


Seasonal Developing Xylem Transcriptome Analysis of Pinus densiflora Unveils Novel Insights for Compression Wood Formation




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Wood-Forming Tissue Sampling and RNA Sequencing
2.2. Transcript Assembly, Abundance Estimation, and Annotation
2.3. Differentially Expressed Gene and Gene Ontology Analysis
2.4. Quantitative Real Time PCR (qRT-PCR) and Semi-Quantitative Reverse Transcription PCR (RT-PCR)
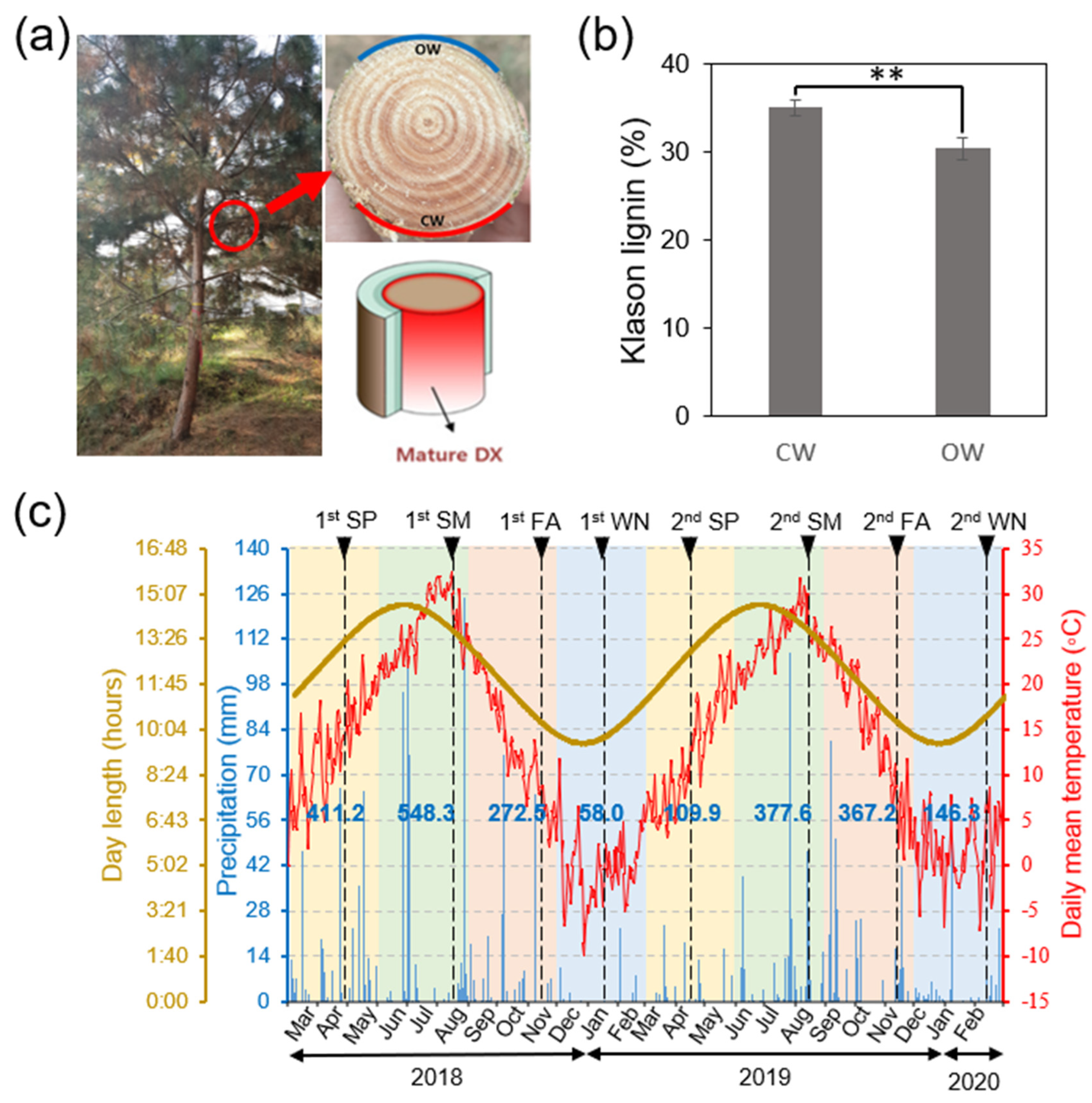
2.5. Klason Lignin Quantification
2.6. Transient Overexpression Analysis by the Agrobacteria-Infiltration Method and Laccase Activity Assays
3. Results
3.1. Analyzing the Seasonal Xylem Development in P. densiflora
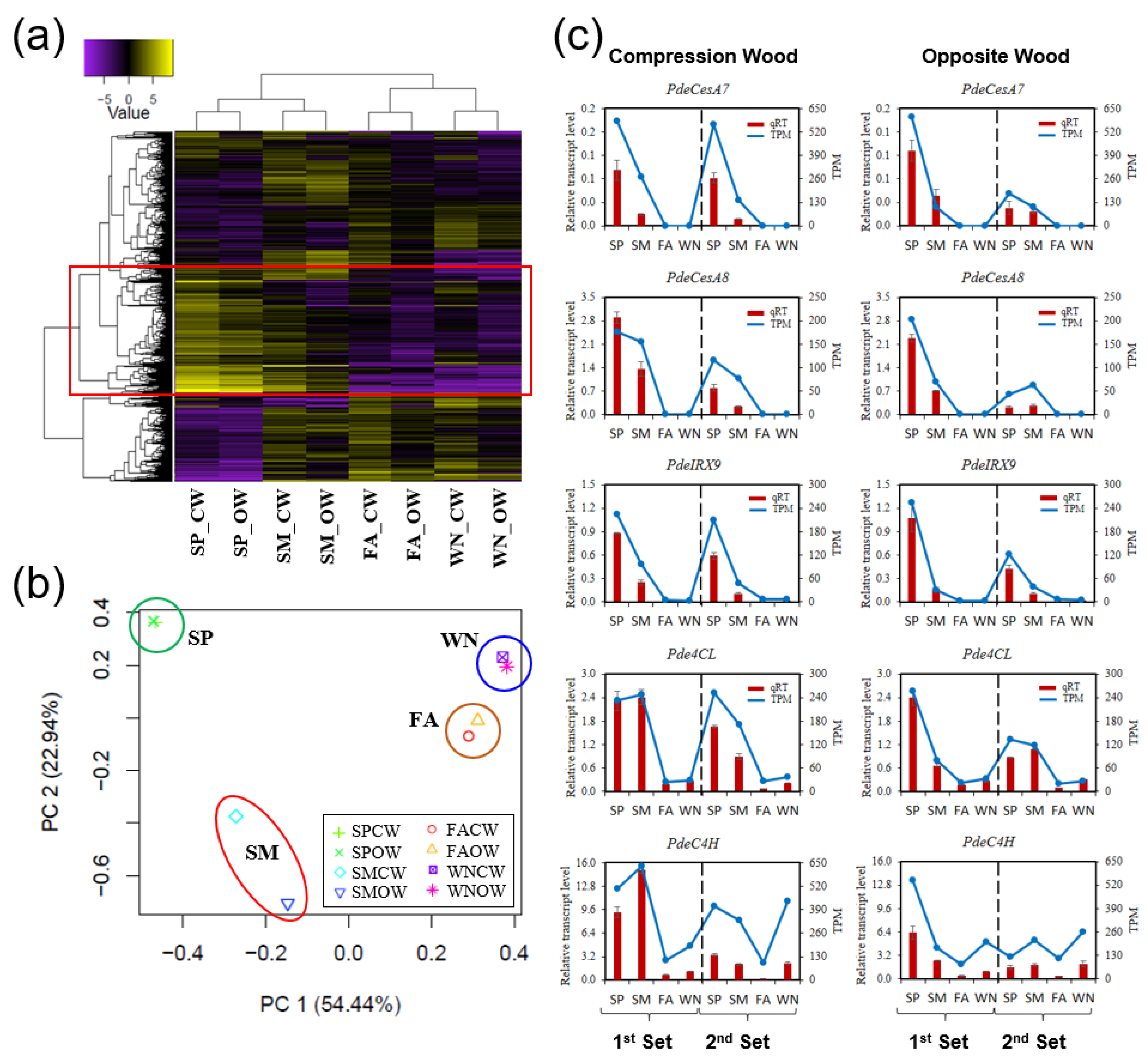
3.2. Building the Seasonal Transcriptomes of the Developing Xylem of P. densiflora
3.3. Coordinated Gene Expression Shapes Seasonal CW and OW Formation
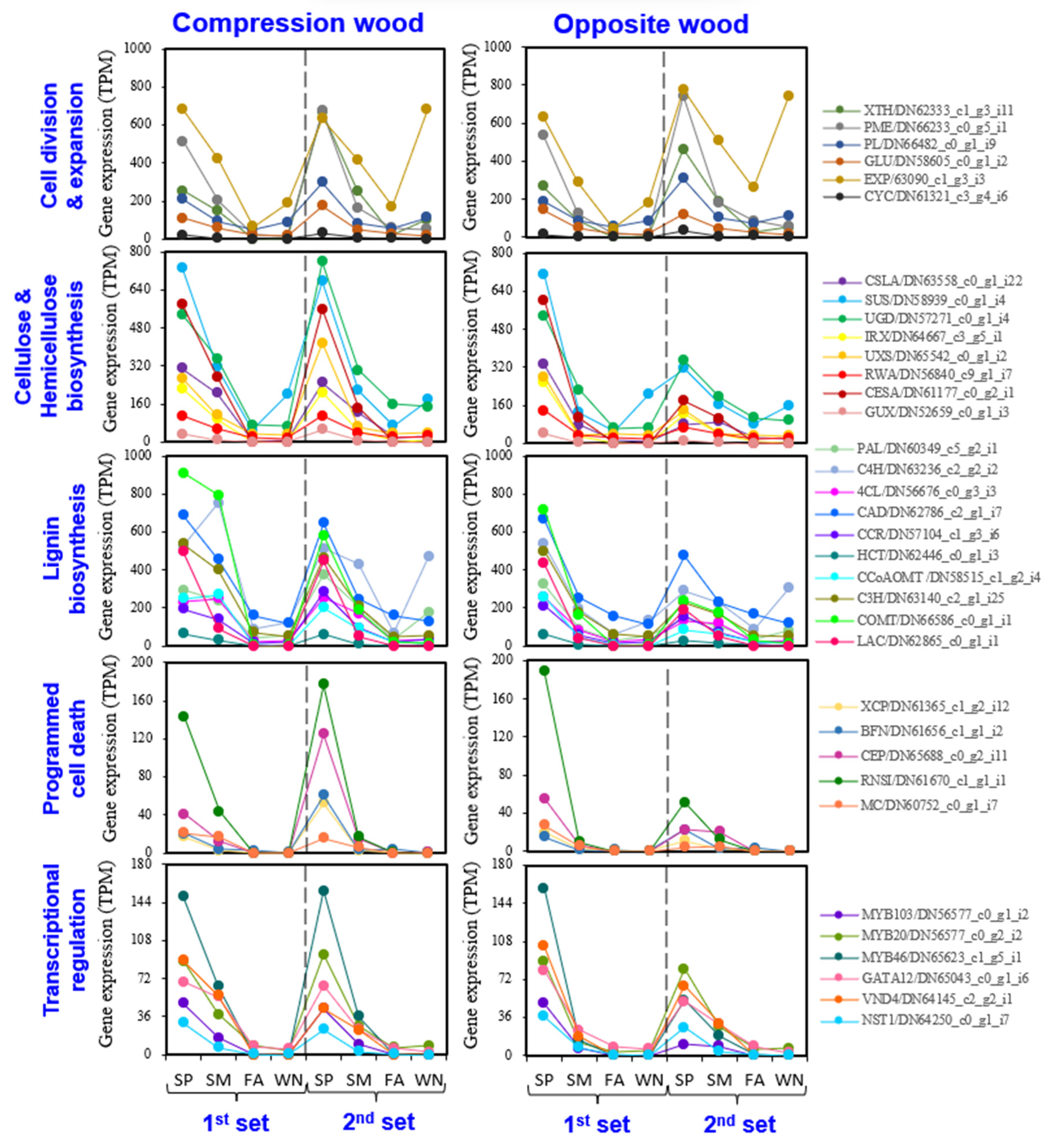
3.4. Evaluating Seasonal Variations in Gene Expression Involved in Wood Formation
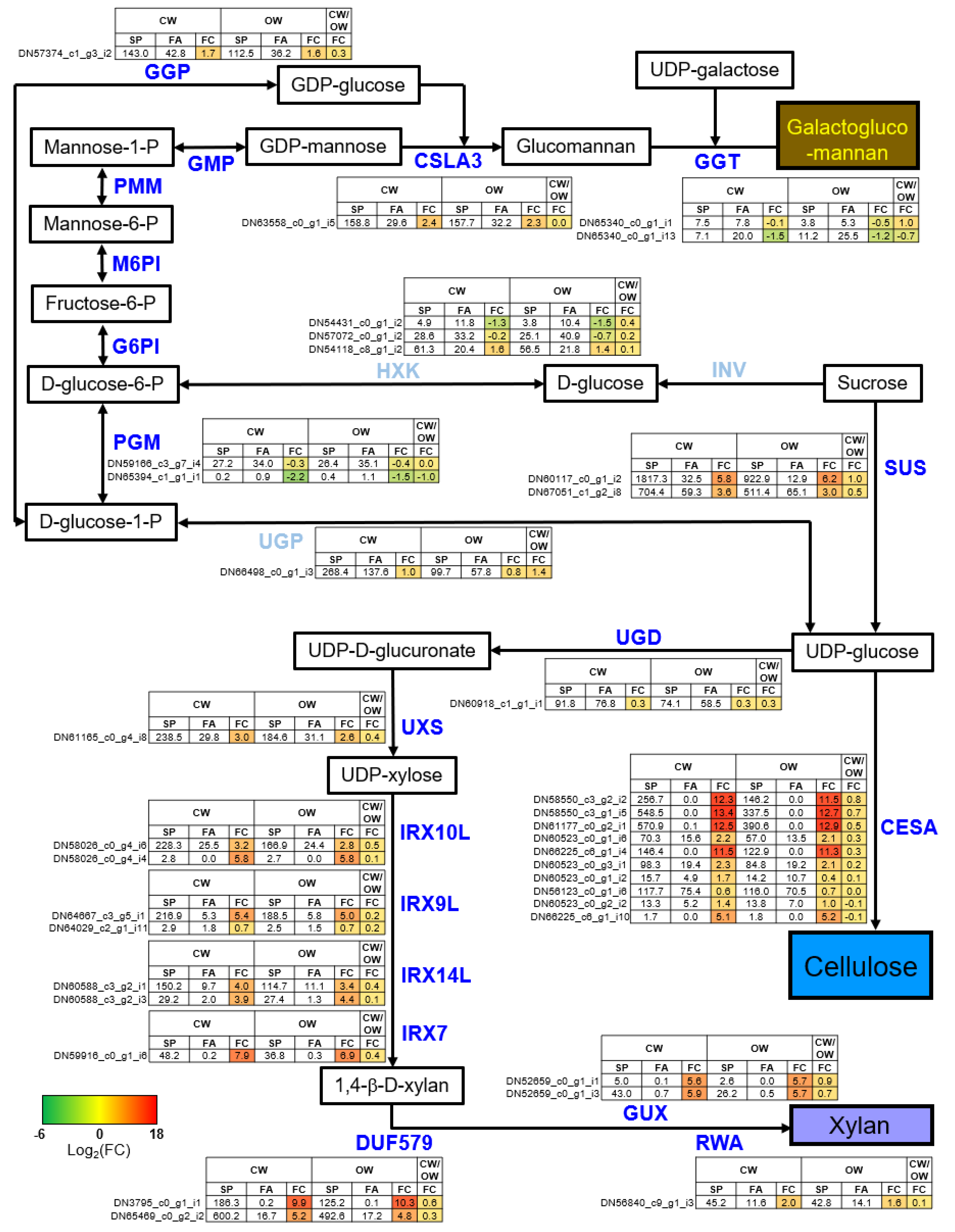
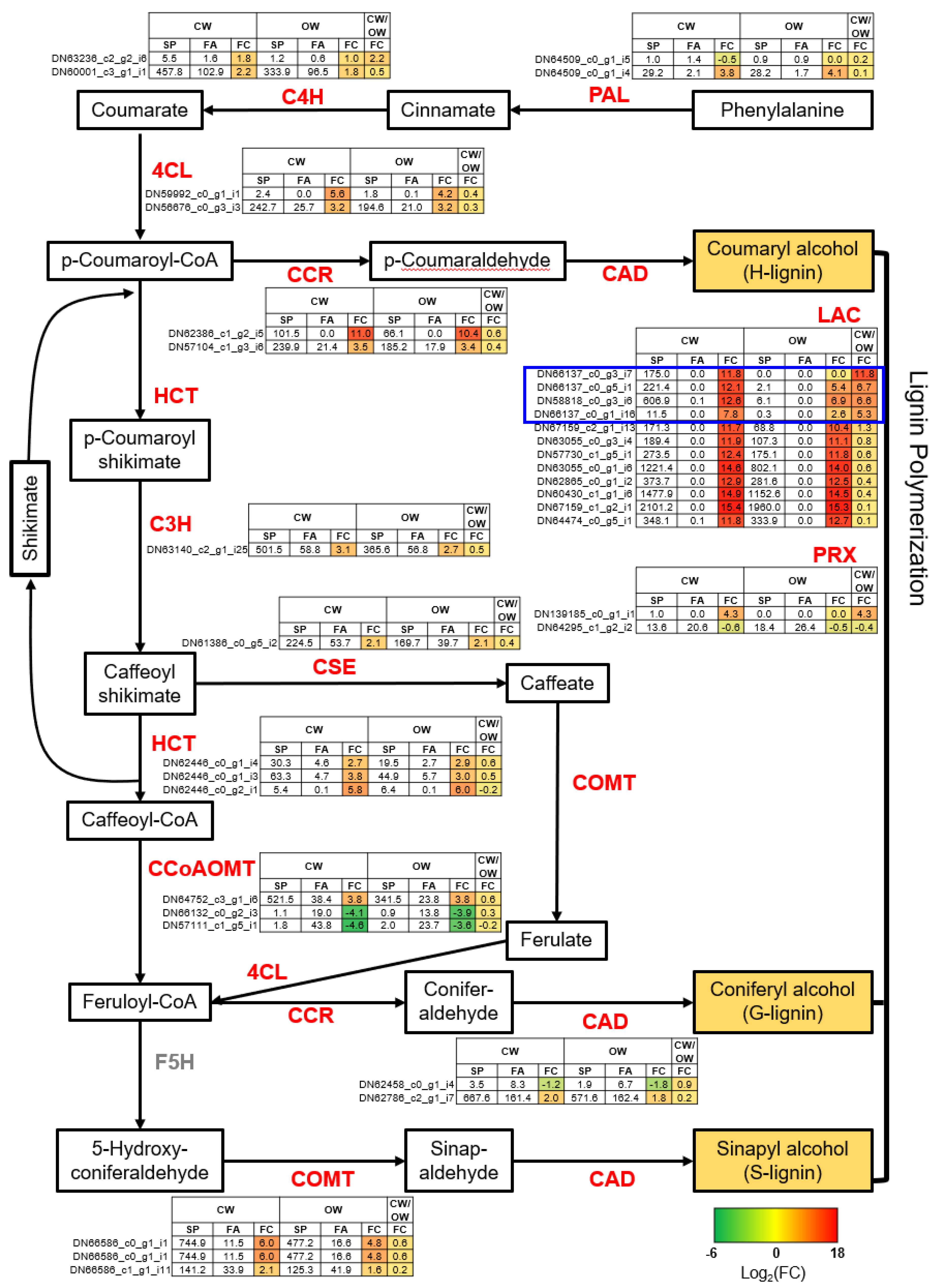
3.5. Uncovering Key Genes in the SCW Biosynthesis Crucial for CW Formation in P. densiflora
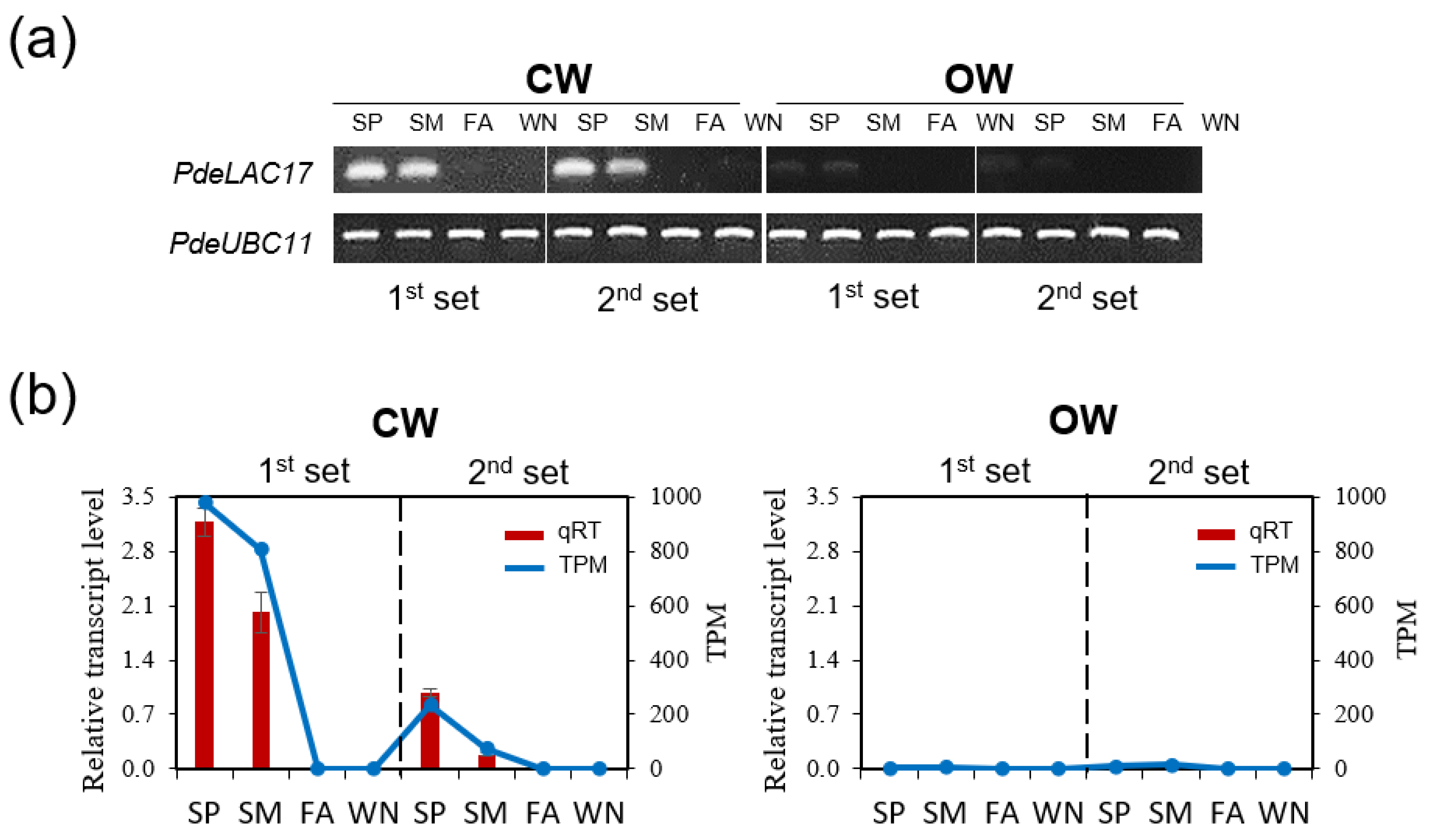
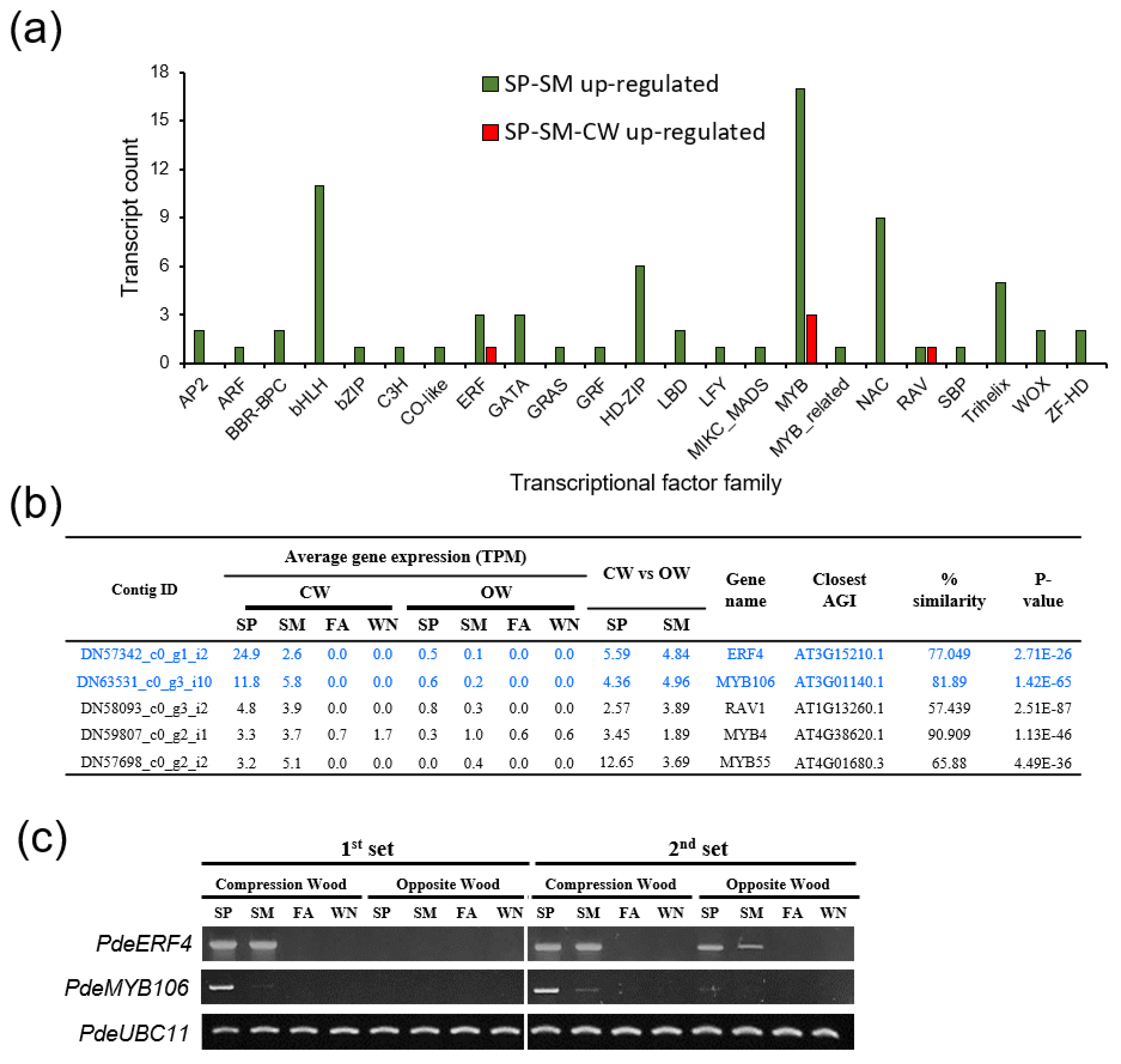
3.6. Identifying Differentially Expressed Genes Involved in CW Formation
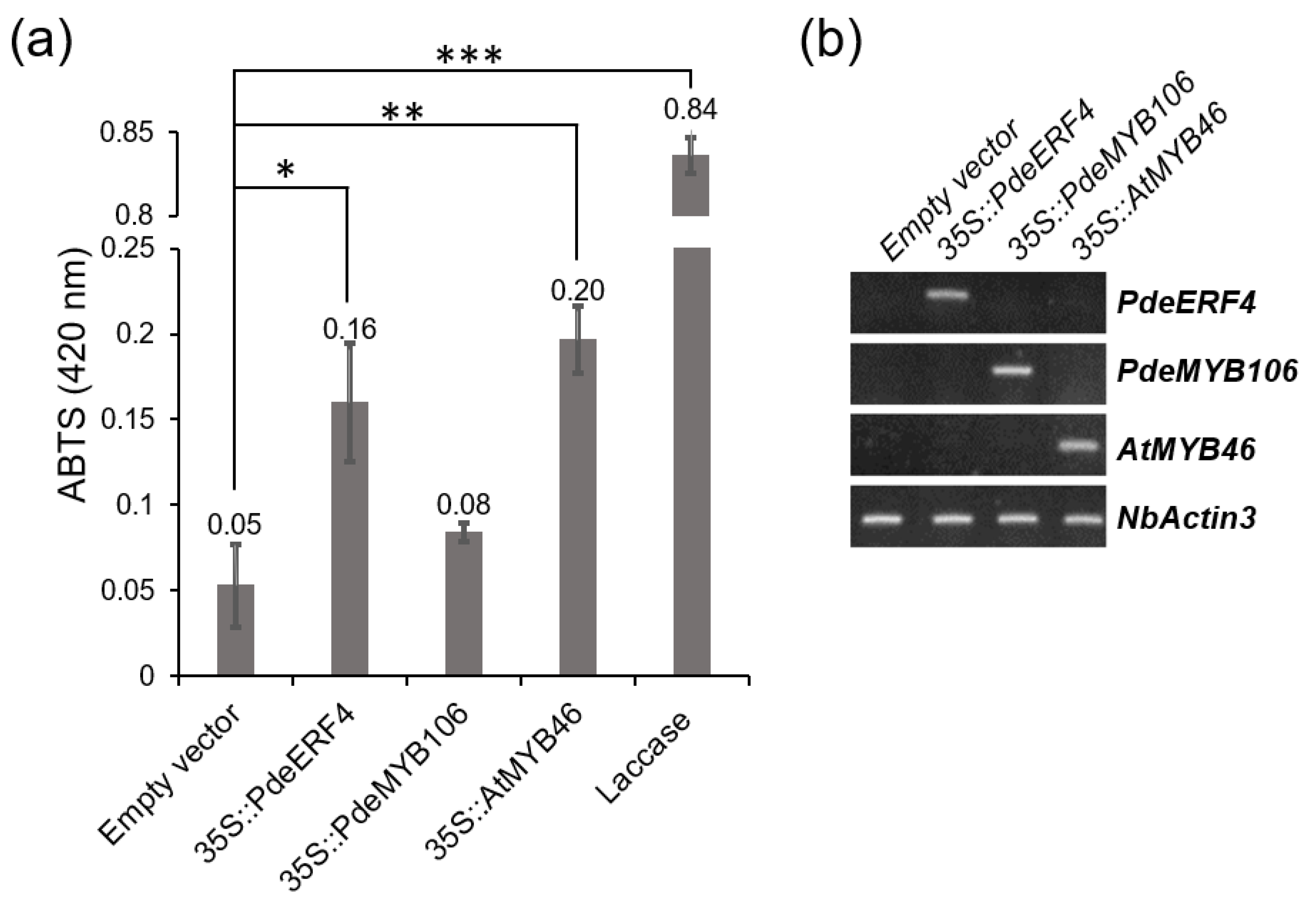
3.7. Functional Examination of PdeERF4 and PdeMYB106 as Possible Regulators of Laccase Activity
4. Discussion
4.1. Seasonal Variation in Gene Expression in the CW and OW Formation
4.2. Hormonal Regulation of Wood Formation in Seasonal Change
4.3. Identifying Key Genes in Compression Wood Formation in P. densiflora
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Ragauskas, A.; Beckham, G.; Biddy, M.; Chandra, R.; Chen, F.; Davis, M.; Davison, B.; Dixon, R.; Gilna, P.; Keller, M.; et al. Lignin Valorization: Improving Lignin Processing in the Biorefinery. Science 2014, 344, 1246843. [Google Scholar]
- Isikgor, F.H.; Becer, C.R. Lignocellulosic biomass: A sustainable platform for the production of bio-based chemicals and polymers. Polym. Chem. 2015, 6, 4497–4559. [Google Scholar]
