
Hsp70 Gene Family in Sebastiscus marmoratus: The Genome-Wide Identification and Transcriptome Analysis under Thermal Stress

Abstract
:1. Introduction
2. Results
2.1. The Hsp70 Genes in S. marmoratus Based on Genome Data Analysis
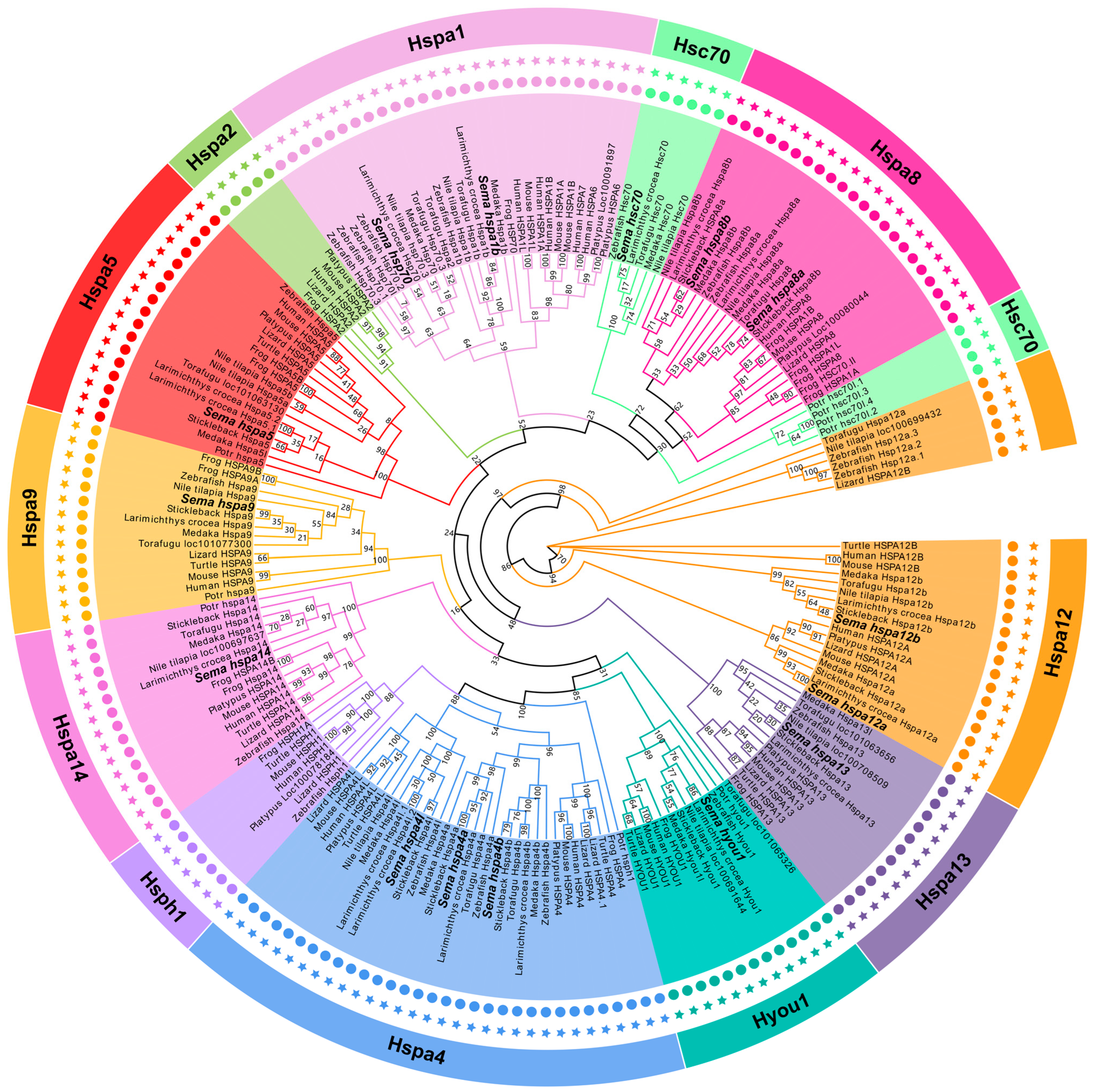
2.2. The Phylogeny of the Hsp70 Genes among Species
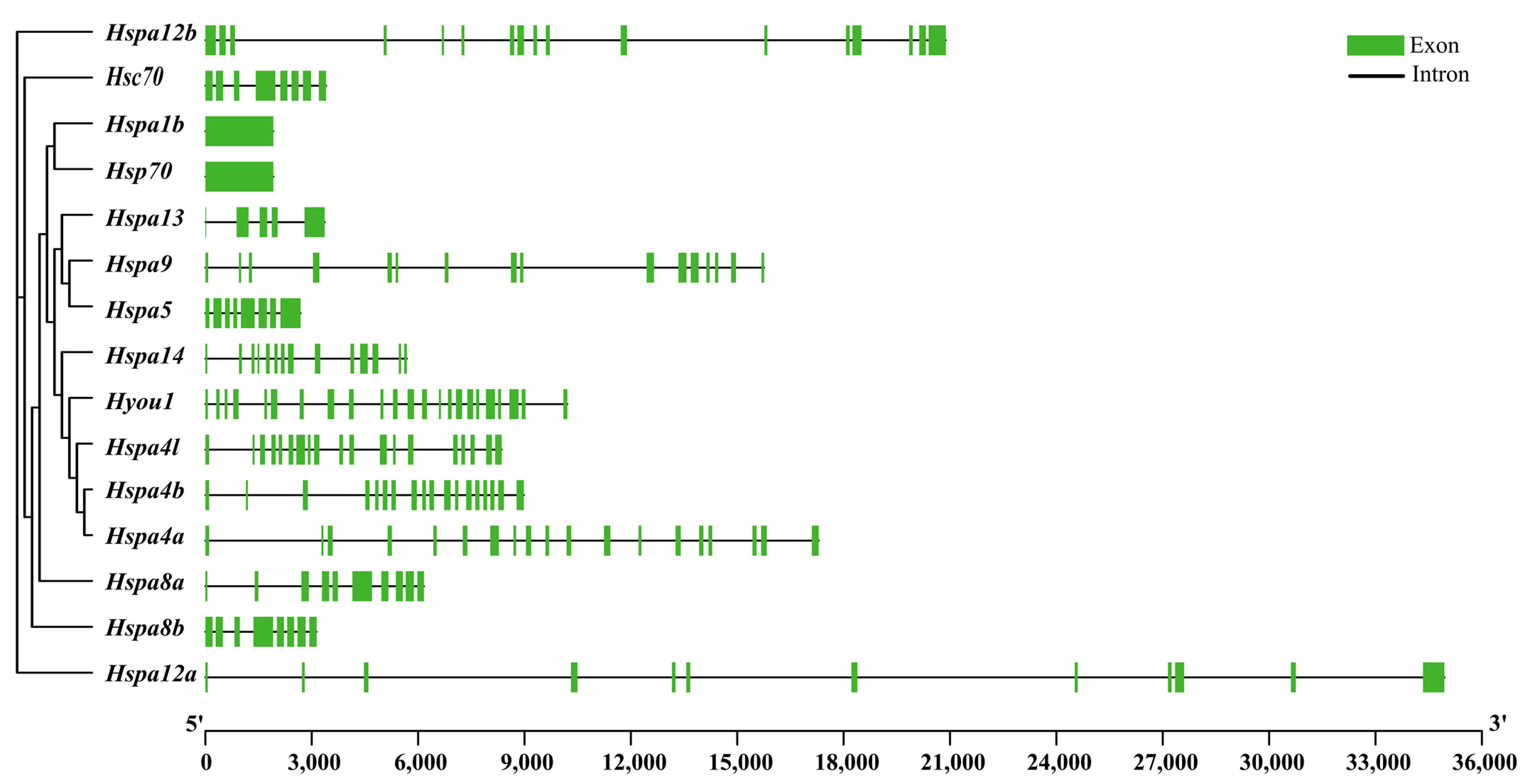
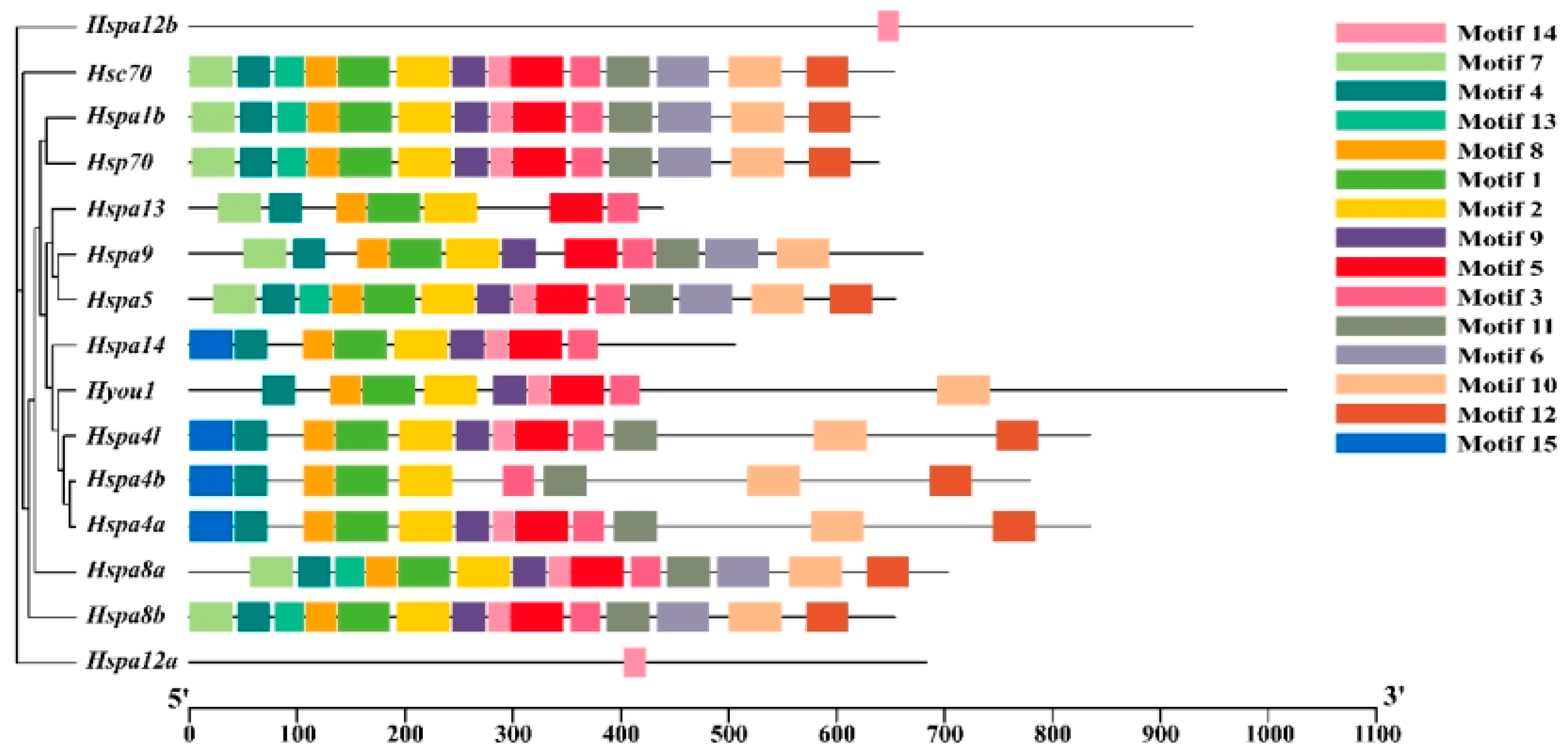
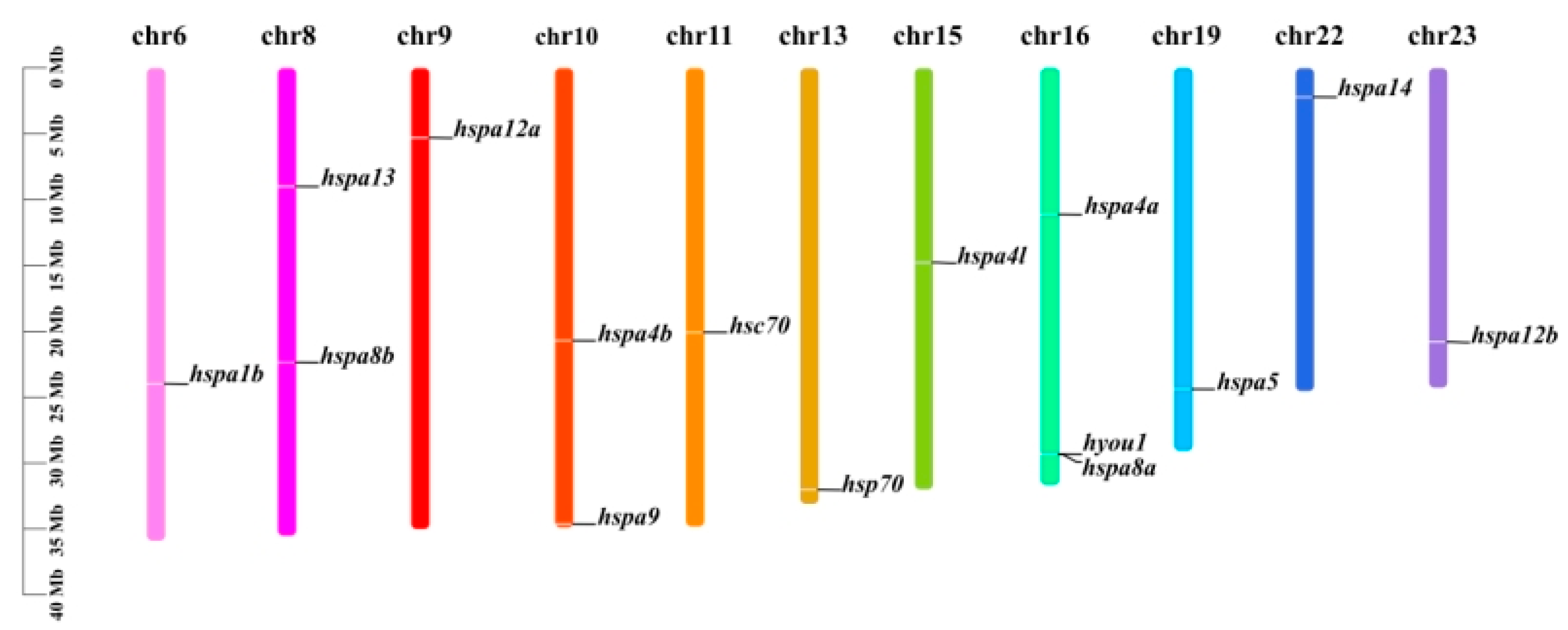
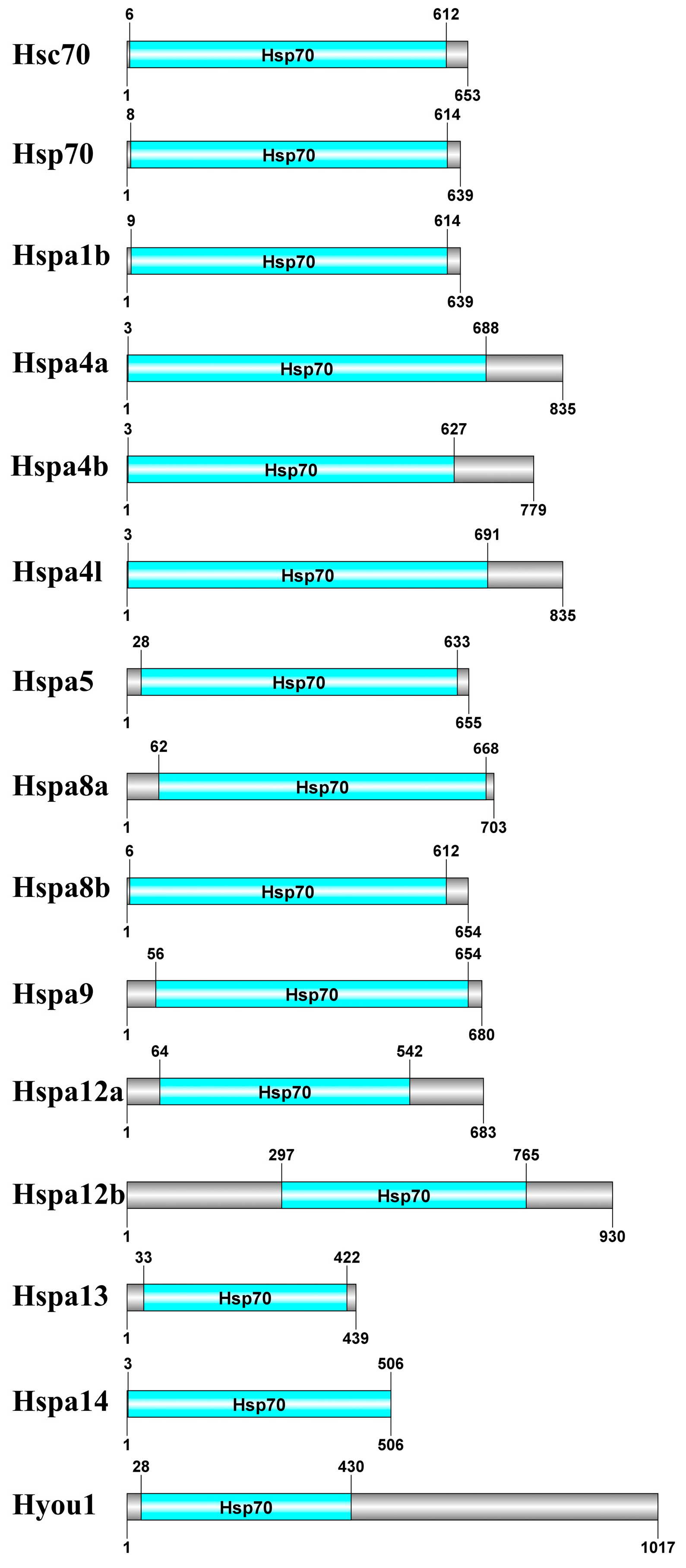
2.3. Gene Structure, Motif, and Chromosomal Location Analysis
2.4. Protein Signal Peptides Predictive Analysis
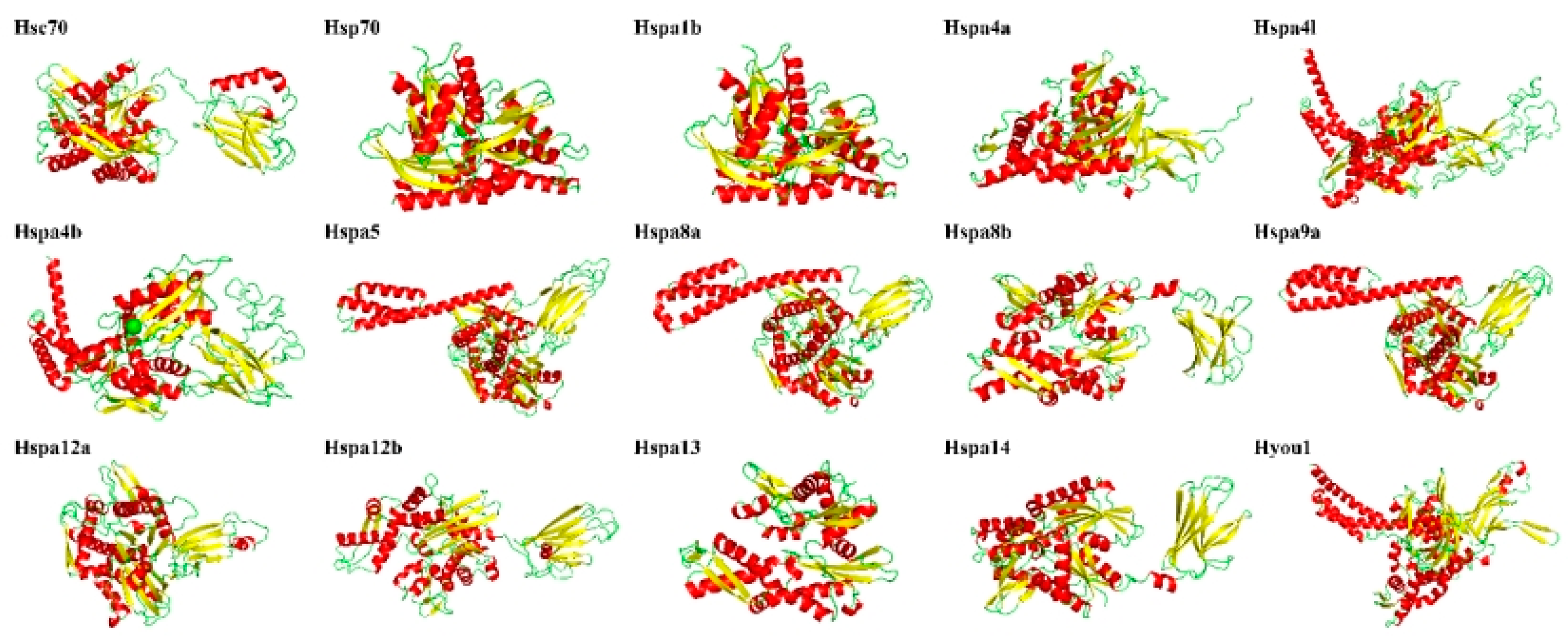
2.5. Secondary and Third Structure Prediction, Subcellular Localization of Hsp70 Proteins
2.6. Selection Test on Duplicated Hsp70 Gene Pairs
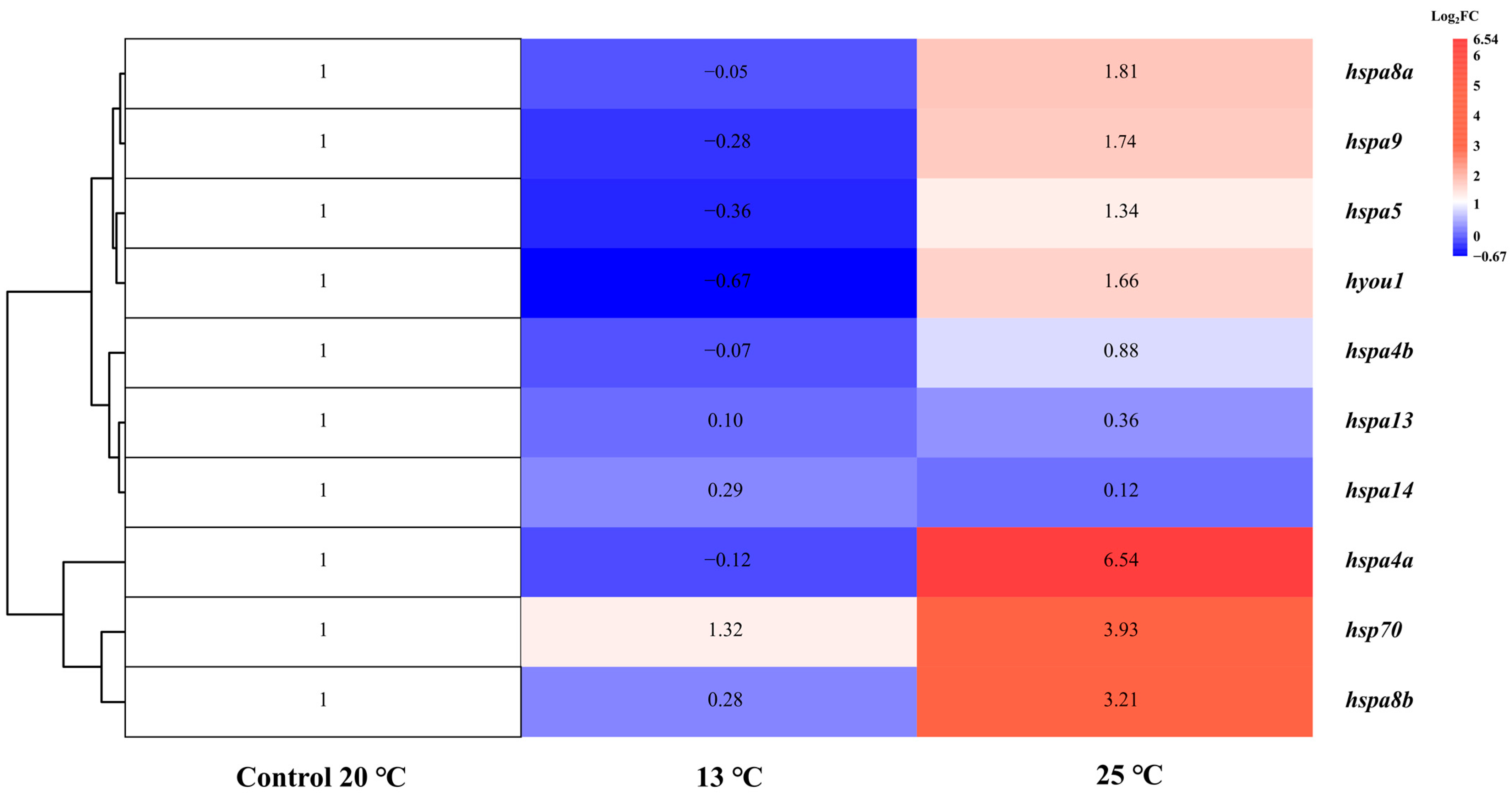
2.7. Expression Profiles of Hsp70 Genes under Thermal Stress Treatment
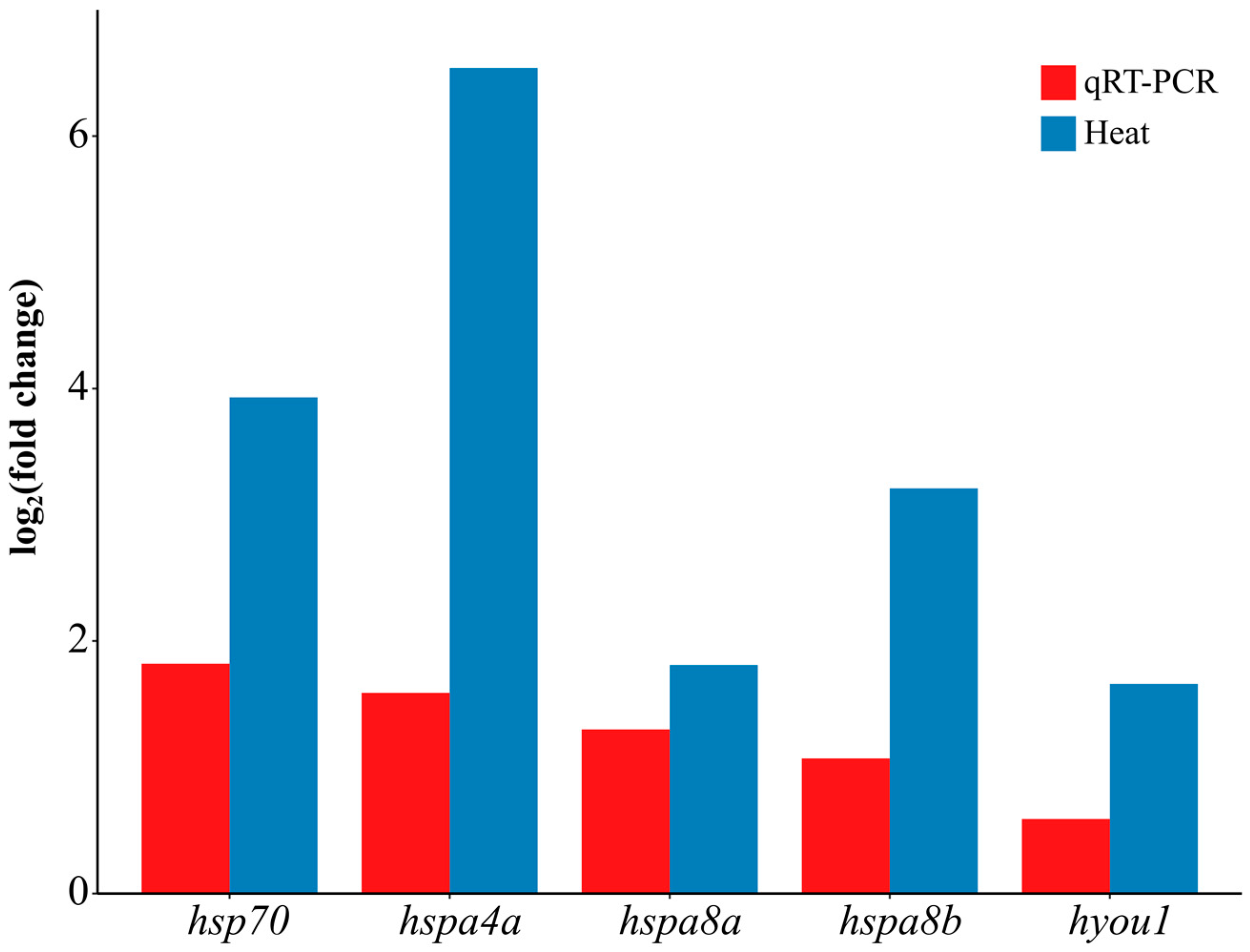
2.8. Validation of Transcriptomic Data via qRT-PCR
3. Discussion
4. Materials and Methods
4.1. Animal Materials and Experiment
4.2. Identification of Hsp70 Members
4.3. Phylogenetic Relationship Analysis
4.4. Gene Structure, Motif, Chromosomal Location, and Conserved Domain Analysis
4.5. Three-Dimensional Structure Analysis of Hsp70 Proteins
4.6. Predictive Analysis of Protein Signal Peptides and Selection Test of Hsp70s
4.7. Expression Pattern Analysis of Hsp70 Genes in S. marmoratus
4.8. Primer Design, RNA Isolation, and Quantitative Real-Time PCR (qRT-PCR) Validation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gestin, M.; Falato, L.; Ciccarelli, M.; Cerrato, C.P.; Andréasson, C.; Langel, Ü. Transfection of Heat Shock Protein 70 kDa (HSP70). Int. J. Pept. Res. Ther. 2022, 28, 105. [Google Scholar] [CrossRef]
- Grewal, H.S.; Yoshinaga, T.; Ehsan, H.; Yu, E.; Kaneko, G. A genome-wide screening of the 70 kDa heat shock protein (HSP70) genes in the rotifer Brachionus plicatilis sensu stricto with a characterization of two heat-inducible HSP70 genes. Cell Stress Chaperones 2022, 28, 593–594. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Zhang, J.; Li, C.; Yao, J.; Jiang, C.; Li, Y.; Liu, S.; Liu, Z. Genome-wide identification of hsp40 genes in channel catfish and their regulated expression after bacterial infection. PLoS ONE 2014, 9, e115752. [Google Scholar] [CrossRef] [PubMed]
