
Time-Series Transcriptome of Cucumis melo Reveals Extensive Transcriptomic Differences with Different Maturity


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Measurement of the Quality of Melon Flesh
2.3. RNA Extraction and Sequencing
2.4. Differentially Expressed Gene Identification
2.5. Characterization of Alternative Splicing Events
2.6. Enrichment Analysis of Differentially Expressed Genes and Alternative Splicing (AS) Analysis
3. Results
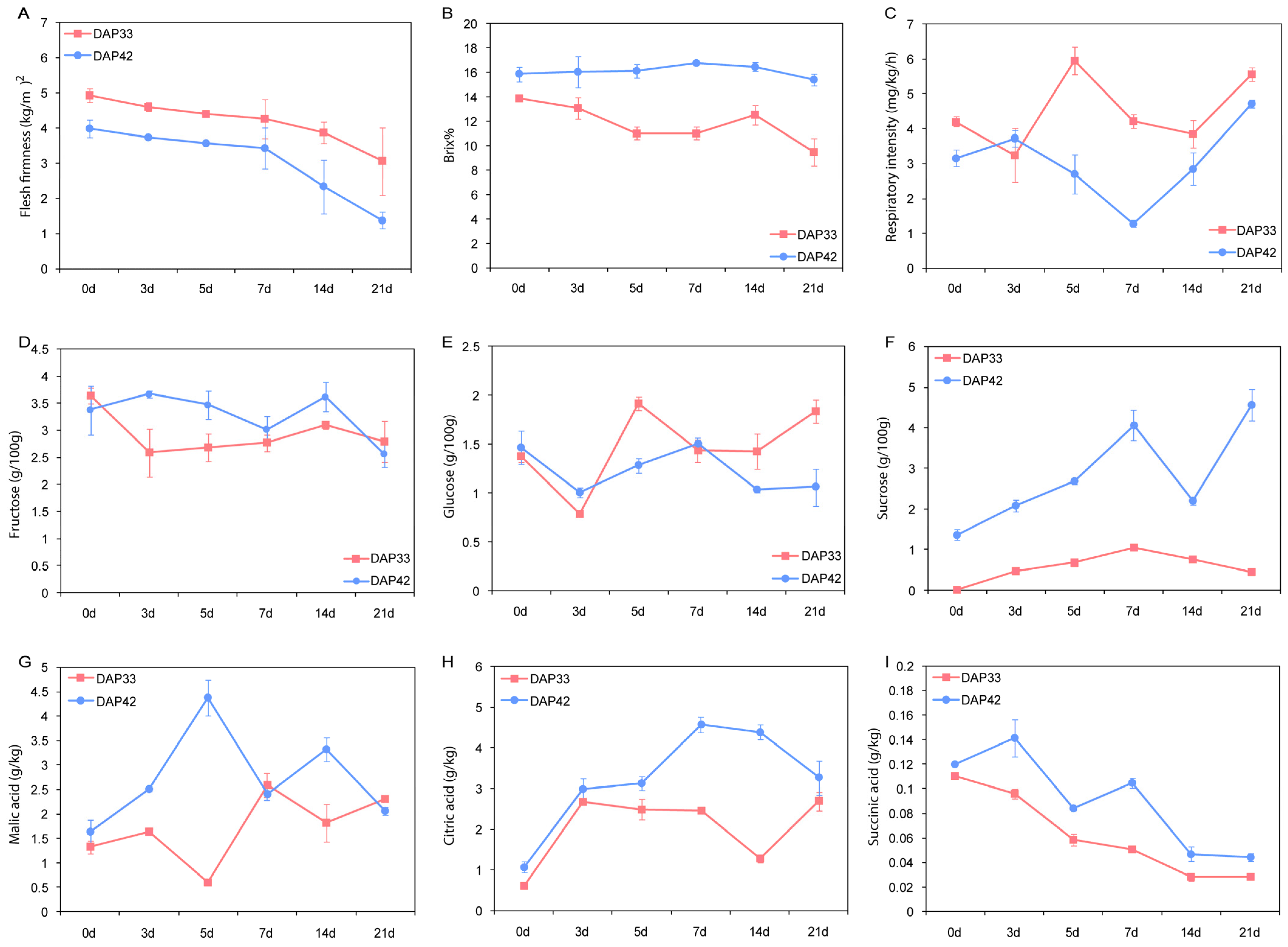
3.1. Rapid Changes in Fruit Firmness and Metabolic Profiles between 60% and 90% Maturity
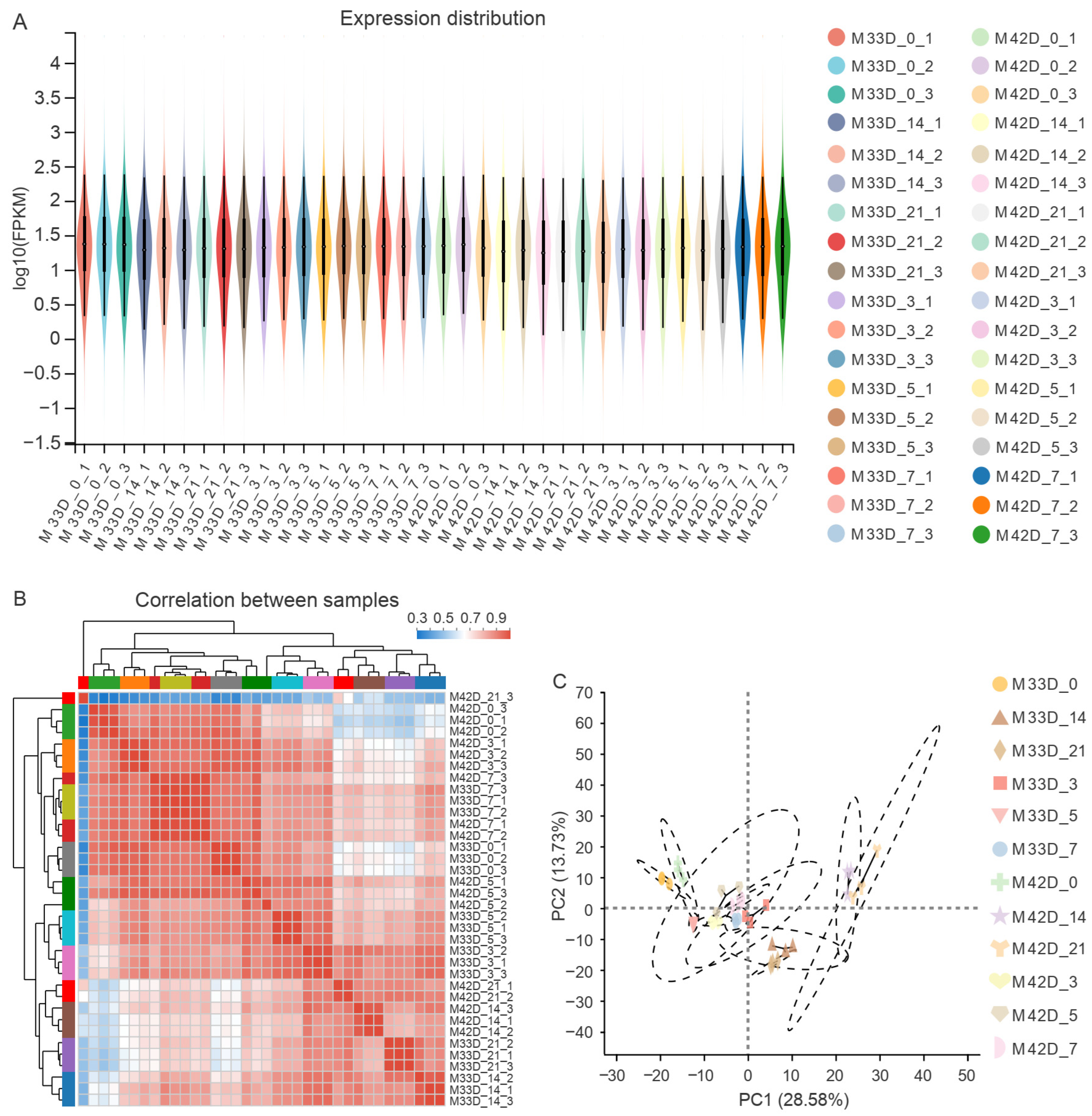
3.2. Transcriptome Profiles of Storage between 60% and 90% Melon Maturity
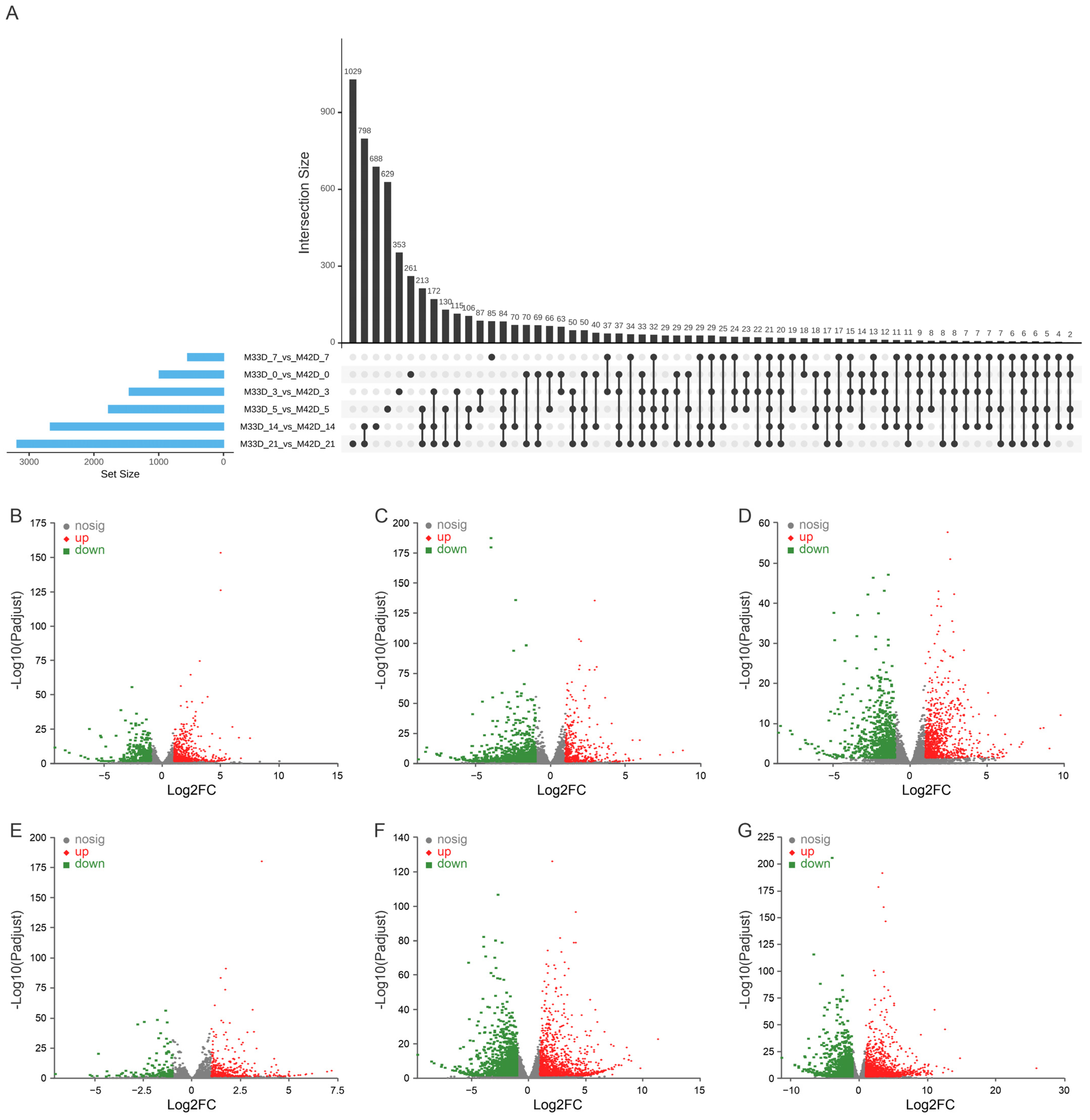
3.3. Differentially Expressed Genes between 60% and 90% Maturity Melons
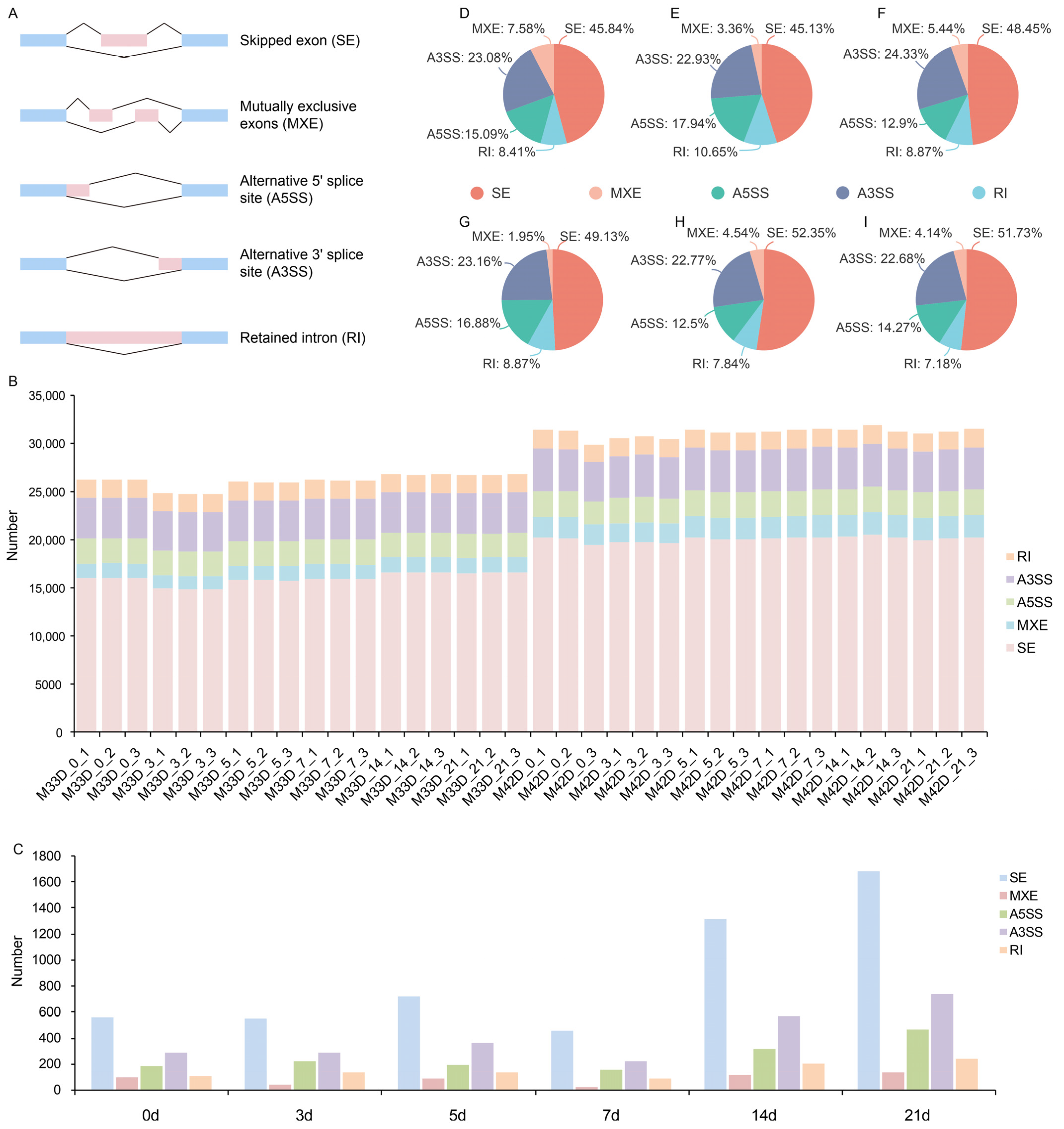
3.4. Alternative Splicing between 60% and 90% Melon Maturity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Garcia-Mas, J.; Benjak, A.; Sanseverino, W.; Bourgeois, M.; Mir, G.; González, V.M.; Hénaff, E.; Câmara, F.; Cozzuto, L.; Lowy, E.; et al. The genome of melon (Cucumis melo L.). Proc. Natl. Acad. Sci. USA 2012, 109, 11872–11877. [Google Scholar] [CrossRef] [PubMed]
