
RNA-Seq Identified Putative Genes Conferring Photosynthesis and Root Development of Melon under Salt Stress

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Material and Salt Stress
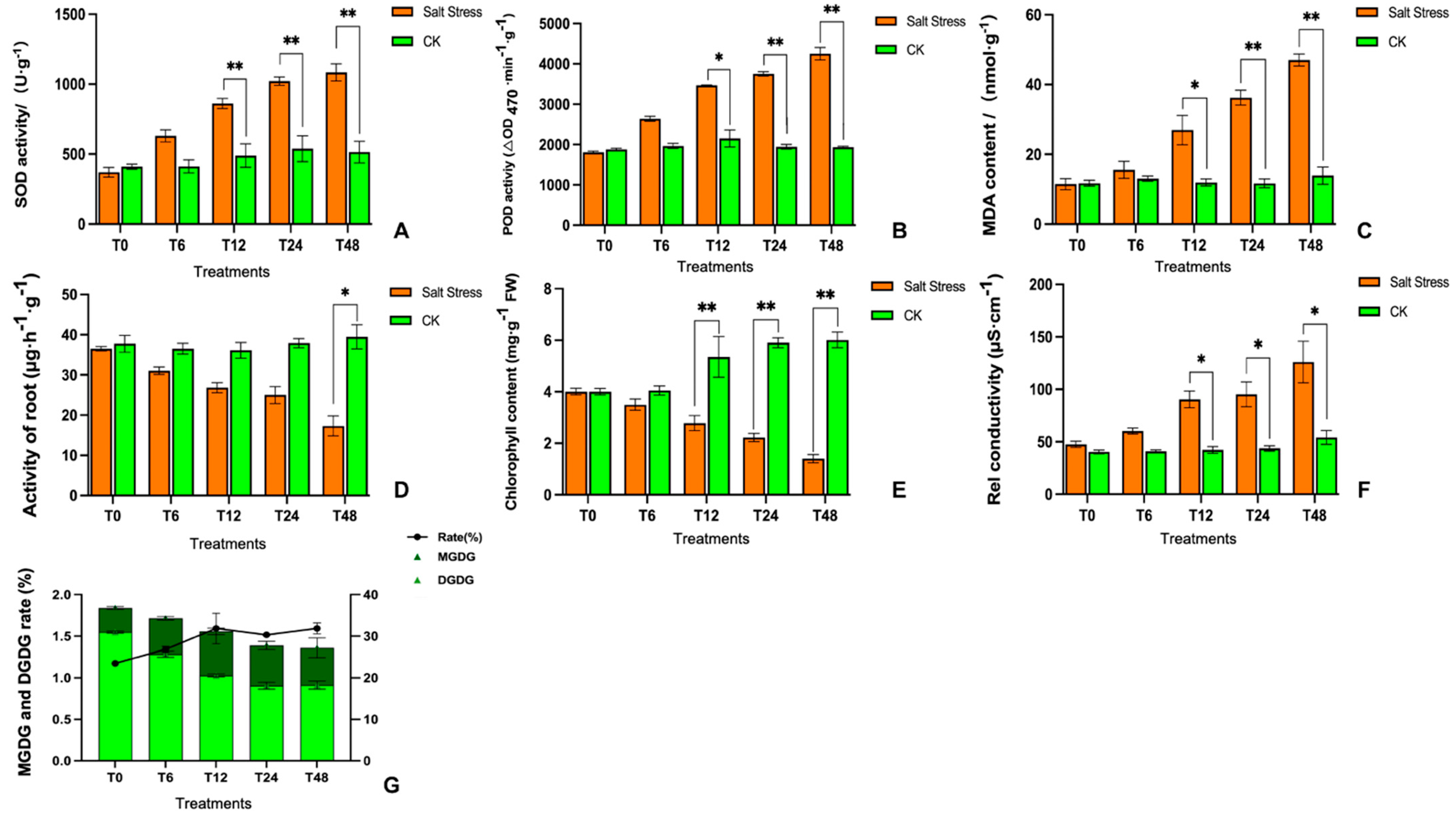
2.2. Evaluation of Metabolic Activity of Roots and Chlorophyll Contents of Leaves
2.3. Identification of Antioxidant Enzyme Activity and Cell Membrane Permeability
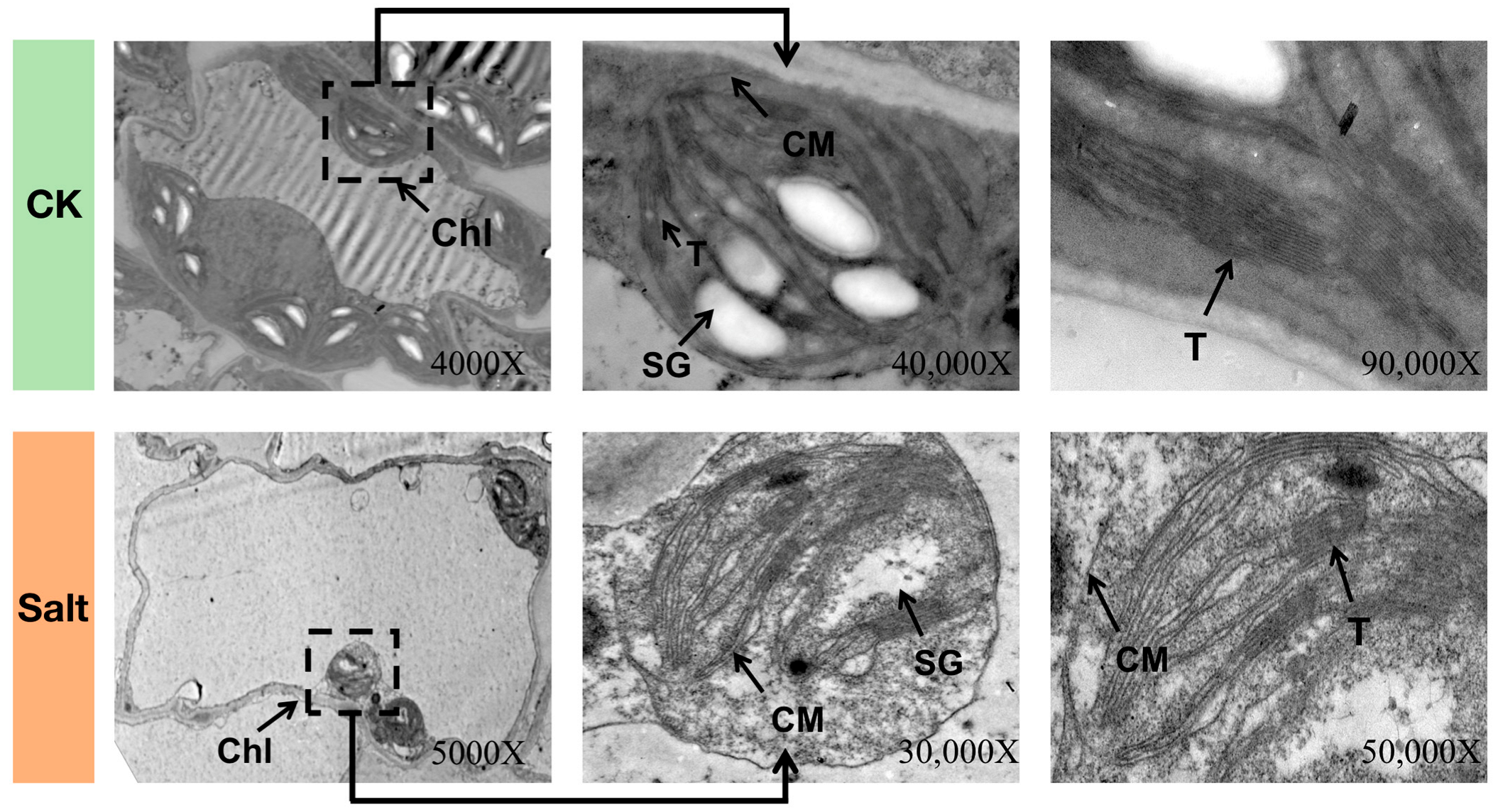
2.4. Analysis of Transmission Electron Microscopy (TEM)
2.5. Analysis of Transcriptomic Sequencing
2.6. DEG Identification and Analysis
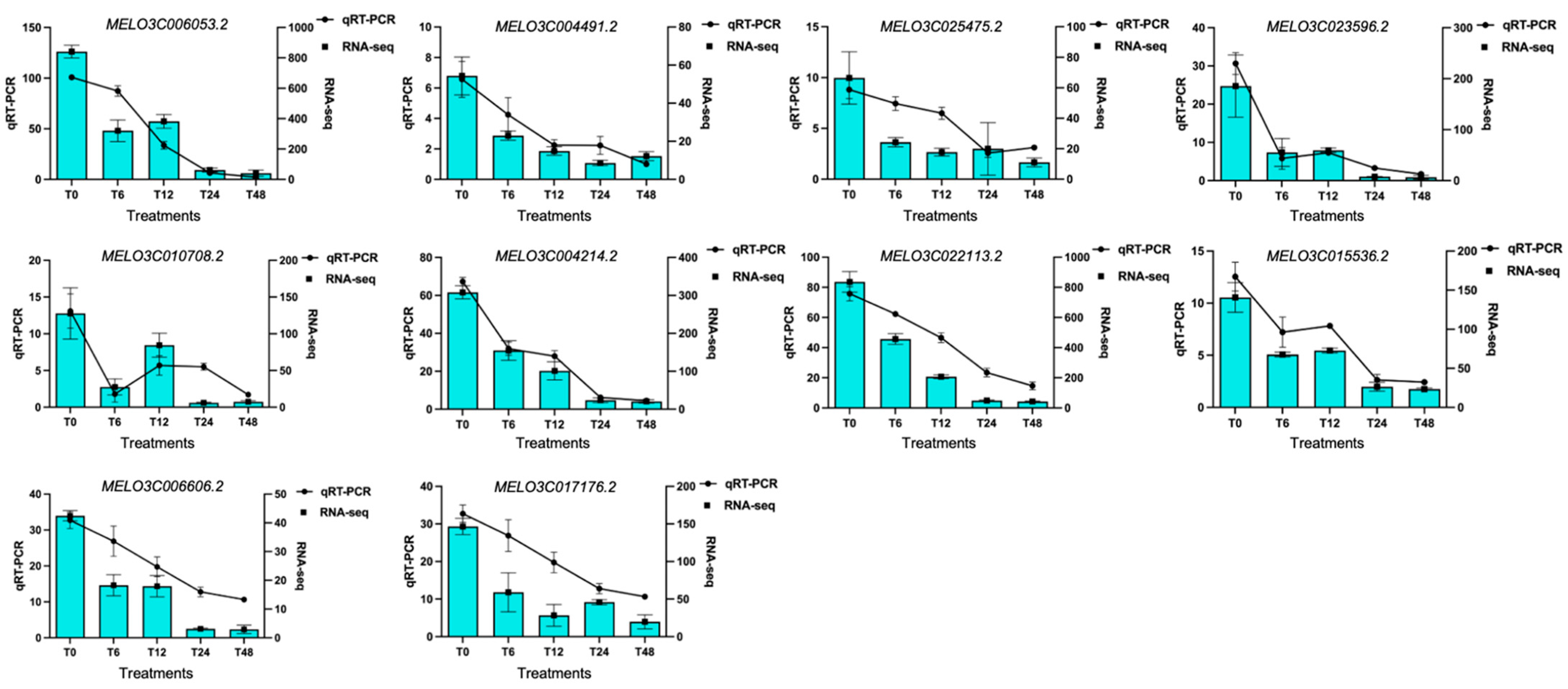
2.7. qRT-PCR Analysis
2.8. WGCN and Pathway Analysis
3. Results
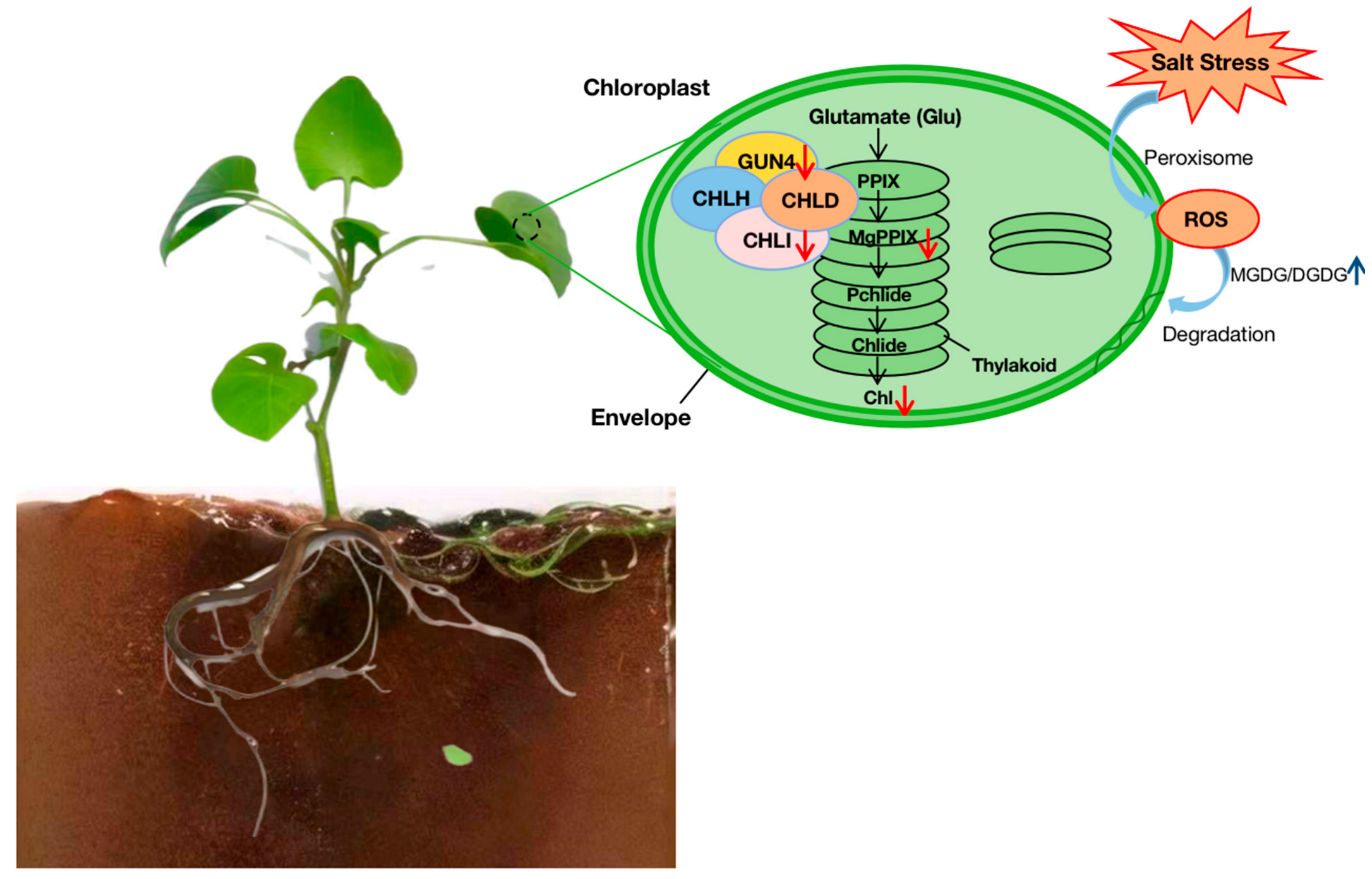
3.1. Phenotypic and Cytological Observations of Melon Seedlings
3.2. Physiological Variations in Leaves of Melon Seedling
3.3. Overview of Transcriptomic Data
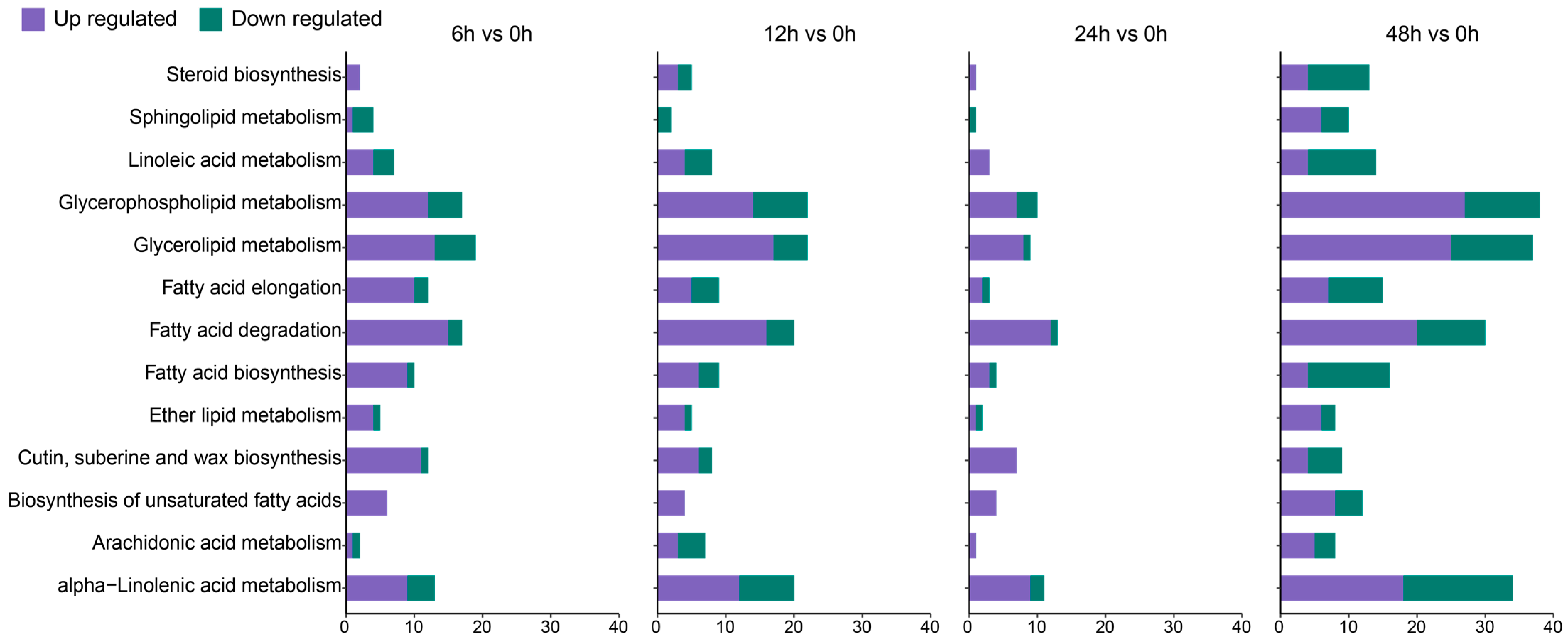
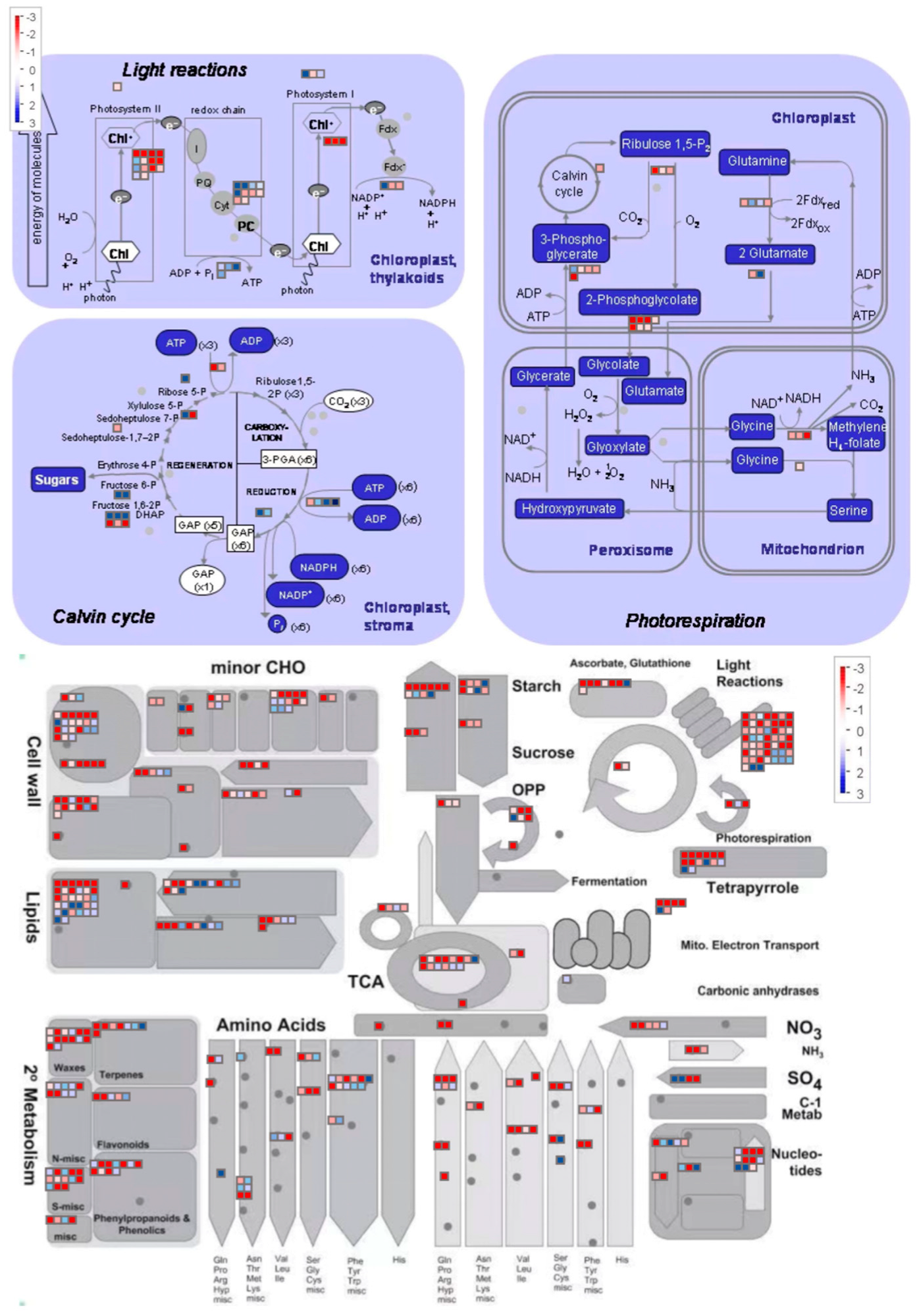
3.4. DEGs Associated with Key Biological Processes
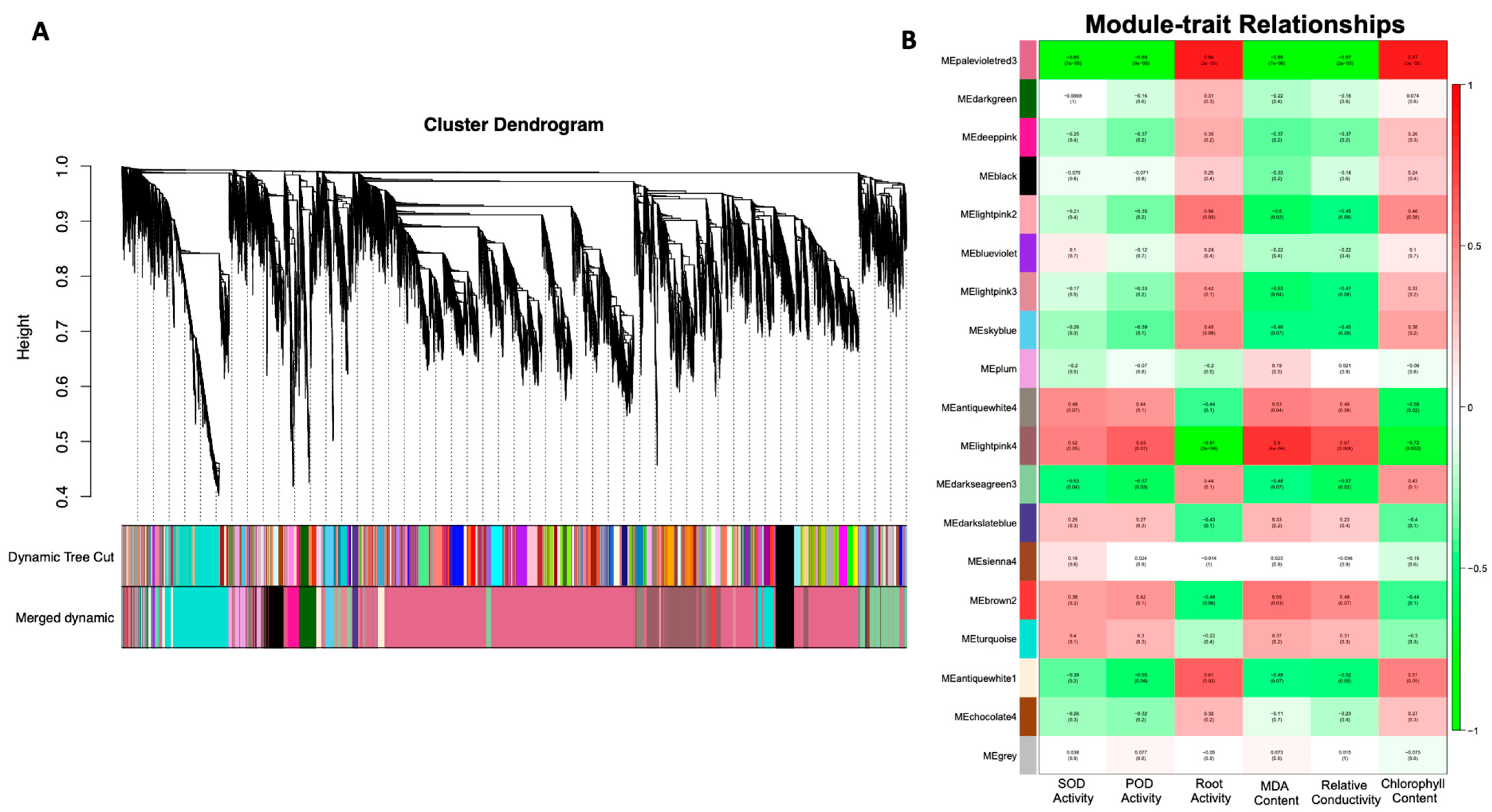
3.5. Identification of WGCN-Associated DEGs under Salt Stress
4. Discussion
4.1. Synergistic Network Response of Lipid Metabolism and Photosynthesis in Membrane
4.2. Enzymes, Non-Enzymatic Factors, and Hormones Inducing Retarded Root Growth
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef]
- Li, N.; Cao, B.L.; Chen, Z.J.; Xu, K. Root morphology ion absorption and antioxidative defense system of two Chinese cabbage cultivars (Brassica rapa L.) reveal the different adaptation mechanisms to salt and alkali stress. Protoplasma 2022, 259, 385–398. [Google Scholar] [CrossRef]
- Jacoby, R.P.; Millar, A.H.; Taylor, N.L. Wheat mitochondrial proteomes provide new links between antioxidant defense and plant salinity tolerance. J. Proteome Res. 2010, 9, 6595–6604. [Google Scholar] [CrossRef]
- Niewiadomska, E.; Wiciarz, M. Adaptations of chloroplastic metabolism in halophytic plants. In Progress in Botany; Lüttge, U., Beyschlag, W., Eds.; Springer: Cham, Switzerland, 2015; Volume 76, pp. 177–193. [Google Scholar]
- He, Y.; Fu, J.L.; Yu, C.L.; Wang, X.M.; Jiang, Q.S.; Hong, J.; Lu, K.X.; Xue, G.P.; Yan, C.Q.; James, A.; et al. Increasing cyclic electron flow is related to naþ sequestration into vacuoles for salt tolerance in soybean. J. Exp. Bot. 2015, 66, 6877–6889. [Google Scholar] [CrossRef]