- Zhang, J.; Nieminen, K.; Serra, J.A.A.; Helariutta, Y. The formation of wood and its control. Curr. Opin. Plant Biol. 2014, 17, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.H.; Tran, T.N.A.; Cho, J.S.; Park, E.J.; Lee, H.; Kim, D.G.; Hwang, S.; Ko, J.H. Wood transcriptome analysis of Pinus densiflora identifies genes critical for secondary cell wall formation and NAC transcription factors involved in tracheid formation. Tree Physiol. 2021, 41, 1289–1305. [Google Scholar] [CrossRef]
- Ko, J.H.; Jeon, H.W.; Kim, W.C.; Han, K.H. The MYB46/MYB83-mediated transcriptional regulatory program is a gatekeeper of secondary wall biosynthesis. Ann. Bot. 2014, 114, 1099–1107. [Google Scholar] [CrossRef]
- Kim, M.H.; Cho, J.S.; Jeon, H.W.; Sangsawang, K.; Shim, D.; Choi, Y.I.; Park, E.J.; Lee, H.; Ko, J.H. Wood transcriptome profiling identifies critical pathway genes of secondary wall biosynthesis and novel regulators for vascular cambium development in populus. Genes 2019, 10, 690. [Google Scholar] [CrossRef] [PubMed]
- Edlin, H.L. The summerwood: Springwood ratio in conifers: How it arises, varies, and affects end use for timber, board, and paper. For. Int. J. For. Res. 1965, 38, 91–112. [Google Scholar] [CrossRef]
- Nguyen, T.T.T.; Bae, E.K.; Tran, T.N.A.; Lee, H.; Ko, J.H. Exploring the Seasonal Dynamics and Molecular Mechanism of Wood Formation in Gymnosperm Trees. Int. J. Mol. Sci. 2023, 24, 8624. [Google Scholar]
- Ren, P.; Rossi, S.; Gricar, J.; Liang, E.; Cufar, K. Is precipitation a trigger for the onset of xylogenesis in Juniperus przewalskii on the north-eastern Tibetan Plateau? Ann Bot. 2015, 115, 629–639. [Google Scholar] [CrossRef]
- Ren, P.; Rossi, S.; Camarero, J.J.; Ellison, A.M.; Liang, E.; Peñuelas, J. Critical temperature and precipitation thresholds for the onset of xylogenesis of Juniperus przewalskii in a semi-arid area of the north-eastern Tibetan Plateau. Ann. Bot. 2018, 121, 617–624. [Google Scholar] [CrossRef]
- Zhang, M.; Ji, C.; Zhu, J.; Wang, X.; Wang, D.; Han, W. Comparison of wood physical and mechanical traits between major gymnosperm and angiosperm tree species in China. Wood Sci. Technol. 2017, 51, 1405–1419. [Google Scholar] [CrossRef]