- Sebastian, P.; Schaefer, H.; Telford, I.R.; Renner, S.S. Cucumber (Cucumis sativus) and melon (C. melo) have numerous wild relatives in Asia and Australia, and the sister species of melon is from Australia. Proc. Natl. Acad. Sci. USA 2010, 107, 14269–14273. [Google Scholar] [CrossRef] [PubMed]
- Schaffer, A.A.; Paris, H.S. Melons, squashes, and gourds. In Encyclopedia of Food Sciences and Nutrition, 2nd ed.; Academic Press: Cambridge, MA, USA, 2003. [Google Scholar]
- Sageret, A. Considérations sur la production des hybrides, des variantes et des variétés en général, et sur celles de la famille des Cucurbitacées en particulier [Considerations on the production of hybrids, variants and varieties in general and those of the Cucurbitaceae family in particular]. In Annales des Sciences Naturelles; Biodiversity Heritage Library: Washington, DC, USA, 1826; Volume 8, p. 294. [Google Scholar]
- Boualem, A.; Fergany, M.; Fernandez, R.; Troadec, C.; Martin, A.; Morin, H.; Sari, M.A.; Collin, F.; Flowers, J.M.; Pitrat, M.; et al. A conserved mutation in an ethylene biosynthesis enzyme leads to andromonoecy in melons. Science 2008, 321, 836–838. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.; Troadec, C.; Boualem, A.; Rajab, M.; Fernandez, R.; Morin, H.; Pitrat, M.; Dogimont, C.; Bendahmane, A. A transposon-induced epigenetic change leads to sex determination in melon. Nature 2009, 461, 1135–1138. