

Transcriptome and Physiological Analysis Highlight Lignin Metabolism of the Fruit Dots Disordering during Postharvest Cold Storage in ‘Danxiahong’ Pear
 ,
,
Abstract
:1. Introduction
2. Results
2.1. ‘Danxiahong’ Pear Fruit Dots Morphology and Lignin Content
2.2. Scanning Electron Microscopy and Paraffin Section Observation
2.3. RNA Sequencing and Analysis of Differentially Expressed Genes
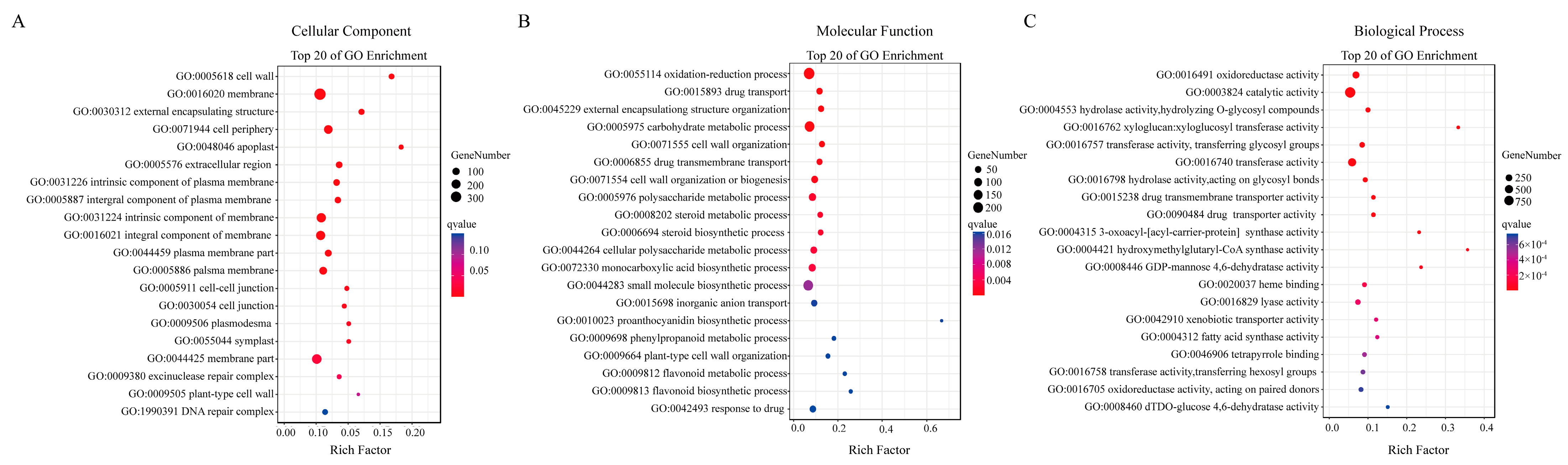
2.4. Gene Ontology (GO) Enrichment Analysis of DEGs
2.5. KEGG Enrichment Analysis of DEGs
2.6. Analysis of Lignin Biosynthesis Structural Genes
2.7. Analysis of Lignin Biosynthesis Regulatory Genes
2.8. RT-qPCR Analysis of the Lignin Biosynthesis Genes
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Experimental Treatments
4.2. Morphological Index Determination
4.3. Preparation and Observation of Paraffin Sections
4.4. Scanning Electron-Microscopy (SEM) Observation
4.5. Lignin Content Determination
4.6. Transcriptome Analysis with RNA-Seq Data Validation
4.7. DEG, GO, and KEGG Analysis
4.8. RNA Isolation and RT-qPCR Analysis
4.9. Principal Component Analysis
4.10. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wen, H.; Jiang, X.; Wang, W.Q.; Wu, M.Y.; Bai, H.J.; Wu, C.Y.; Shen, L. Comparative transcriptome analysis of candidate genes involved in chlorogenic acid biosynthesis during fruit development in three pear varieties of Xinjiang Uygur Autonomous Region. J. Zhejiang Univ. Sci. B 2022, 23, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Duan, R.W.; Zhang, X.Z.; Li, B.; Wang, M.R.; Xie, Y.R.; Li, P.; Wang, L.; Yang, J.; Xue, H.B. Comprehensive Evaluation of Fruit Spots in 296 Pear Germplasm Resources. Acta Hortic. Sin. 2023, in press. [Google Scholar]
- Tao, S.T.; Khanizadeh, S.; Zhang, H.; Zhang, S.L. Anatomy, ultrastructure and lignin distribution of stone cells in two Pyrus species. Plant Sci. 2009, 176, 413–419. [Google Scholar] [CrossRef]
- Wang, B.G.; Wang, Y.X.; Li, W.S.; Zhou, J.H.; Chang, H.; Golding, J.B. Effect of 1-MCP and ethylene absorbent on the development of lenticel disorder of ‘Xinli No.7′ pear and possible mechanisms. J. Sci. Food Agric. 2021, 101, 2525–2533. [Google Scholar] [CrossRef]
- Sub, M.; Dai, H.Y.; Li, P.H.; Wang, R.; Lin, Z.H. The development of fruit dots in pear ‘Chili’ and ‘Dangshan’. Deciduous Fruits 1983, 3–5. [Google Scholar]
- Wang, Y.L.; Zhang, X.F.; Wang RBai, Y.X.; Liu, C.L.; Yuan, Y.B.; Yang, Y.J.; Yang, S.L. Differential gene expression analysis of ‘Chili’ ( Pyrus bretschneideri ) fruit pericarp with two types of bagging treatments. Hortic. Res. 2017, 4, 17005. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Y.F.; Zhang, P.F.; Bian, Y.H.; Liu, Z.Y.; Zhang, C.; Liu, X.; Wang, C.L. An integrated metabolic and transcriptomic analysis reveals the mechanism through which fruit bagging alleviates exocarp semi-russeting in pear fruit. Tree Physiol. 2021, 41, 1306–1318. [Google Scholar] [CrossRef]