- Feder, M.E.; Hofmann, G.E. Heat-shock proteins, molecular chaperones, and the stress response: Evolutionary and ecological physiology. Annu. Rev. Physiol. 1999, 61, 243–282. [Google Scholar] [CrossRef]
- Fan, K.P.; Hua, X.T.; Liu, Y.F.; Zhang, Z.Q.; Li, X.H.; Liu, Y.; Liu, P.F. HSP70 gene expression responses to the temperature stress in pufferfish (Takifugu rubripes). Biosci. Biotechnol. Biochem. 2021, 85, 1088–1096. [Google Scholar] [CrossRef]
- Chen, B.; Feder, M.E.; Kang, L. Evolution of heat-shock protein expression underlying adaptive responses to environmental stress. Mol. Ecol. 2018, 27, 3040–3054. [Google Scholar] [CrossRef]
- Wang, F.; Li, D.; Chen, Q.; Ma, L. Genome-wide survey and characterization of the small heat shock protein gene family in Bursaphelenchus xylophilus. Gene 2016, 579, 153–161. [Google Scholar] [CrossRef]
- Song, L.; Li, C.; Xie, Y.; Liu, S.; Zhang, J.; Yao, J.; Jiang, C.; Li, Y.; Liu, Z. Genome-wide identification of Hsp70 genes in channel catfish and their regulated expression after bacterial infection. Fish Shellfish Immunol. 2016, 49, 154–162. [Google Scholar] [CrossRef]
- Liu, J.; Wang, R.; Liu, W.; Zhang, H.; Guo, Y.; Wen, R. Genome-Wide Characterization of Heat-Shock Protein 70s from Chenopodium quinoa and Expression Analyses of Cqhsp70s in Response to Drought Stress. Genes 2018, 9, 35. [Google Scholar] [CrossRef]
- Karlin, S.; Brocchieri, L. Heat shock protein 70 family: Multiple sequence comparisons, function, and evolution. J. Mol. Evol. 1998, 47, 565–577. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, X.; Shi, H.; Song, N.; Han, F.; Chai, X.; Liu, Q.; Wang, Y.; Gao, T. Comparative transcriptomic analysis of the liver and spleen in marbled rockfish (Sebastiscus marmoratus) challenged with polyriboinosinic polyribocytidylic acid (poly(I:C)). Aquaculture 2022, 554, 738144. [Google Scholar] [CrossRef]
- Xu, S.-Y.; Sun, D.-R.; Song, N.; Gao, T.-X.; Han, Z.-Q.; Shui, B.-N. Local adaptation shapes pattern of mitochondrial population structure in Sebastiscus marmoratus. Environ. Biol. Fishes 2017, 100, 763–774. [Google Scholar] [CrossRef]