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Tolstikov, V.; Turnbull, C.; Hicks, L.M.; Fiehn, O. Divergent metabolome and proteome suggest functional independence of dual phloem transport systems in cucurbits. Proc. Natl. Acad. Sci. USA 2010, 107, 13532–13537. [Google Scholar] [CrossRef] [PubMed]
- Pech, J.C.; Bouzayen, M.; Latché, A.J.P.S. Climacteric fruit ripening: Ethylene-dependent and independent regulation of ripening pathways in melon fruit. Plant Sci. 2008, 175, 114–120. [Google Scholar] [CrossRef]
- McMurchie, E.J.; McGlasson, W.B.; Eaks, I.L. Treatment of fruit with propylene gives information about the biogenesis of ethylene. Nature 1972, 237, 235–236. [Google Scholar] [CrossRef] [PubMed]
- Giovannoni, J.J. Genetic regulation of fruit development and ripening. Plant Cell 2004, 16, S170–S180. [Google Scholar] [CrossRef]
- Saladié, M.; Cañizares, J.; Phillips, M.A.; Rodriguez-Concepcion, M.; Larrigaudière, C.; Gibon, Y.; Stitt, M.; Lunn, J.E.; Garcia-Mas, J. Comparative transcriptional profiling analysis of developing melon (Cucumis melo L.) fruit from climacteric and non-climacteric varieties. BMC Genom. 2015, 16, 440. [Google Scholar] [CrossRef]
- Leida, C.; Moser, C.; Esteras, C.; Sulpice, R.; Lunn, J.E.; de Langen, F.; Monforte, A.J.; Picó, B. Variability of candidate genes, genetic structure and association with sugar accumulation and climacteric behavior in a broad germplasm collection of melon (Cucumis melo L.). BMC Genet. 