- Groover, A. Gravitropisms and reaction woods of forest trees—Evolution, functions and mechanisms. New Phytol. 2016, 211, 790–802. [Google Scholar] [CrossRef] [PubMed]
- Morales-Quintana, L.; Ramos, P. A Talk between Flavonoids and Hormones to Reorient the Growth of Gymnosperms. Int. J. Mol. Sci. 2021, 22, 12630. [Google Scholar] [CrossRef]
- Cai, M.; Huang, H.; Ni, F.; Tong, Z.; Lin, E.; Zhu, M. RNA-Seq analysis of differential gene expression in Betula luminifera xylem during the early stages of tension wood formation. PeerJ 2018, 6, e5427. [Google Scholar] [CrossRef]
- Xiao, Y.; Yi, F.; Ling, J.; Wang, Z.; Zhao, K.; Lu, N.; Qu, G.; Kong, L.; Ma, W.; Wang, J. Transcriptomics and Proteomics Reveal the Cellulose and Pectin Metabolic Processes in the Tension Wood (Non-G-Layer) of Catalpa bungei. IJMS 2020, 21, 1686. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Liu, J.; Yu, J.; Wang, Z.; Sun, Y.; Li, S.; Lin, Y.J.; Chiang, V.L.; Li, W.; Wang, J.P. Transcriptional reprogramming of xylem cell wall biosynthesis in tension wood. Plant Physiol. 2021, 186, 250–269. [Google Scholar]
- Meng, X.; Wang, Y.; Li, J.; Jiao, N.; Zhang, X.; Zhang, Y.; Chen, J.; Tu, Z. RNA Sequencing Reveals Phenylpropanoid Biosynthesis Genes and Transcription Factors for Hevea brasiliensis Reaction Wood Formation. Front. Genet. 2021, 12, 763841. [Google Scholar] [CrossRef]
- Xiao, Y.; Ling, J.; Yi, F.; Ma, W.; Lu, N.; Zhu, T.; Wang, J.; Zhao, K.; Yun, H. Transcriptomic, Proteomic, and Metabolic Profiles of Catalpa bungei Tension Wood Reveal New Insight into Lignin Biosynthesis Involving Transcription Factor Regulation. Front. Plant Sci. 2021, 12, 704262. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Yoshida, M.; Hiraide, H.; Ihara, K.; Yamamoto, H. Transcriptome Analysis of Reaction Wood in Gymnosperms by Next-Generation Sequencing. Am. J. Plant Sci. 2014, 5, 2785–2798. [Google Scholar] [CrossRef]
- Li, X.; Yang, X.; Wu, H.X. Transcriptome profiling of radiata pine branches reveals new insights into reaction wood formation with implications in plant gravitropism. BMC Genom. 2013, 14, 768. [Google Scholar] [CrossRef] [PubMed]