- Chinnusamy, V.; Zhu, J.H.; Zhu, J.K. Cold stress regulation of gene expression in plants. Trends Plant Sci. 2007, 12, 444–451. [Google Scholar] [CrossRef]
- Borrell, J.H.; Domènech, Ò.; Keough, K.M. Membrane Protein-Lipid Interactions: Physics and Chemistry in the Bilayer; Springer: Cham, Switzerland, 2016. [Google Scholar]
- Reszczyńska, E.; Hanaka, A. Lipids composition in plant membranes. Cell Biochem. Biophys. 2020, 78, 401–414. [Google Scholar] [CrossRef]
- Guo, Q.; Liu, L.; Barkla, B.J. Membrane lipid remodeling in response to salinity. Int. J. Mol. Sci. 2019, 20, 4264. [Google Scholar] [CrossRef]
- Kalisch, B.; Dörmann, P.; Hölzl, G. DGDG and glycolipids in plants and algae. In Lipids in Plant and Algae Development; Nakamura, Y., Li-Beisson, Y., Eds.; Springer: Cham, Switzerland, 2016; Volume 86, pp. 51–83. [Google Scholar]
- Negi, J.; Munemasa, S.; Song, B.; Tadakuma, R.; Fujita, M.; Azoulay-Shemer, T.; Engineer, C.B.; Kusumi, K.; Nishida, I.; Schroeder, J.I.; et al. Eukaryotic lipid metabolic pathway is essential for functional chloroplasts and CO2 and light responses in Arabidopsis guard cells. Proc. Natl. Acad. Sci. USA 2018, 115, 9038–9043. [Google Scholar] [CrossRef]
- Li, Q.; Zheng, Q.; Shen, W.Y.; Cram, D.; Fowler, D.B.; Wei, Y.D.; Zou, J.T. Understanding the biochemical basis of temperature-induced lipid pathway adjustments in plants. Plant Cell 2015, 27, 86–103. [Google Scholar] [CrossRef]
- Campos, P.S.; Quartin, V.N.; Ramalho, J.C.; Nunes, M.A. Electrolyte leakage and lipid degradation account for cold sensitivity in leaves of Coffea sp. plants. J. Plant Physiol. 2003, 160, 283–292. [Google Scholar] [CrossRef]
- Moellering, E.R.; Muthan, B.; Benning, C. Freezing tolerance in plants requires lipid remodeling at the outer chloroplast membrane. Science 2010, 330, 226–228. [Google Scholar] [CrossRef]
- Lamine, M.; Gargouri, M.; Mliki, A. Identifification of the NaCl-responsive metabolites in Citrus roots: A lipidomic and volatomic signature. Plant Signal. Behav. 2020, 15, 1777376. [Google Scholar] [CrossRef]
- Li, W.Q.; Wang, R.P.; Li, M.Y.; Li, L.X.; Wang, C.M.; Welti, R.; Wang, X.M. Differential degradation of extraplastidic and plastidic lipids during freezing and post-freezing recovery in Arabidopsis thaliana. J. Biol. Chem. 2008, 283, 461–468. [Google Scholar] [CrossRef]