2015, 16, 28. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Hughes, D.L.; Yabumoto, K.; Jennings, W.G. Quality of cantaloupe muskmelons: Variability and attributes. Sci. Hortic. 1977, 6, 59–70. [Google Scholar] [CrossRef]
- Li, Y.; Huang, H.; Abid, M.; Gu, H.; Fang, J.; Cheng, Z.; Qi, X. Characterization and Identification of a Ripening-Related Gene AaPG18 in Actinidia arguta. Int. J. Mol. Sci. 2022, 23, 2597. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Barbazuk, W.B.; Fu, Y.; McGinnis, K.M. Genome-wide analyses of alternative splicing in plants: Opportunities and challenges. Genome Res. 2008, 18, 1381–1392. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.; Park, J.W.; Lu, Z.X.; Lin, L.; Henry, M.D.; Wu, Y.N.; Zhou, Q.; Xing, Y. rMATS: Robust and flexible detection of differential alternative splicing from replicate RNA-Seq data. Proc. Natl. Acad. Sci. USA 2014, 111, E5593–E5601. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef]
- White, P.J. Recent advances in fruit development and ripening: An overview. J. Exp. Bot. 2002, 53, 1995–2000. [Google Scholar] [CrossRef]
- Cheng, H.; Kong, W.; Tang, T.; Ren, K.; Zhang, K.; Wei, H.; Lin, T. Identification of Key Gene Networks Controlling Soluble Sugar and Organic Acid Metabolism During Oriental Melon Fruit Development by Integrated Analysis of Metabolic and Transcriptomic Analyses. Front. Plant. Sci. 2022, 13, 830517. [Google Scholar] [CrossRef]