- Villalobos, D.P.; Díaz-Moreno, S.M.; el-Said, S.S.; Cañas, R.A.; Osuna, D.; Van Kerckhoven, S.H.; Bautista, R.; Claros, M.G.; Cánovas, F.M.; Cantón, F.R. Reprogramming of gene expression during compression wood formation in pine: Coordinated modulation of S-adenosylmethionine, lignin and lignan related genes. BMC Plant Biol. 2012, 12, 100. [Google Scholar] [CrossRef] [PubMed]
- Hiraide, H.; Tobimatsu, Y.; Yoshinaga, A.; Lam, P.Y.; Kobayashi, M.; Matsushita, Y.; Fukushima, K.; Takabe, K. Localised laccase activity modulates distribution of lignin polymers in gymnosperm compression wood. New Phytol. 2021, 230, 2186–2199. [Google Scholar] [CrossRef] [PubMed]
- Lourenço, A.; Pereira, H. Compositional Variability of Lignin in Biomass. In Lignin-Trends and Applications; BoD—Books on Demand: Norderstedt, Germany, 2018; 308p. [Google Scholar]
- Zhang, Z.; Wang, H.; Wu, J.; Jin, Y.; Xiao, S.; Li, T.; Liu, X.; Zhang, H.; Zhang, Z.; Su, J.; et al. Comprehensive Transcriptome Analysis of Stem-Differentiating Xylem Upon Compression Stress in Cunninghamia Lanceolata. Front Genet. 2022, 3, 843269. [Google Scholar] [CrossRef]
- Kocaturk, E.; Salan, T.; Ozcelik, O.; Alma, M.H.; Candan, Z. Recent advances in lignin-based biofuel production. Energies 2023, 16, 3382. [Google Scholar] [CrossRef]
- Nystedt, B.; Street, N.R.; Wetterbom, A.; Zuccolo, A.; Lin, Y.-C.; Scofield, D.G.; Vezzi, F.; Delhomme, N.; Giacomello, S.; Alexeyenko, A. The Norway spruce genome sequence and conifer genome evolution. Nature 2013, 497, 579–584. [Google Scholar] [CrossRef]
- Kirst, M.; Johnson, A.F.; Baucom, C.; Ulrich, E.; Hubbard, K.; Staggs, R.; Paule, C.; Retzel, E.; Whetten, R.; Sederoff, R. Apparent homology of expressed genes from wood-forming tissues of loblolly pine (Pinus taeda L.) with Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2003, 100, 7383–7388. [Google Scholar] [CrossRef]
- Jokipii-Lukkari, S.; Sundell, D.; Nilsson, O.; Hvidsten, T.R.; Street, N.R.; Tuominen, H. NorWood: A gene expression resource for evo-devo studies of conifer wood development. New Phytol. 2017, 216, 482–494. [Google Scholar] [CrossRef]