- Wang, Y.N.; Li, K.X.; Li, X. Auxin redistribution modulates plastic development of root system architecture under salt stress in Arabidopsis thaliana. J. Plant Physiol. 2009, 166, 1637–1645. [Google Scholar] [CrossRef] [PubMed]
- You, J.; Chan, Z.L. ROS regulation during abiotic stress responses in crop plants. Front. Plant Sci. 2015, 6, 1092. [Google Scholar] [CrossRef]
- Tommasino, E.; Griffa, S.; Grunberg, K.; Ribotta, A.; Colomba, E.L.; Carloni, E.; Quiroga, M.; Luna, C.M. Malondialdehyde content as a potential biochemical indicator of tolerant Cenchrus ciliaris L. genotypes under heat stress treatment. Grass Forage Sci. 2012, 67, 456–459. [Google Scholar] [CrossRef]
- Zhang, F.; Zhu, G.Z.; Du, L.; Shang, X.G.; Cheng, C.Z.; Yang, B.; Hu, Y.; Cai, C.P.; Guo, W.Z. Genetic regulation of salt stress tolerance revealed by RNASeq in cotton diploid wild species, Gossypium davidsonii. Sci. Rep. 2016, 6, 20582. [Google Scholar] [CrossRef]
- Rose, J.K.C.; Braam, J.; Fry, S.C.; Nishitani, K. The XTH family of enzymes involved in xyloglucan endotransglucosylation and endohydrolysis: Current perspectives and a new unifying nomenclature. Plant Cell Physiol. 2002, 43, 1421–1435. [Google Scholar] [CrossRef]
- Blilou, I.; Xu, J.; Wildwater, M.; Willemsen, V.; Paponov, I.; Friml, J.; Heidstra, R.; Aida, M.; Palme, K.; Scheres, B. The PIN auxin efflux facilitator network controls growth and patterning in Arabidopsis roots. Nature 2005, 433, 39–44. [Google Scholar] [CrossRef]
- Zhu, Q.; Gao, P.; Liu, S.; Zhu, Z.C.; Amanullah, S.; Davis, A.R.; Luan, F. Comparative transcriptome analysis of two contrasting watermelon genotypes during fruit development and ripening. BMC Genome 2017, 18, 3. [Google Scholar] [CrossRef]
- Qian, X.; Ba, Y.; Zhuang, Q.F.; Zhong, G.F. RNA-seq technology and its application in fish transcriptomics. OMICS 2014, 18, 98–110. [Google Scholar] [CrossRef]
- Deyholos, M.K. Making the most of drought and salinity transcriptomics. Plant Cell Environ. 2010, 33, 648–654. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.C.; Chu, Y.G.; Liu, G.F.; Wang, M.H.; Jiang, J.; Hou, Y.J.; Qu, G.Z.; Yang, C.P. Identification of expressed sequence tags in an alkali grass (Puccinellia tenuiflora) cDNA library. J. Plant Physiol. 2007, 164, 78–89. [Google Scholar] [CrossRef] [PubMed]
