [Google Scholar] [CrossRef]
- Peremarti, A.; Bassie, L.; Yuan, D.; Pelacho, A.; Christou, P.; Capell, T. Transcriptional regulation of the rice arginine decarboxylase (Adc1) and S-adenosylmethionine decarboxylase (Samdc) genes by methyl jasmonate. Plant Physi-Ology Biochem. 2010, 48, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Phillips-Cremins, J.E.; Corces, V.G. Chromatin Insulators: Linking Genome Organization to Cellular Function. Mol. Cell 2013, 50, 461–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Li, S.; Yang, W.; Mu, B.; Jiao, Y.; Zhou, X.; Zhang, C.; Fan, Y.; Chen, R. Synthesis of Seed-Specific Bidirectional Promoters for Metabolic Engineering of Anthocyanin-Rich Maize. Plant Cell Physiol. 2018, 59, 1942–1955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venter, M.; Botha, F.C. Synthetic promoter engineering. Plant Dev. Biol. 2010, 2, 393–414. [Google Scholar]
- Ren, Q.; Zhong, Z.; Wang, Y.; You, Q.; Li, Q.; Yuan, M.; He, Y.; Qi, C.; Tang, X.; Zheng, X.; et al. Bidirectional Promoter-Based CRISPR-Cas9 Systems for Plant Genome Editing. Front. Plant Sci. 2019, 10, 1173. [Google Scholar] [CrossRef]
- Dash, A.; Gurdaswani, V.; D’souza, J.S.; Ghag, S.B. Functional characterization of an inducible bidirectional promoter from Fusarium oxysporum f. sp. cubense. Sci. Rep. 2020, 10, 2323. [Google Scholar] [CrossRef] [Green Version]
- Jefferson, R.A.; Kavanagh, T.A.; Bevan, M.W. GUS fusions: Beta-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 1987, 6, 3901–3907. [Google Scholar] [CrossRef] [PubMed]
- Heim, R.; Cubitt, A.B.; Tsien, R.Y. Improved green fluorescence. Nature 1995, 373, 663–664. [Google Scholar] [CrossRef] [PubMed]
- Chiu, W.-L.; Niwa, Y.; Zeng, W.; Hirano, T.; Kobayashi, H.; Sheen, J. Engineered GFP as a vital reporter in plants. Curr. Biol. 1996, 6, 325–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ow, D.W.; Wood, K.V.; DeLuca, M.; de Wet, J.R.; Helinski, D.R.; Howell, S.H. Transient and Stable Expression of the Firefly Luciferase Gene in Plant Cells and Transgenic Plants. Science 1986, 234, 856–859. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Golden, S.S.; Kondo, T.; Ishiura, M.; Johnson, C.H. Bacterial luciferase as a reporter of circadian gene expression in cyanobacteria. J. Bacteriol. 1995, 177, 2080–2086. [Google Scholar] [CrossRef] [Green Version]
- Hraška, M.; Rakouský, S.; Čurn, V. Green fluorescent protein as a vital marker for non-destructive detection of transformation events in transgenic plants. Plant Cell Tissue Organ Cult. 2006, 86, 303–318. [Google Scholar] [CrossRef]
- Elliott, A.R.; Campbell, J.A.; Dugdale, B.; Brettell, R.I.S.; Grof, C.P.L. Green-fluorescent protein facilitates rapid in vivo detection of genetically transformed plant cells. Plant Cell Rep. 1999, 18, 707–714. [Google Scholar] [CrossRef]
- Maximova, S.N.; Dandekar, A.M.; Guiltinan, M.J. Investigation of Agrobacterium-mediated transformation of apple using green fluorescent protein: High transient expression and low stable transformation suggest that factors other than T-DNA transfer are rate-limiting. Plant Mol. Biol. 1998, 37, 549–559. [Google Scholar] [CrossRef]
- Verkhusha, V.V.; Shavlovsky, M.M.; Nevzglyadova, O.V.; Gaivoronsky, A.A.; Artemov, A.V.; Stepanenko, O.V.; Kuznetsova, I.M.; Turoverov, K.K. Expression of recombinant GFP-actin fusion protein in the methylotrophic yeast. FEMS Yeast Res. 2003, 3, 105–111. [Google Scholar] [CrossRef]
- Van Leeuwen, W.; Hagendoorn, M.J.M.; Ruttink, T.; Van Poecke, R.; Van Der Plas, L.H.W.; Van Der Krol, A.R. The use of the luciferase reporter system forin planta gene expression studies. Plant Mol. Biol. Rep. 2000, 18, 143–144. [Google Scholar] [CrossRef]
- Remy, S.; Weerdt, G.; Deconinck, I.; Swennen, R.; Sagi, L.; Jain, S. An Ultrasensitive Luminescent Detection System in Banana Biotechnology: From Promoter Tagging to Southern Hybridization; Science Publishers, Inc.: Enfield, NH, USA, 2004; pp. 307–319. [Google Scholar]
- Ali, S.; Kim, W.C. A Fruitful Decade Using Synthetic Promoters in the Improvement of Transgenic Plants. Front Plant Sci. 2019, 10, 1433. [Google Scholar] [CrossRef] [PubMed]