- Birol, I.; Raymond, A.; Jackman, S.D.; Pleasance, S.; Coope, R.; Taylor, G.A.; Yuen, M.M.S.; Keeling, C.I.; Brand, D.; Vandervalk, B.P.; et al. Assembling the 20 Gb white spruce (Picea glauca) genome from whole-genome shotgun sequencing data. Bioinformatics 2013, 29, 1492–1497. [Google Scholar] [CrossRef]
- Neale, D.B.; Kremer, A. Forest tree genomics: Growing resources and applications. Nat. Rev. Genet. 2011, 12, 111–122. [Google Scholar] [PubMed]
- Stevens, K.A.; Wegrzyn, J.L.; Zimin, A.; Puiu, D.; Crepeau, M.; Cardeno, C.; Paul, R.; Gonzalez-Ibeas, D.; Koriabine, M.; Holtz-Morris, A.E.; et al. Sequence of the sugar pine megagenome. Genetics 2016, 204, 1613–1626. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.H.; Kim, J.C.; Kim, K.J.; Son, Y.S.; Young, S.W.; Han, J.S. Seasonal variations of monoterpene emissions from Pinus densiflora in East Asia. Chemosphere. 2008, 73, 470–478. [Google Scholar] [CrossRef]
- Choung, Y.; Lee, J.; Cho, S.; Noh, J. Review on the succession process of Pinus densiflora forests in South Korea: Progressive and disturbance-driven succession. J. Ecol. Environ. 2020, 44, 16. [Google Scholar] [CrossRef]
- Zhang, C.H.; Fujii, T.; Abe, H.; Fujiwara, T.; Fujita, M.; Takabe, K. Anatomical Features of Radial Resin Canals In Pinus Densiflora. IAWA J. 2008, 29, 179–187. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De novo transcript sequence reconstruction from RNA-seq using the Trinity platform for reference generation and analysis. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef]
- Li, W.; Godzik, A. Cd-hit: A fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009, 10, R25. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinfom. 2011, 12, 323. [Google Scholar] [CrossRef]
- Ge, S.X.; Son, E.W.; Yao, R. iDEP: An integrated web application for differential expression and pathway analysis of RNA-Seq data. BMC Bioinfom. 2018, 19, 534. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID Bioinformatics Resources. Nat. Protoc. 2009, 4, 44–45. [Google Scholar] [CrossRef]
- Dence, C.W. The determination of lignin. In Methods in Lignin Chemistry; Springer: Berlin/Heidelberg, Germany, 1992; pp. 33–61. [Google Scholar]