- Kita, J.; Tsuchida, S.; Setoguma, T. Temperature preference and tolerance, and oxygen consumption of the marbled rockfish, Sebastiscus marmoratus. Mar. Biol. 1996, 125, 467–471. [Google Scholar] [CrossRef]
- Yin, F.; Gong, Q.; Li, Y.; Dan, X.; Sun, P.; Gao, Q.; Shi, Z.; Peng, S.; Li, A. Effects of Cryptocaryon irritans infection on the survival, feeding, respiratory rate and ionic regulation of the marbled rockfish Sebastiscus marmoratus. Parasitology 2014, 141, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zhang, X.; Li, C.; Zhang, H.; Yanagimoto, T.; Na, S.; Gao, T. Population genetic structure of Marbled Rockfish, Sebastiscus marmoratus (Cuvier, 1829), in the northwestern Pacific Ocean. ZooKeys 2019, 830, 127–144. [Google Scholar] [CrossRef] [PubMed]
- Yin, F.; Qian, D. Transcriptomic analysis reveals the key immune-related signalling pathways of Sebastiscus marmoratus in response to infection with the parasitic ciliate Cryptocaryon irritans. Parasites Vectors 2017, 10, 576. [Google Scholar] [CrossRef]
- Peng, S.; Yue, Y.; Gao, Q.; Shi, Z.; Yin, F.; Wang, J. Influence of dietary n-3 LC-PUFA on growth, nutritional composition and immune function in marine fish Sebastiscus marmoratus. Chin. J. Oceanol. Limnol. 2014, 32, 1000–1008. [Google Scholar] [CrossRef]
- Wu, C.J.O.Z.O.U. The Effect of Several Environmental Factors on the Survival Rate of Sebasticus marmoratus Larvae. J. Zhejiang Ocean. Univ. 2000, 19, 12–16. [Google Scholar]
- Xu, M. Preliminry Study on The Fishrdes Biology of Sebastiscus marmoratus. Mar. Tieheries 1999, 4, 159–162. [Google Scholar]
- Giorno, F.; Wolters-Arts, M.; Grillo, S.; Scharf, K.D.; Vriezen, W.H.; Mariani, C. Developmental and heat stress-regulated expression of HsfA2 and small heat shock proteins in tomato anthers. J. Exp. Bot. 2010, 61, 453–462. [Google Scholar] [CrossRef]
- Zhao, P.; Wang, D.; Wang, R.; Kong, N.; Zhang, C.; Yang, C.; Wu, W.; Ma, H.; Chen, Q. Genome-wide analysis of the potato Hsp20 gene family: Identification, genomic organization and expression profiles in response to heat stress. BMC Genom. 2018, 19, 61. [Google Scholar] [CrossRef]