- Zhang, A.; Zheng, J.; Chen, X.; Shi, X.; Wang, H.; Fu, Q. Comprehensive Analysis of Transcriptome and Metabolome Reveals the Flavonoid Metabolic Pathway Is Associated with Fruit Peel Coloration of Melon. Molecules 2021, 26, 2830. [Google Scholar] [CrossRef]
- Shin, A.Y.; Kim, Y.M.; Koo, N.; Lee, S.M.; Nahm, S.; Kwon, S.Y. Transcriptome analysis of the oriental melon (Cucumis melo L. var. makuwa) during fruit development. PeerJ 2017, 5, e2834. [Google Scholar] [CrossRef] [PubMed]
- Burger, Y.; Schaffer, A.A. The contribution of sucrose metabolism enzymes to sucrose accumulation in Cucumis melo. J. Am. Soc. Hortic. Sci. 2007, 132, 704–712. [Google Scholar] [CrossRef]
- Hubbard, N.L.; Huber, S.C.; Pharr, D.M. Sucrose Phosphate Synthase and Acid Invertase as Determinants of Sucrose Concentration in Developing Muskmelon (Cucumis melo L.) Fruits. Plant Physiol. 1989, 91, 1527–1534. [Google Scholar] [CrossRef] [PubMed]
- Rosa, J. Changes in composition during ripening and storage of melons. Hilgardia 1928, 3, 421–443. [Google Scholar] [CrossRef]
- Schaffer, A.A.; Aloni, B.; Fogelman, E. Sucrose metabolism and accumulation in developing fruit of Cucumis. Phytochemistry 1987, 26, 1883–1887. [Google Scholar] [CrossRef]