- Zhao, Q. Lignification: Flexibility, Biosynthesis and Regulation. Trends Plant Sci. 2016, 21, 713–721. [Google Scholar] [CrossRef]
- Guan, Y.Q.; Qin, X.L.; Pei, Y.; Li, X.L.; Feng, Y.X.; Wei, C.Q.; Cheng, Y.D.; Guan, J.F. Effect of Exogenous Chlorogenic Acid on Fruit Dot Formation and Expression of Related Genes in ‘Xuehua’ Pear (Pyrus bretschneideri). J. Agric. Biotechnol. 2021, 29, 258–267. [Google Scholar]
- Ali, M.M.; Anwar, R.; Yousef, A.F.; Li, B.; Luvisi, A.; Bellis, L.; Aprile, A.; Chen, F. Influence of Bagging on the Development and Quality of Fruits. Plants 2021, 10, 358. [Google Scholar] [CrossRef]
- Li, D.; Cheng, Y.D.; Guan, J.F. Effects of 1-methylcyclopropene on surface wax and related gene expression in cold-stored ‘Hongxiangsu’ pears. J. Sci. Food Agric. 2019, 99, 2438–2446. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.H.; Wang, X.Q.; Xu, J.F.; Zhang, Y.X.; Qi, B.X.; Jun, L. Dissecting the molecular mechanism of russeting in sand pear (Pyrus pyrifolia Nakai) by metabolomics, transcriptomics, and proteomics. Plant J. 2021, 108, 1644–1661. [Google Scholar] [CrossRef]
- Vanholme, R.B.; De Meester, B.; Ralph, J.; Boerjan, W. Lignin biosynthesis and its integrationin to metabolism. Curr. Opin. Biotechnol. 2019, 56, 230–239. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.K.; Sun, M.Y.; Li, J.M.; Song, B.B.; Liu, Y.Y.; Tian, Y.K.; Wang, C.H.; Wu, J. Comparative transcriptome analysis 1provides insights into the mechanism of pear dwarfing. J. Integr. Agric. 2022, 21, 1952–1967. Available online: https://www.journals.elsevier.com/journal-of-integrative-agriculture (accessed on 21 August 2023). [CrossRef]
- Qu, G.Y.; Peng DYu, Z.Q.; Chen, X.L.; Cheng, X.R.; Yang, Y.Z.; Ye, T.; Lv, Q.; Ji, W.J.; Deng, X.W.; Zhou, B. Advances in the role of auxin for transcriptional regulation of lignin biosynthesis. Funct. Plant Biol. 2021, 48, 743–754. [Google Scholar] [CrossRef]
- Yang, S.L.; Zhang, X.N.; Lu, G.L.; Wang, C.R.; Wang, R. Regulation of gibberellin on gene expressions related with the lignin biosynthesis in ‘Wangkumbae’ pear (Pyrus pyrifolia Nakai) fruit. Plant Growth Regul. 2015, 76, 127–134. [Google Scholar] [CrossRef]
- Li, G.H.; Wang, H.; Cheng, X.; Su, X.Q.; Zhao, Y.; Jiang, T.S.; Jin, Q.; Lin, Y.; Cai, Y.P. Comparative genomic analysis of the PAL genes in five Rosaceae species and functional identification of Chinese white pear. PeerJ 2019, 7, e8064. [Google Scholar] [CrossRef]
- Weng, J.K.; Chapple, C. The origin and evolution of lignin biosynthesis. New Phytol. 2010, 187, 273–285. [Google Scholar] [CrossRef]
- Cheng, X.; Li, M.L.; Li, D.H.; Zhang, J.Y.; Jin, L.; Sheng, L.L.; Cai, Y.P.; Lin, Y. Characterization and analysis of CCR and CAD gene families at the whole-genome level for lignin synthesis of stone cells in pear (Pyrus bretschneideri) fruit. Biol. Open 2017, 6, 1602–1613. Available online: https://www.x-mol.com/paperRedirect/1212918904181825540 (accessed on 21 August 2023). [CrossRef]