- Lou, F.; Han, Z. Full-length transcripts facilitates Portunus trituberculatus genome structure annotation. J. Oceanol. Limnol. 2022, 40, 2042–2051. [Google Scholar] [CrossRef]
- Han, Z.Q.; Gao, T.X.; Lou, F.R.; Liu, Q. Charybdis japonica genome provides insights into desiccation adaptation and sex-determining region. Zool. Res. 2022, 43, 927–930. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, M.; Chen, J.; Rong, J.; Ding, M. Genome-wide analysis of HSP70 superfamily in Gossypium raimondii and the expression of orthologs in Gossypium hirsutum. Yi Chuan = Hereditas 2014, 36, 921–933. [Google Scholar]
- Wang, J.; Chen, J.; Hu, Y.; Ying, S.H.; Feng, M.G. Roles of six Hsp70 genes in virulence, cell wall integrity, antioxidant activity and multiple stress tolerance of Beauveria bassiana. Fungal Genet. Biol. FG B 2020, 144, 103437. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Shi, F.; Li, Y.; Zong, S.; Tao, J. Genome-wide identification and expression analysis of the Hsp gene superfamily in Asian long-horned beetle (Anoplophora glabripennis). Int. J. Biol. Macromol. 2022, 200, 583–592. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Deng, Z.; Xu, S.; Zhang, H.; Han, Z. Genome-wide identification and low-salinity stress analysis of the Hsp70 gene family in swimming crab (Portunus trituberculatus). Int. J. Biol. Macromol. 2022, 208, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Brocchieri, L.; Conway de Macario, E.; Macario, A.J. hsp70 genes in the human genome: Conservation and differentiation patterns predict a wide array of overlapping and specialized functions. BMC Evol. Biol. 2008, 8, 19. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.D.; Xu, H.X.; Han, Z.Q. Genome-Wide Identification of Hsp70 Genes in the Large Yellow Croaker (Larimichthys crocea) and Their Regulated Expression Under Cold and Heat Stress. Genes 2018, 9, 590. [Google Scholar] [CrossRef]
- Deng, Z.; Sun, S.; Gao, T.; Han, Z. The Hsp70 Gene Family in Boleophthalmus pectinirostris: Genome-Wide Identification and Expression Analysis under High Ammonia Stress. Animals 2019, 9, 36. [Google Scholar] [CrossRef]