- Postnikova, O.A.; Shao, J.; Nemchinov, L.G. Analysis of the alfalfa root transcriptome in response to salinity stress. Plant Cell Physiol. 2013, 54, 1041–1055. [Google Scholar] [CrossRef]
- Ruan, J.H.; Dean, A.K.; Zhang, W.X. A general co-expression networkbased approach to gene expression analysis: Comparison and applications. BMC Syst. Biol. 2010, 4, 8. [Google Scholar] [CrossRef]
- Perumal, D.; Leshchenko, V.V.; Kuo, P.Y.; Jiang, Z.W.; Readhead, B.; Eden, C.; Divakar, S.K.A.; Zhang, W.J.; Cho, H.J.; Chari, A.; et al. Weighted gene co-expression network analysis (WGCNA) identifies highly proliferative myeloma sub-group responsive to CDK4/ARK5. Blood 2014, 124, 3445. [Google Scholar] [CrossRef]
- Liu, J.J.; Wei, Z.; Li, J.H. Effects of copper on leaf membrane structure and root activity of maize seedling. Bot. Stud. 2014, 55, 47. [Google Scholar] [CrossRef]
- Li, W.X.; Yang, S.B.; Lu, Z.G.; He, Z.C.; Ye, Y.L.; Zhao, B.B.; Wang, L.; Jin, B. Cytological, physiological, and transcriptomic analyses of golden leaf coloration in Ginkgo biloba L. Hortic. Res. 2018, 5, 12. [Google Scholar] [CrossRef]
- Wu, S.Q.; Wang, Y.; Zhang, J.K.; Gong, X.J.; Zhang, Z.; Sun, J.J.; Chen, X.S.; Wang, Y.L. Exogenous melatonin improves physiological characteristics and promotes growth of strawberry seedlings under cadmium stress. Hortic. Plant J. 2021, 7, 13–22. [Google Scholar] [CrossRef]
- Cui, C.; Cai, J.; Zhang, S.X. Allelopathic effects of walnut (Juglans regia L.) rhizospheric soil extracts on germination and seedling growth of turnip (Brassica rapa L.). Allelopathy J. 2013, 32, 37–48. [Google Scholar]
- Narayanan, S.; Prasad, P.V.; Welti, R. Wheat leaf lipids during heat stress: II. Lipids experiencing coordinated metabolism are detected by analysis of lipid co-occurrence. Plant Cell Environ. 2016, 39, 608–617. [Google Scholar] [CrossRef] [PubMed]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattoori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2008, 36, D480–D484. [Google Scholar] [CrossRef]
- Kogelman, L.J.A.; Cirera, S.; Zhernakova, D.V.; Fredholm, M.; Franke, L.; Kadarmideen, H.N. Identification of co-expression gene networks, regulatory genes and pathways for obesity based on adipose tissue RNA Sequencing in a porcine model. BMC Med. Genom. 2014, 7, 57. [Google Scholar] [CrossRef]