- Ohtani, M.; Demura, T. The quest for transcriptional hubs of lignin biosynthesis: Beyond the NAC-MYB-gene regulatory network model. Curr. Opin. Biotechnol. 2019, 56, 82–87. [Google Scholar] [CrossRef]
- Ko, J.H.; Jeon, H.W.; Kim, W.C.; Kim, J.Y.; Han, K.H. The MYB46/MYB83-mediated transcriptional regulatory programme is a gatekeeper of secondary wall biosynthesis. Ann. Bot. 2014, 114, 1099–1107. [Google Scholar] [CrossRef] [PubMed]
- Ko, J.H.; Kim, W.C.; Kim, J.Y.; Ahn, S.J.; Han, K.H. MYB46-mediated transcriptional regulation of secondary wall biosynthesis. Mol. Plant 2012, 5, 961–963. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, R.L.; Zhong, R.; Ye, Z.H. MYB83 is a direct target of SND1 and acts redundantly with MYB46 in the regulation of secondary cell wall biosynthesis in Arabidopsis. Plant Cell Physiol. 2009, 50, 1950–1964. [Google Scholar] [CrossRef]
- Yu, Y.B.; Zhang, S.; Yu, Y.; Cai, N.; Yu, G.C.; Zhao, H.Y.; Meng, X.N.; Fan, H.Y. The pivotal role of MYB transcription factors in plant disease resistance. Planta 2023, 258, 16. [Google Scholar] [CrossRef] [PubMed]
- Xiao, R.; Zhang, C.; Guo, X.; Li, H.; Lu, H. MYB Transcription Factors and Its Regulation in Secondary Cell Wall Formation and Lignin Biosynthesis during Xylem Development. Int. J. Mol. Sci. 2021, 22, 3560. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Han, Y.; Li, D.; Lin, Y.; Cai, Y. MYB Transcription Factors in Chinese Pear (Pyrus bretschneideri Rehd.): Genome-Wide Identification, Classification, and Expression Profiling during Fruit Development. Front. Plant Sci. 2016, 7, 577. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Lam, P.Y.; Lee, M.-H.; Jeon, H.S.; Tobimatsu, Y.; Park, O.K. The Arabidopsis R2R3 MYB Transcription Factor MYB15 Is a Key Regulator of Lignin Biosynthesis in Effector-Triggered Immunity. Front. Plant Sci. 2020, 11, 583153. [Google Scholar] [CrossRef] [PubMed]
- Llerena, J.P.P.; Figueiredo, R.; Ferreira, S.S.; Cesarino, I.; Mazzafera, P. Isolation of Promoters and Transcription Factors Involved in the Regulation of Lignin Biosynthesis in Saccharum Species. Methods Mol. Biol. 2022, 2469, 103–118. [Google Scholar] [CrossRef]
- Deluc, L.; Barrieu, F.; Marchive, C.; Lauvergeat, V.; Decendit, A.; Richard, T.; Carde, J.-P.; Mérillon, J.-M.; Hamdi, S. Characterization of a Grapevine R2R3-MYB Transcription Factor That Regulates the Phenylpropanoid Pathway. Plant Physiol. 2006, 140, 499–511. [Google Scholar] [CrossRef]
- Li, M.; Cheng, C.; Zhang, X.; Zhou, S.; Wang, C.; Ma, C.; Yang, S. PpNAC187 Enhances Lignin Synthesis in ‘Whangkeumbae’ Pear (Pyrus pyrifolia) ‘Hard-End’ Fruit. Molecules 2019, 24, 4338. [Google Scholar] [CrossRef]