- Dehal, P.; Boore, J.L. Two rounds of whole genome duplication in the ancestral vertebrate. PLoS Biol. 2005, 3, e314. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Komatsu, S. Plant subcellular proteomics: Application for exploring optimal cell function in soybean. J. Proteom. 2016, 143, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Sung, D.Y.; Vierling, E.; Guy, C.L. Comprehensive expression profile analysis of the Arabidopsis Hsp70 gene family. Plant Physiol. 2001, 126, 789–800. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.H.; Gho, H.J.; Nguyen, M.X.; Kim, S.R.; An, G. Genome-wide expression analysis of HSP70 family genes in rice and identification of a cytosolic HSP70 gene highly induced under heat stress. Funct. Integr. Genom. 2013, 13, 391–402. [Google Scholar] [CrossRef]
- Jungkunz, I.; Link, K.; Vogel, F.; Voll, L.M.; Sonnewald, S.; Sonnewald, U. AtHsp70-15-deficient Arabidopsis plants are characterized by reduced growth, a constitutive cytosolic protein response and enhanced resistance to TuMV. Plant J. 2011, 66, 983–995. [Google Scholar] [CrossRef]
- Freeling, M. Bias in plant gene content following different sorts of duplication: Tandem, whole-genome, segmental, or by transposition. Annu. Rev. Plant Biol. 2009, 60, 433–453. [Google Scholar] [CrossRef]
- Holland, P.W.; Marlétaz, F.; Maeso, I.; Dunwell, T.L.; Paps, J. New genes from old: Asymmetric divergence of gene duplicates and the evolution of development. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2017, 372, 20150480. [Google Scholar] [CrossRef]
- Conant, G.C.; Wolfe, K.H. Turning a hobby into a job: How duplicated genes find new functions. Nat. Rev. Genet. 2008, 9, 938–950. [Google Scholar] [CrossRef]
- Voldoire, E.; Brunet, F.; Naville, M.; Volff, J.N.; Galiana, D. Expansion by whole genome duplication and evolution of the sox gene family in teleost fish. PLoS ONE 2017, 12, e0180936. [Google Scholar] [CrossRef]
- Giri, S.S.; Sen, S.S.; Sukumaran, V. Role of HSP70 in cytoplasm protection against thermal stress in rohu, Labeo rohita. Fish Shellfish Immunol. 2014, 41, 294–299. [Google Scholar] [CrossRef]