- Wang, N.; Wang, R.; Wang, R.; Chen, S. Transcriptomics analysis revealing candidate networks and genes for the body size sexual dimorphism of Chinese tongue sole (Cynoglossus semilaevis). Funct. Integr. Genom. 2018, 18, 327–339. [Google Scholar] [CrossRef]
- Hu, W.; Yan, Y.; Hou, X.W.; He, Y.Z.; Wei, Y.X.; Yang, G.X.; He, G.Y.; Peng, M. TaPP2C1, a group F2 protein phosphatase 2C gene, confers resistance to salt stress in transgenic tobacco. PLoS ONE 2015, 10, e0129589. [Google Scholar] [CrossRef] [PubMed]
- Kwon, T.; Sparks, J.A.; Nakashima, J.; Allen, S.A.; Tang, Y.; Blancaflor, E.B. Transcriptional response of Arabidopsis seedlings during spaceflight reveals peroxidase and cell wall remodeling genes associated with root hair development. Am. J. Bot. 2015, 102, 21–35. [Google Scholar] [CrossRef] [PubMed]
- Ishida, K.; Yokoyama, R. Reconsidering the function of the xyloglucan endotransglucosylase/hydrolase family. J. Plant Res. 2022, 135, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Bao, F.; Li, J.Y. Evidence that the auxin signaling pathway interacts with plant stress response. Acta Bot. Sin. 2002, 44, 532–536. (In Chinese) [Google Scholar]
- de Kruijff, B. Lipid polymorphism and biomembrane function. Curr. Opin. Chem. Biol. 1997, 1, 564–569. [Google Scholar] [CrossRef]
- Frolov, V.A.; Shnyrova, A.V.; Zimmerberg, J. Lipid polymorphisms and membrane shape. Cold Spring Harb. Perspect. Biol. 2021, 3, a004747. [Google Scholar] [CrossRef]
- Yu, L.H.; Zhou, C.; Fan, J.L.; Shanklin, J.; Xu, C.C. Mechanisms and functions of membrane lipid remodeling in plants. Plant J. 2009, 107, 37–53. [Google Scholar] [CrossRef]
- Benning, C. Mechanisms of lipid transport involved in organelle biogenesis in plant cells. Annu. Rev. Cell Dev. Biol. 2009, 25, 71–91. [Google Scholar] [CrossRef]
- Hori, K.; Nobusawa, T.; Watanabe, T.; Madoka, Y.; Suzuki, H.; Shibata, D.; Shimojima, M.; Ohta, H. Tangled evolutionary processes with commonality and diversity in plastidial glycolipid synthesis in photosynthetic organisms. Biochim. Biophys. Acta 2016, 1861, 1294–1308. [Google Scholar] [CrossRef]