- Mitsuda, N.; Iwase, A.; Yamamoto, H.; Yoshida, M.; Seki, M.; Shinozaki, K.; Ohme-Takagi, M. NAC Transcription Factors, NST1 and NST3, Are Key Regulators of the Formation of Secondary Walls in Woody Tissues of Arabidopsis. Plant Cell 2007, 19, 270–280. [Google Scholar] [CrossRef]
- Zhong, R.; Ye, Z.-H. Transcriptional regulation of lignin biosynthesis. Plant Signal. Behav. 2009, 4, 1028–1034. [Google Scholar] [CrossRef] [PubMed]
- Hirano, K.; Kondo, M.; Aya, K.; Miyao, A.; Sato, Y.; Antonio, B.A.; Namiki, N.; Nagamura, Y.; Matsuoka, M. Identification of Transcription Factors Involved in Rice Secondary Cell Wall Formation. Plant Cell Physiol. 2013, 54, 1791–1802. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Xu, C.; Kang, Y.; Gu, T.; Wang, D.; Zhao, S.; Xia, G. The heterologous expression in Arabidopsis thaliana of sorghum transcription factor SbbHLH1 downregulates lignin synthesis. J. Exp. Bot. 2013, 64, 3021–3032. [Google Scholar] [CrossRef]
- Cao, Y.; Meng, D.; Li, X.; Wang, L.; Cai, Y.; Jiang, L. A Chinese White Pear (Pyrus bretschneideri) BZR Gene PbBZR1 Act as a Transcriptional Repressor of Lignin Biosynthetic Genes in Fruits. Front. Plant Sci. 2020, 11, 1087. [Google Scholar] [CrossRef]
- Konarska, A. The Relationship between the Morphology and Structure and the Quality of Fruits of Two Pear Cultivars (Pyrus communis L.) during Their Development and Maturation. Sci. World J. 2013, 2013, 846796. [Google Scholar] [CrossRef]
- Lewandowska, M.; Keyl, A.; Feussner, I. Wax biosynthesis in response to danger: Its regulation upon abiotic and biotic stress. New Phytol. 2020, 227, 698–713. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, P.; Nguyen, N.; Hykkerud, A.L.; Häggman, H.; Martinussen, I.; Jaakola, L.; Karppinen, K. Developmental and Environmental Regulation of Cuticular Wax Biosynthesis in Fleshy Fruits. Front. Plant Sci. 2019, 10, 431. [Google Scholar] [CrossRef]
- Mao, H.; Lv, Y.; Chen, G.; Jiang, Y. Effects of cuticular wax on the postharvest physiology in fragrant pear at different storages. J. Sci. Food Agric. 2022, 102, 4425–4434. [Google Scholar] [CrossRef]
- Jiao, J.; Jin, M.; Liu, H.; Suo, J.; Yin, X.; Zhu, Q.; Rao, J. Application of melatonin in kiwifruit (Actinidia chinensis) alleviated chilling injury during cold storage. Sci. Hortic. 2022, 296, 110876. [Google Scholar] [CrossRef]
- Wang, D.; Chen, Q.; Chen, W.; Guo, Q.; Xia, Y.; Wu, D.; Jing, D.; Liang, G. Melatonin treatment maintains quality and delays lignification in loquat fruit during cold storage. Sci. Hortic. 2021, 284, 110126. Available online: https://www.x-mol.com/paperRedirect/1375237835992809472 (accessed on 21 August 2023). [CrossRef]
- Höll, J.; Vannozzi, A.; Czemmel, S.; D’Onofrio, C.; Walker, A.R.; Rausch, T.; Lucchin, M.; Boss, P.K.; Dry, I.B.; Bogs, J. The R2R3-MYB Transcription Factors MYB14 and MYB15 Regulate Stilbene Biosynthesis in Vitis vinifera. Plant Cell 2013, 25, 4135–4149. [Google Scholar] [CrossRef] [PubMed]