- Schaffer, A.A.; Madore, M.; Pharr, D.M. Cucurbits. In Photoassimilate Distribution in Plants and Crops; Zamski, E., Schaffer, A.A., Eds.; CRC Press: New York, NY, USA, 1996; pp. 729–757. [Google Scholar]
- Cohen, S.; Tzuri, G.; Harel-Beja, R.; Itkin, M.; Portnoy, V.; Sa’ar, U.; Lev, S.; Yeselson, L.; Petrikov, M.; Rogachev, I.; et al. Co-mapping studies of QTLs for fruit acidity and candidate genes of organic acid metabolism and proton transport in sweet melon (Cucumis melo L.). Theor. Appl. Genet. 2012, 125, 343–353. [Google Scholar] [CrossRef]
- Burger, Y.; Paris, H.S.; Cohen, R.; Katzir, N.; Tadmor, Y.; Lewinsohn, E.; Schaffer, A.A. 3 Genetic Diversity of Cucumis melo. Hortic. Rev. 2010, 36, 165–198. [Google Scholar]
- Danin-Poleg, Y.; Tadmor, Y.; Tzuri, G.; Reis, N.; Hirschberg, J.; Katzir, N. Construction of a genetic map of melon with molecular markers and horticultural traits, and localization of genes associated with ZYMV resistance. Euphytica 2002, 125, 373–384. [Google Scholar] [CrossRef]
- Ramaswamy, B.; Seshadri, V.S.; Sharma, J.C. Inheritance of some fruit characters in muskmelon. Sci. Hortic. 1977, 6, 107–120. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, H.; Yi, H.; Zhai, W.; Wang, G.; Fu, Q. Transcriptome profiling of Cucumis melo fruit development and ripening. Hortic. Res. 2016, 3, 16014. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene Name | Gene Description |