- Karimi, M.; Inzé, D.; Depicker, A. GATEWAY™ vectors for Agrobacterium-mediated plant transformation. Trends Plant Sci. 2002, 7, 193–195. [Google Scholar] [CrossRef]
- Berthet, S.; Demont-Caulet, N.; Pollet, B.; Bidzinski, P.; Cézard, L.; Le Bris, P.; Borrega, N.; Hervé, J.; Blondet, E.; Balzergue, S.; et al. Disruption of LACCASE4 and 17 results in tissue-specific alterations to lignification of Arabidopsis thaliana stems. Plant Cell 2011, 23, 1124–1137. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Kumar, R.; Kaur, J.; Jain, S.; Kumar, A. Optimization of laccase production from Aspergillus flavus by design of experiment technique: Partial purification and characterization. J. Genet. Eng. Biotechnol. 2016, 14, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.-M.; Hu, Z.; Chang, H.-M.; Li, B. Micro Analytical Methods for Determination of Compression Wood Content in Loblolly Pine. J. Wood Chem. Technol. 2007, 27, 169–178. [Google Scholar] [CrossRef]
- Jokipii-Lukkari, S.; Delhomme, N.; Schiffthaler, B.; Mannapperuma, C.; Prestele, J.; Nilsson, O.; Street, N.R.; Tuominen, H. Transcriptional roadmap to seasonal variation in wood formation of Norway Spruce. Plant Physiol. 2018, 176, 2851–2870. [Google Scholar] [CrossRef] [PubMed]
- Palombo, C.; Fonti, P.; Lasserre, B.; Cherubini, P.; Marchetti, M.; Tognetti, R. Xylogenesis of compression and opposite wood in mountain pine at a Mediterranean tree line. Ann. For. Sci. 2018, 75, 93. [Google Scholar] [CrossRef]
- Huang, J.G.; Guo, X.; Rossi, S.; Zhai, L.; Yu, B.; Zhang, S.; Zhang, M. Intra-annual wood formation of subtropical Chinese red pine shows better growth in dry season than wet season. Tree Physiol. 2018, 38, 1225–1236. [Google Scholar] [CrossRef]
- Guo, X.; Huang, J.G.; Buttò, V.; Luo, D.; Shen, C.; Li, J.; Liang, H.; Zhang, S.; Hou, X.; Zhao, P.; et al. Auxin concentration and xylem production of Pinus massoniana in a subtropical forest in south China. Tree Physiol. 2022, 42, 317–324. [Google Scholar] [CrossRef]