- Bate, N.J.; Orr, J.; Ni, W.; Meromi, A.; Nadler-Hassar, T.; Doerner, P.W.; A Dixon, R.; Lamb, C.J.; Elkind, Y. Quantitative relationship between phenylalanine ammonia-lyase levels and phenylpropanoid accumulation in transgenic tobacco identifies a rate-determining step in natural product synthesis. Proc. Natl. Acad. Sci. USA 1994, 91, 7608–7612. [Google Scholar] [CrossRef] [PubMed]
- Wanner, L.A.; Li, G.; Ware, D.; Somssich, I.E.; Davis, K.R. The phenylalanine ammonia-lyase gene family in Arabidopsis thaliana. Plant Mol. Biol. 1995, 27, 327–338. [Google Scholar] [CrossRef]
- Cheng, X.; Li, G.; Ma, C.; Abdullah, M.; Zhang, J.; Zhao, H.; Jin, Q.; Cai, Y.; Lin, Y. Comprehensive genome-wide analysis of the pear (Pyrus bretschneideri) laccase gene (PbLAC) family and functional identification of PbLAC1 involved in lignin biosynthesis. PLoS ONE 2019, 14, e0210892. [Google Scholar] [CrossRef]
- Li, M.; Cheng, C.; Zhang, X.; Zhou, S.; Li, L.; Yang, S. Overexpression of Pear (Pyrus pyrifolia) CAD2 in Tomato Affects Lignin Content. Molecules 2019, 24, 2595. [Google Scholar] [CrossRef]
- Su, X.; Zhao, Y.; Wang, H.; Li, G.; Cheng, X.; Jin, Q.; Cai, Y. Transcriptomic analysis of early fruit development in Chinese white pear (Pyrus bretschneideri Rehd.) and functional identification of PbCCR1 in lignin biosynthesis. BMC Plant Biol. 2019, 19, 417. [Google Scholar] [CrossRef]
- Li, W.; Hao, Z.Y.; Yang, L.C.; Xia, H.; Tu, Z.H.; Cui, Z.K.; Wu, J.P.; Li, H.G. Genome-wide identification and characterization of LcCCR13 reveals its potential role in lignin biosynthesis in Liriodendron chinense. Front Plant Sci. 2023, 13, 1110639. [Google Scholar] [CrossRef]
- Li, H.; Suo, J.; Han, Y.; Liang, C.; Jin, M.; Zhang, Z.; Rao, J. The effect of 1-methylcyclopropene, methyl jasmonate and methyl salicylate on lignin accumulation and gene expression in postharvest ‘Xuxiang’ kiwifruit during cold storage. Postharvest Biol. Technol. 2017, 124, 107–118. [Google Scholar] [CrossRef]
- Sitbon, F.; Hennion, S.; Little, C.; Sundberg, B. Enhanced ethylene production and peroxidase activity in IAA-overproducing transgenic tobacco plants is associated with increased lignin content and altered lignin composition. Plant Sci. 1999, 141, 165–173. [Google Scholar] [CrossRef]
- Elkind, Y.; Edwards, R.; Mavandad, M.; A Hedrick, S.; Ribak, O.; A Dixon, R.; Lamb, C.J. Abnormal plant development and down-regulation of phenylpropanoid biosynthesis in transgenic tobacco containing a heterologous phenylalanine ammonia-lyase gene. Proc. Natl. Acad. Sci. USA 1990, 87, 9057–9061. [Google Scholar] [CrossRef]