|---|---|---|
| MELO3C024383 | CmCIN2 | Cell wall invertase 2 |
| MELO3C020357 | CmSPS2 | Sucrose-P synthase 2 |
| MELO3C015469 | CmBGAL3 | β-galactosidase-like |
| MELO3C020357 | CmSPS2 | Sucrose-phosphate synthase |
| MELO3C001956 | CmSUS-LIKE1 | Sucrose synthase |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, F.; Shao, X.; Fan, Y.; Jia, B.; He, W.; Wang, Y.; Wang, F.; Wang, C. Time-Series Transcriptome of Cucumis melo Reveals Extensive Transcriptomic Differences with Different Maturity. Genes 2024, 15, 149. https://doi.org/10.3390/genes15020149
Liu F, Shao X, Fan Y, Jia B, He W, Wang Y, Wang F, Wang C. Time-Series Transcriptome of Cucumis melo Reveals Extensive Transcriptomic Differences with Different Maturity. Genes. 2024; 15(2):149. https://doi.org/10.3390/genes15020149
Chicago/Turabian StyleLiu, Fengjuan, Xupeng Shao, Yingying Fan, Binxin Jia, Weizhong He, Yan Wang, Fengzhong Wang, and Cheng Wang. 2024. "Time-Series Transcriptome of Cucumis melo Reveals Extensive Transcriptomic Differences with Different Maturity" Genes 15, no. 2: 149. https://doi.org/10.3390/genes15020149
APA StyleLiu, F., Shao, X., Fan, Y., Jia, B., He, W., Wang, Y., Wang, F., & Wang, C. (2024). Time-Series Transcriptome of Cucumis melo Reveals Extensive Transcriptomic Differences with Different Maturity. Genes, 15(2), 149. https://doi.org/10.3390/genes15020149