- Demé, B.; Cataye, C.; Block, M.A.; Maréchal, E.; Jouhet, J. Contribution of galactoglycerolipids to the 3-dimensional architecture of thylakoids. FASEB J. 2014, 28, 3373–3383. [Google Scholar] [CrossRef]
- Nunes-Nesi, A.; Nascimento Vde, L.; de Oliveira Silva, F.M.; Zsogon, A.; Araujo, W.L.; Sulpice, R. Natural genetic variation for morphological and molecular determinants of plant growth and yield. J. Exp. Bot. 2016, 67, 2989–3001. [Google Scholar] [CrossRef]
- Rasouli, F.; Kiani-Pouya, A.; Tahir, A.; Shabala, L.; Chen, Z.; Shabala, S. A comparative analysis of stomatal traits and photosynthetic responses in closely related halophytic and glycophytic species under saline conditions. Environ. Exp. Bot. 2021, 181, 104300. [Google Scholar] [CrossRef]
- Chutipaijit, S.; Cha-um, S.; Sompornpailin, K. High contents of proline and anthocyanin increase protective response to salinity in Oryza sativa L. spp. Indica. Aust. J. Crop Sci. 2011, 5, 1191–1198. [Google Scholar]
- Haque, M.I.; Siddiqui, S.A.; Jha, B.; Rathore, M.S. Interactive effects of abiotic stress and elevated CO2 on physio-chemical and photosynthetic responses in Suaeda species. J. Plant Growth Reg. 2021, 41, 2930–2948. [Google Scholar] [CrossRef]
- Khatri, K.; Rathore, M.S. Photosystem photochemistry, prompt and delayed fluorescence, photosynthetic responses and electron flow in tobacco under drought and salt stress. Photosynthetica 2019, 57, 61–74. [Google Scholar] [CrossRef]
- Santos, C.V. Regulation of chlorophyll biosynthesis and degradation by salt stress in sunflower leaves. Sci. Hortic. 2004, 103, 93–99. [Google Scholar] [CrossRef]
- Li, J.M.; Hu, L.P.; Zhang, L.; Pan, X.B.; Hu, X.H. Exogenous spermidine is enhancing tomato tolerance to salinity–alkalinity stress by regulating chloroplast antioxidant system and chlorophyll metabolism. BMC Plant Biol. 2015, 15, 303. [Google Scholar] [CrossRef]
- Tanka, R.; Kobayashi, K.; Masuda, T. Tetrapyrrole metabolism in Arabidopsis thaliana. Arab. Book 2011, 9, e0145. [Google Scholar] [CrossRef]
- Verdecia, M.A.; Larkin, R.M.; Ferrer, J.L.; Riek, R.; Chory, J.; Noel, J.P. Structure of the Mg-Chelatase Cofactor GUN4 Reveals a Novel Hand-Shaped Fold for Porphyrin Binding. PLoS Biol. 2005, 3, e151. [Google Scholar] [CrossRef]
- Sairam, R.; Tyagi, A. Physiology and molecular biology of salinity stress tolerance in plants. Curr. Sci. 2004, 86, 407–421. [Google Scholar]