- Das, T.; Pal, A.; Chakraborty, S.; Saydmohammed, M.; Chatterjee, N.; Apte, S. Metabolic Elasticity and Induction of Heat Shock Protein 70 in Labeo rohita Acclimated to Three Temperatures. Asian-Australas. J. Anim. Sci. 2006, 19, 1033–1039. [Google Scholar] [CrossRef]
- Sonna, L.A.; Fujita, J.; Gaffin, S.L.; Lilly, C.M. Invited review: Effects of heat and cold stress on mammalian gene expression. J. Appl. Physiol. 2002, 92, 1725–1742. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela Castillo, A.; Sánchez-Paz, A.; Castro-Longoria, R.; López-Torres, M.; Grijalva-Chon, J.M. Hsp70 function and polymorphism, implications for mollusk aquaculture Hsp70 function and polymorphism, its implications for mollusk aquaculture: A review. Lat. Am. J. Aquat. Res. 2019, 47, 224–231. [Google Scholar] [CrossRef]
- Ma, F.; Luo, L. Genome-wide identification of Hsp70/110 genes in rainbow trout and their regulated expression in response to heat stress. PeerJ 2020, 8, e10022. [Google Scholar] [CrossRef] [PubMed]
- Eddy, S.R. Profile hidden Markov models. Bioinformatics 1998, 14, 755–763. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39, W29–W37. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef]
- Geourjon, C.; Deléage, G. SOPMA: Significant improvements in protein secondary structure prediction by consensus prediction from multiple alignments. Comput. Appl. Biosci. 1995, 11, 681–684. [Google Scholar] [CrossRef]
- Horton, P.; Park, K.J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C.J.; Nakai, K. WoLF PSORT: Protein localization predictor. Nucleic Acids Res. 2007, 35, W585–W587. [Google Scholar] [CrossRef]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Ren, J.; Wen, L.; Gao, X.; Jin, C.; Xue, Y.; Yao, X. DOG 1.0: Illustrator of protein domain structures. Cell Res. 2009, 19, 271–273. [Google Scholar] [CrossRef]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef]
- Colovos, C.; Yeates, T.O. Verification of protein structures: Patterns of nonbonded atomic interactions. Protein. Sci. 1993, 2, 1511–1519. [Google Scholar] [CrossRef]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef]