- Li, S.; Yang, N.; Chen, L. Paraffin section observation of flower bud differentiation of Chimonanthus praecox in Kunming and comparison of the differentiation processes in different regions, China. Hortic. Plant J. 2022, 8, 221–229. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Shirasawa, K.; Itai, A.; Isobe, S. Chromosome-scale genome assembly of Japanese pear (Pyrus pyrifolia) variety ‘Nijisseiki’. DNA Res. 2021, 28, dsab001. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; Van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Gao, X.; Zhang, X.; Yang, J.; Wang, L.; Wang, S.; Wang, L.; Xue, H. Fine mapping and candidate gene analysis of flesh browning in pear (Pyrus L.). Sci. Hortic. 2022, 302, 111140. [Google Scholar] [CrossRef]
- Zhang, X.; Li, B.; Duan, R.; Han, C.; Wang, L.; Yang, J.; Wang, L.; Wang, S.; Su, Y.; Xue, H. Transcriptome Analysis Reveals Roles of Sucrose in Anthocyanin Accumulation in ‘Kuerle Xiangli’ (Pyrus sinkiangensis Yü). Genes 2022, 13, 1064. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Raw Data | Clean Data (%) | Q20 (%) | Q30 (%) | GC (%) | Total Mapped (%) |
|---|---|---|---|---|---|---|
| Normal 1 | 44,838,532 | 44,699,178 (99.69%) | 97.86% | 93.75% | 46.45% | 41,436,589 (92.77%) |
| Normal 2 | 53,906,118 | 53,762,512 (99.73%) | 94.04% | 94.04% | 46.51% | 49,783,567 (92.68%) |
| Normal 3 | 47,423,092 | 47,287,074 (99.71%) | 97.92% | 93.86% | 46.56% | 43,925,452 (92.96%) |
| Protrudent 1 | 43,242,926 | 43,136,608 (99.75%) | 98.06% | 94.18% | 46.16% | 40,063,778 (92.94%) |
| Protrudent 2 | 44,343,242 | 44,233,162 (99.75%) | 97.85% | 93.68% | 46.24% | 41,018,154 (92.79%) |
| Protrudent 3 | 48,156,128 | 48,026,282 (99.73%) | 97.88% | 93.78% | 46.18% | 44,482,464 (92.71%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duan, R.; Zhang, X.; Liu, Y.; Wang, L.; Yang, J.; Wang, L.; Wang, S.; Su, Y.; Xue, H. Transcriptome and Physiological Analysis Highlight Lignin Metabolism of the Fruit Dots Disordering during Postharvest Cold Storage in ‘Danxiahong’ Pear. Genes 2023, 14, 1785. https://doi.org/10.3390/genes14091785
Duan R, Zhang X, Liu Y, Wang L, Yang J, Wang L, Wang S, Su Y, Xue H. Transcriptome and Physiological Analysis Highlight Lignin Metabolism of the Fruit Dots Disordering during Postharvest Cold Storage in ‘Danxiahong’ Pear. Genes. 2023; 14(9):1785. https://doi.org/10.3390/genes14091785
Chicago/Turabian StyleDuan, Ruiwei, Xiangzhan Zhang, Yudong Liu, Lei Wang, Jian Yang, Long Wang, Suke Wang, Yanli Su, and Huabai Xue. 2023. "Transcriptome and Physiological Analysis Highlight Lignin Metabolism of the Fruit Dots Disordering during Postharvest Cold Storage in ‘Danxiahong’ Pear" Genes 14, no. 9: 1785. https://doi.org/10.3390/genes14091785
APA StyleDuan, R., Zhang, X., Liu, Y., Wang, L., Yang, J., Wang, L., Wang, S., Su, Y., & Xue, H. (2023). Transcriptome and Physiological Analysis Highlight Lignin Metabolism of the Fruit Dots Disordering during Postharvest Cold Storage in ‘Danxiahong’ Pear. Genes, 14(9), 1785. https://doi.org/10.3390/genes14091785