- Chaves, M.; Costa, J.; Saibo, N. Recent advances in photosynthesis under drought and salinity. Adv. Bot. Res. 2011, 57, 49–104. [Google Scholar] [CrossRef]
- Wang, R.L.; Hua, C.; Zhou, F.; Zhou, Q.C. Effects of NaCl stress on photochemical activity and thylakoid membrane polypeptide composition of a salt-tolerant and a saltsensitive rice cultivar. Photosynthetica 2009, 47, 125–127. [Google Scholar] [CrossRef]
- Arena, C.; Figlioli, F.; Sorrentino, M.C.; Izzo, L.G.; Capozzi, F.; Giordano, S.; Spagnuolo, V. Ultrastructural, protein and photosynthetic alterations induced by Pb and Cd in Cynara cardunculus L., and its potential for phytoremediation. Ecotoxicol. Environ. Saf. 2017, 145, 83–89. [Google Scholar] [CrossRef]
- Bajji, M.; Kinet, J.M.; Lutts, S. The use of the electrolyte leakage method for assessing cell membrane stability as a water stress tolerance test in durum wheat. Plant Growth Regul. 2002, 36, 61–70. [Google Scholar] [CrossRef]
- Restrepo, H.; Gómez, M.I.; Garzón, A.; Manrique, L.; Alzate, F.; López, J.; Rodríguez, A. Respuesta bioquímica de plántulas de maíz (Zea mays L.) a diferentes condiciones de temperatureas nocturnas. Rev. Colomb. Cienc. Hortíc. 2013, 7, 252–262. [Google Scholar] [CrossRef]
- Kurusu, T.; Kuchisu, K.; Tada, Y. Plant signaling networks involving Ca2+ and Rboh/Nox-mediated ROS production under salinity stress. Front. Plant Sci. 2015, 6, 427. [Google Scholar] [CrossRef]
- Wolf, S.; Hematy, K.; Hofte, H. Growth control and cell wall signaling in plants. Annu. Rev. Plant Biol. 2012, 63, 381–407. [Google Scholar] [CrossRef] [PubMed]
- Neumann, P.M.; Azaizeh, H.; Leon, D. Hardening of root cell walls: A growth inhibitory response to salinity stress. Plant Cell Environ. 1994, 17, 303–309. [Google Scholar] [CrossRef]
- Fry, S.C.; Willis, S.C.; Paterson, A.E.J. Intraprotoplasmic and wall localised formation of arabinoxylan-bound diferulates and larger ferulate coupling-products in maize cell-suspension cultures. Planta 2000, 211, 679–692. [Google Scholar] [CrossRef]
- Jung, J.H.; Park, C.M. Auxin modulation of salt stress signaling in Arabidopsis seed germination. Plant Signal. Behav. 2014, 6, 1198–1200. [Google Scholar] [CrossRef] [PubMed]