- Wang, D.; Zhang, Y.; Zhang, Z.; Zhu, J.; Yu, J. KaKs_Calculator 2.0: A toolkit incorporating gamma-series methods and sliding window strategies. Genom. Proteom. Bioinform. 2010, 8, 77–80. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Gene Name | Gene ID | CDS * Length (bp) | Protein Length (aa) | Hsp70 Domain Location (aa) | Molecular Weight (kDa) | Theoretic al pI * | Chromosome | Location |
|---|---|---|---|---|---|---|---|---|---|
| 1 | hsc70 | Seb016770 | 1962 | 653 | 6-612 | 71,379.77 | 5.19 | chr11 | 20077654:20081059 |
| 2 | hsp70 | Seb019324 | 1920 | 639 | 8-614 | 70,157.35 | 5.41 | chr13 | 32013287:32015206 |
| 3 | hspa1b | Seb001220 | 1920 | 639 | 9-614 | 70,308.53 | 5.44 | chr6 | 23970762:23972681 |
| 4 | hspa4a | Seb020794 | 2508 | 835 | 3-688 | 93,851.78 | 5.61 | chr16 | 11119016:11136314 |
| 5 | hspa4b | Seb006269 | 2340 | 779 | 3-627 | 87,193.47 | 5.06 | chr10 | 20703166:20712148 |
| 6 | hspa4l | Seb001425 | 2508 | 835 | 3-691 | 93,694.10 | 5.50 | chr15 | 14780364:14788721 |
| 7 | hspa5 | Seb006799 | 1968 | 655 | 28-633 | 72,285.76 | 4.98 | chr19 | 24390720:24393407 |
| 8 | hspa8a | Seb009609 | 2112 | 703 | 62-668 | 76,830.31 | 5.47 | chr16 | 29350778:29356943 |
| 9 | hspa8b | Seb013994 | 1965 | 654 | 6-612 | 71,622.94 | 5.24 | chr8 | 22354391:22357531 |
| 10 | hspa9 | Seb014614 | 2043 | 680 | 56-654 | 73,556.67 | 6.24 | chr10 | 34615309:34631069 |
| 11 | hspa12a | Seb017726 | 2052 | 683 | 64-542 | 76,345.16 | 8.73 | chr9 | 5309800:5344743 |
| 12 | hspa12b | Seb009063 | 2793 | 930 | 297-765 | 103,325.02 | 6.86 | chr23 | 20798860:20819742 |
| 13 | hspa13 | Seb000354 | 1320 | 439 | 33-422 | 48,108.26 | 5.35 | chr8 | 9021496:9024862 |
| 14 | hspa14 | Seb019773 | 1521 | 506 | 3-506 | 54,473.08 | 5.81 | chr22 | 2233197:2238880 |
| 15 | hyou1 | Seb009605 | 3054 | 1017 | 28-430 | 113,767.84 | 5.38 | chr16 | 29310839:29321048 |
| Protein | α Helix | β Turn | Random Coil | Extended Strand | Subcellular Location Prediction |
|---|---|---|---|---|---|
| Hsc70 | 41.65% | 7.04% | 33.38% | 17.92% | Cytoplasm |
| Hsp70 | 41.94% | 7.04% | 32.71% | 18.31% | Cytoplasm |
| Hspa1b | 42.41% | 6.73% | 31.77% | 19.09% | Cytoplasm |
| Hspa4a | 43.47% | 3.47% | 38.32% | 14.73% | Cytoplasm |
| Hspa4b | 40.56% | 3.34% | 40.95% | 15.15% | Cytoplasm |
| Hspa4l | 43.11% | 3.11% | 40.00% | 13.77% | Cytoplasm |