- Kummari, D.; Palakolanu, S.R.; Kishor, P.B.K.; Bhatnagar-Mathur, P.; Singam, P.; Vadez, V.; Sharma, K.K. An update and perspectives on the use of promoters in plant genetic engineering. J. Biosci. 2020, 45, 119. [Google Scholar] [CrossRef] [PubMed]
- Amack, S.C.; Antunes, M.S. CaMV35S promoter—A plant biology and biotechnology workhorse in the era of synthetic biology. Curr. Plant Biol. 2020, 24, 100179. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Promoter | Source | Expression | References |
|---|---|---|---|
| Act1 | Actin gene, rice | The whole plant, preferably monocots | [20,21] |
| Adh1 | Alcohol dehydrogenase gene, maize | Roots, meristematic tissue, endosperm, and pollen (anaerobic regulation) preference in monocots | [22,23] |
| HSP18.2 | Arabidopsis thaliana | Leaves, vascular system | [24] |
| ScBV | Bacilliform virus, sugarcane | Leaves, vascular system, monocots, and dicots | [25,26] |
| Ubi-1 | Ubiquitin gene, maize | Protoplast, monocots. | [27] |
| RUBQ1/RUBQ2 | Ubiquitin gene, rice | Genes expression, monocots. | [28] |
| Gmubi | Soybean | The whole plant | [29] |
| CaMV35S | Cauliflower mosaic virus | Expression throughout the plant, monocots, and dicots | [30] |
| nos | Nopaline synthase gene, Agrobacterium tumefaciens | Expression throughout the plant, monocots, and dicots | [31,32] |
| CmYLCV | Cestrum Yellow Leaf Curling Virus (CmYLCV) | Growth and development | [33] |
| KST1 | Solanum tuberosum (potato) | Guard cell promoter | [34] |
| Cula11/Cula08 | Cunninghamia lanceolate (Chinese fir) | Protoplast, monocots, and dicots | [35] |
| P OsCon1 | Rice | The whole plant, monocots, and dicots | [36] |
| TCTP | Oil palm | Immature embryo, embryogenic callus, embryoid, a young leaflet from a mature palm, green leaf, mesocarp, and stem | [37] |
| Type of Promoter | Promoter | Source | Expression | References |
|---|---|---|---|---|
| Tissue specific | β-phaseolin | Phaseolus vulgaris (phas) | Flowers, seeds, embryogenesis | [53,54] |
| EXP1 | Banana | Ripening fruit | [46,48,49,55] | |
| GSSP1, GSSP3, GSSP5, GSSP6, GSSP7 | Oryza sativa | Bidirectional green tissue | [47] | |
| MT3-A | Oil palm | Mesocarp | [56] | |
| LC01 | Oil palm | Leaf specific | [57] | |
| SynR2 SynR1 | N. tabacum | Root | [58] | |
| Inducible | pCL | S. tuberosum | Gene regulation of the activity of acid vacuolar invertase in potato tubers at low temperature | [59] |
| LA22CD07, LesAffx.6852.1.S1_at | Tomato ripening-induced genes | Chemical factors induce fruit ripening | [60] | |
| POD, POX | Oxidative stress-inducible peroxidase | Rice peroxidase inhibitor, biotic stress | [61,62] | |
| PR-1a | A. thaliana | Related to pathogenesis, the acquired resistance system (SAR) | [63] | |
| GST1 | Potato | Biotic stress | [64] | |
| SGD24-STR246C | Tobacco | Biotic stress | [65] | |
| Zmap | Maize | Different stressors | [66] | |
| CMPG1 | A. thaliana | Pathogen inducible | [67] | |
| Synp16 | Soybean | Abiotic stress | [68] | |
| GCC | A. thaliana | Jasmonic acid inducible | [69] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Villao-Uzho, L.; Chávez-Navarrete, T.; Pacheco-Coello, R.; Sánchez-Timm, E.; Santos-Ordóñez, E. Plant Promoters: Their Identification, Characterization, and Role in Gene Regulation. Genes 2023, 14, 1226. https://doi.org/10.3390/genes14061226
Villao-Uzho L, Chávez-Navarrete T, Pacheco-Coello R, Sánchez-Timm E, Santos-Ordóñez E. Plant Promoters: Their Identification, Characterization, and Role in Gene Regulation. Genes. 2023; 14(6):1226. https://doi.org/10.3390/genes14061226
Chicago/Turabian StyleVillao-Uzho, Liliana, Tatiana Chávez-Navarrete, Ricardo Pacheco-Coello, Eduardo Sánchez-Timm, and Efrén Santos-Ordóñez. 2023. "Plant Promoters: Their Identification, Characterization, and Role in Gene Regulation" Genes 14, no. 6: 1226. https://doi.org/10.3390/genes14061226
APA StyleVillao-Uzho, L., Chávez-Navarrete, T., Pacheco-Coello, R., Sánchez-Timm, E., & Santos-Ordóñez, E. (2023). Plant Promoters: Their Identification, Characterization, and Role in Gene Regulation. Genes, 14(6), 1226. https://doi.org/10.3390/genes14061226