- Yang, Y.; Xu, R.; Ma, C.J.; Vlot, A.C.; Klessig, D.F.; Pichersky, E. Inactive methyl indole-3-acetic acid ester can be hydrolyzed and activated by several esterases belonging to the AtMES esterase family of Arabidopsis. Plant Physiol. 2008, 147, 1034–1045. [Google Scholar] [CrossRef] [PubMed]
- Salomé, P.A.; To, J.P.; Kieber, J.J.; McClung, C.R. Arabidopsis response regulators ARR3 and ARR4 play cytokinin-independent roles in the control of circadian period. Plant Cell 2006, 18, 55–69. [Google Scholar] [CrossRef] [PubMed]
- Zentella, R.; Wang, Y.; Zahn, E.; Hu, J.; Jiang, L.; Shabanowitz, J.; Hunt, D.F.; Sun, T.P. SPINDLY O-fucosylates nuclear and cytoplasmic proteins involved in diverse cellular processes in plants. Plant Physiol 2023, 191, 1546–1560. [Google Scholar] [CrossRef] [PubMed]
- Cui, Q.; Gao, X.; Wang, L.; Jia, G. Ectopic expression of LhMYC2 increases susceptibility to Botrytis cinerea in Arabidopsis thaliana. Can. J. Plant Sci. 2020, 101, 328–340. [Google Scholar] [CrossRef]
- Ali, M.S.; Baek, K.H. Jasmonic Acid Signaling Pathway in Response to Abiotic Stresses in Plants. Int. J. Mol. Sci. 2020, 21, 621. [Google Scholar] [CrossRef]
- Andersson-Gunnerås, S.; Hellgren, J.M.; Björklund, S.; Regan, S.; Moritz, T.; Sundberg, B. Asymmetric expression of a poplar ACC oxidase controls ethylene production during gravitational induction of tension wood. Plant J. 2003, 34, 339–349. [Google Scholar] [CrossRef]
- Seyfferth, C.; Wessels, B.A.; Gorzsás, A.; Love, J.W.; Rüggeberg, M.; Delhomme, N.; Vain, T.; Antos, K.; Tuominen, H.; Sundberg, B.; et al. Ethylene Signaling Is Required for Fully Functional Tension Wood in Hybrid Aspen. Front Plant Sci. 2019, 26, 1101. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Tian, L.; Latoszek-Green, M.; Brown, D.; Wu, K. Arabidopsis ERF4 is a transcriptional repressor capable of modulating ethylene and abscisic acid responses. Plant Mol. Biol. 2005, 58, 585–596. [Google Scholar] [CrossRef]
- Chen, W.H.; Li, P.F.; Chen, M.K.; Lee, Y.I.; Yang, C.H. Forever Young Flower Negatively Regulates Ethylene Response DNA-Binding Factors by Activating an Ethylene-Responsive Factor to Control Arabidopsis Floral Organ Senescence and Abscission. Plant Physiol. 2015, 168, 1666–1683. [Google Scholar] [CrossRef]