- Schopfer, P.; Liszkay, A.; Bechtold, M.; Frahry, G.; Wagner, A. Evidence that hydroxyl radicals mediate auxin-induced extension growth. Planta 2002, 214, 821–828. [Google Scholar] [CrossRef] [PubMed]
- Lavenus, J.; Goh, T.; Roberts, I.; Guyomarch, S.; Lucas, M.; Smet, I.D.; Fukaki, H.; Beekman, T.; Bennett, M.; Llaplaze, L. Lateral root development in Arabidopsis: Fifty shades of auxin. Trends Plant Sci. 2013, 18, 450–458. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene Annotation |
|---|---|
| MELO3C006053.2 | Protein CURVATURE THYLAKOID 1A, chloroplastic |
| MELO3C004491.2 | Thylakoid lumenal 16.5 kDa protein, chloroplastic |
| MELO3C025475.2 | psbP-like protein 1, chloroplastic |
| MELO3C023596.2 | Photosystem II stability/assembly factor HCF136, chloroplastic |
| MELO3C010708.2 | photosystem II core complex proteins psbY, chloroplastic |
| MELO3C004214.2 | Chlorophyll a-b binding protein, chloroplastic |
| MELO3C022113.2 | PS II-associated light-harvesting complex II |
| MELO3C015536.2 | photosystem II D2 protein-like |
| MELO3C006606.2 | Divinyl chlorophyllide a 8-vinyl-reductase, chloroplastic |
| MELO3C017176.2 | chlorophyll biosynthetic process |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, T.; Amanullah, S.; Xu, H.; Gao, P.; Du, Z.; Hu, X.; Han, M.; Che, Y.; Zhang, L.; Qi, G.; et al. RNA-Seq Identified Putative Genes Conferring Photosynthesis and Root Development of Melon under Salt Stress. Genes 2023, 14, 1728. https://doi.org/10.3390/genes14091728
Liu T, Amanullah S, Xu H, Gao P, Du Z, Hu X, Han M, Che Y, Zhang L, Qi G, et al. RNA-Seq Identified Putative Genes Conferring Photosynthesis and Root Development of Melon under Salt Stress. Genes. 2023; 14(9):1728. https://doi.org/10.3390/genes14091728
Chicago/Turabian StyleLiu, Tai, Sikandar Amanullah, Huichun Xu, Peng Gao, Zhiqiang Du, Xixi Hu, Mo Han, Ye Che, Ling Zhang, Guochao Qi, and et al. 2023. "RNA-Seq Identified Putative Genes Conferring Photosynthesis and Root Development of Melon under Salt Stress" Genes 14, no. 9: 1728. https://doi.org/10.3390/genes14091728
APA StyleLiu, T., Amanullah, S., Xu, H., Gao, P., Du, Z., Hu, X., Han, M., Che, Y., Zhang, L., Qi, G., & Wang, D. (2023). RNA-Seq Identified Putative Genes Conferring Photosynthesis and Root Development of Melon under Salt Stress. Genes, 14(9), 1728. https://doi.org/10.3390/genes14091728