| Hspa5 | 43.66% | 7.18% | 30.23% | 18.93% | Endoplasmic reticulum |
| Hspa8a | 39.40% | 7.68% | 35.14% | 17.78% | Mitochondrion |
| Hspa8b | 42.66% | 7.03% | 32.42% | 17.89% | Cytoplasm |
| Hspa9 | 43.09% | 7.35% | 29.71% | 19.85% | Mitochondrion |
| Hspa12a | 30.89% | 4.39% | 44.66% | 20.06% | Nucleus |
| Hspa12b | 32.90% | 5.91% | 41.72% | 19.46% | Cytoplasm |
| Hspa13 | 40.32% | 7.74% | 31.66% | 20.27% | Plasma membrane |
| Hspa14 | 37.15% | 5.73% | 34.98% | 22.13% | Cytoplasm |
| Hyou1 | 47.89% | 3.44% | 36.28% | 12.39% | Endoplasmic reticulum |
| Gene Pair | Ka | Ks | Ka/Ks |
|---|---|---|---|
| hspa12a-hspa12b | 0.2870 | 3.9431 | 0.0728 |
| hspa4a-hspa4b | 0.1788 | 4.2046 | 0.0425 |
| hspa4a-hspa4l | 0.2649 | 3.9346 | 0.0673 |
| hspa4b-hspa4l | 0.2861 | 3.7453 | 0.0764 |
| hspa8a-hspa8b | 0.0322 | 1.8155 | 0.0177 |
| Gene Name | log2FC | ||
|---|---|---|---|
| Control Group | Low-Temperature | High-Temperature | |
| hsp70 | 1 | 1.32 | 3.93 |
| hspa4a | 1 | −0.12 | 6.54 |
| hspa4b | 1 | −0.07 | 0.88 |
| hspa5 | 1 | −0.36 | 1.34 |
| hspa8a | 1 | −0.05 | 1.81 |
| hspa8b | 1 | 0.28 | 3.21 |
| hspa9 | 1 | −0.28 | 1.74 |
| hspa13 | 1 | 0.10 | 0.36 |
| hspa14 | 1 | 0.29 | 0.12 |
| hyou1 | 1 | −0.67 | 1.66 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, X.; Jin, S.; Shou, C.; Han, Z. Hsp70 Gene Family in Sebastiscus marmoratus: The Genome-Wide Identification and Transcriptome Analysis under Thermal Stress. Genes 2023, 14, 1779. https://doi.org/10.3390/genes14091779
Han X, Jin S, Shou C, Han Z. Hsp70 Gene Family in Sebastiscus marmoratus: The Genome-Wide Identification and Transcriptome Analysis under Thermal Stress. Genes. 2023; 14(9):1779. https://doi.org/10.3390/genes14091779
Chicago/Turabian StyleHan, Xiaolu, Shihuai Jin, Chenyan Shou, and Zhiqiang Han. 2023. "Hsp70 Gene Family in Sebastiscus marmoratus: The Genome-Wide Identification and Transcriptome Analysis under Thermal Stress" Genes 14, no. 9: 1779. https://doi.org/10.3390/genes14091779
APA StyleHan, X., Jin, S., Shou, C., & Han, Z. (2023). Hsp70 Gene Family in Sebastiscus marmoratus: The Genome-Wide Identification and Transcriptome Analysis under Thermal Stress. Genes, 14(9), 1779. https://doi.org/10.3390/genes14091779