- Zhu, L.; Liu, D.; Li, Y.; Li, N. Functional phosphoproteomic analysis reveals that a serine-62-phosphorylated isoform of ethylene response factor110 is involved in Arabidopsis bolting. Plant Physiol. 2013, 161, 904–917. [Google Scholar] [CrossRef]
- Lise, J.; Serge, B.; Julien, M.; Nathalie, D.-C.; Brice, A.; Philippe, L.-B.; Davy, B.; Leplé, J.-C.; Catherine, L. Identification of laccases involved in lignin polymerization and strategies to deregulate their expression in order to modify lignin content in Arabidopsis and poplar. BMC Proc. 2011, 5, 1. [Google Scholar] [CrossRef]
- McDougall, G.J. A comparison of proteins from the developing xylem of compression and non-compression wood of branches of sitka spruce (Picea sitchensis) reveals a differentially expressed laccase. J. Exp. Bot. 2000, 51, 1395–1401. [Google Scholar] [PubMed]
- Hiraide, H.; Yoshida, M.; Ihara, K.; Sato, S.; Yamamoto, H. High lignin deposition on the outer region of the secondary wall middle layer in compression wood matches the expression of a laccase gene in Chamaecyparis obtuse. JPBR 2014, 3, 87–100. [Google Scholar]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, T.T.T.; Kim, M.-H.; Park, E.-J.; Lee, H.; Ko, J.-H. Seasonal Developing Xylem Transcriptome Analysis of Pinus densiflora Unveils Novel Insights for Compression Wood Formation. Genes 2023, 14, 1698. https://doi.org/10.3390/genes14091698
Nguyen TTT, Kim M-H, Park E-J, Lee H, Ko J-H. Seasonal Developing Xylem Transcriptome Analysis of Pinus densiflora Unveils Novel Insights for Compression Wood Formation. Genes. 2023; 14(9):1698. https://doi.org/10.3390/genes14091698
Chicago/Turabian StyleNguyen, Thi Thu Tram, Min-Ha Kim, Eung-Jun Park, Hyoshin Lee, and Jae-Heung Ko. 2023. "Seasonal Developing Xylem Transcriptome Analysis of Pinus densiflora Unveils Novel Insights for Compression Wood Formation" Genes 14, no. 9: 1698. https://doi.org/10.3390/genes14091698
APA StyleNguyen, T. T. T., Kim, M. -H., Park, E. -J., Lee, H., & Ko, J. -H. (2023). Seasonal Developing Xylem Transcriptome Analysis of Pinus densiflora Unveils Novel Insights for Compression Wood Formation. Genes, 14(9), 1698. https://doi.org/10.3390/genes14091698