
Preservation of 5-Hydroxymethylcytosine Levels in LRIG1 across Genomic DNA and Cell-Free DNA in Glioma Patients


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
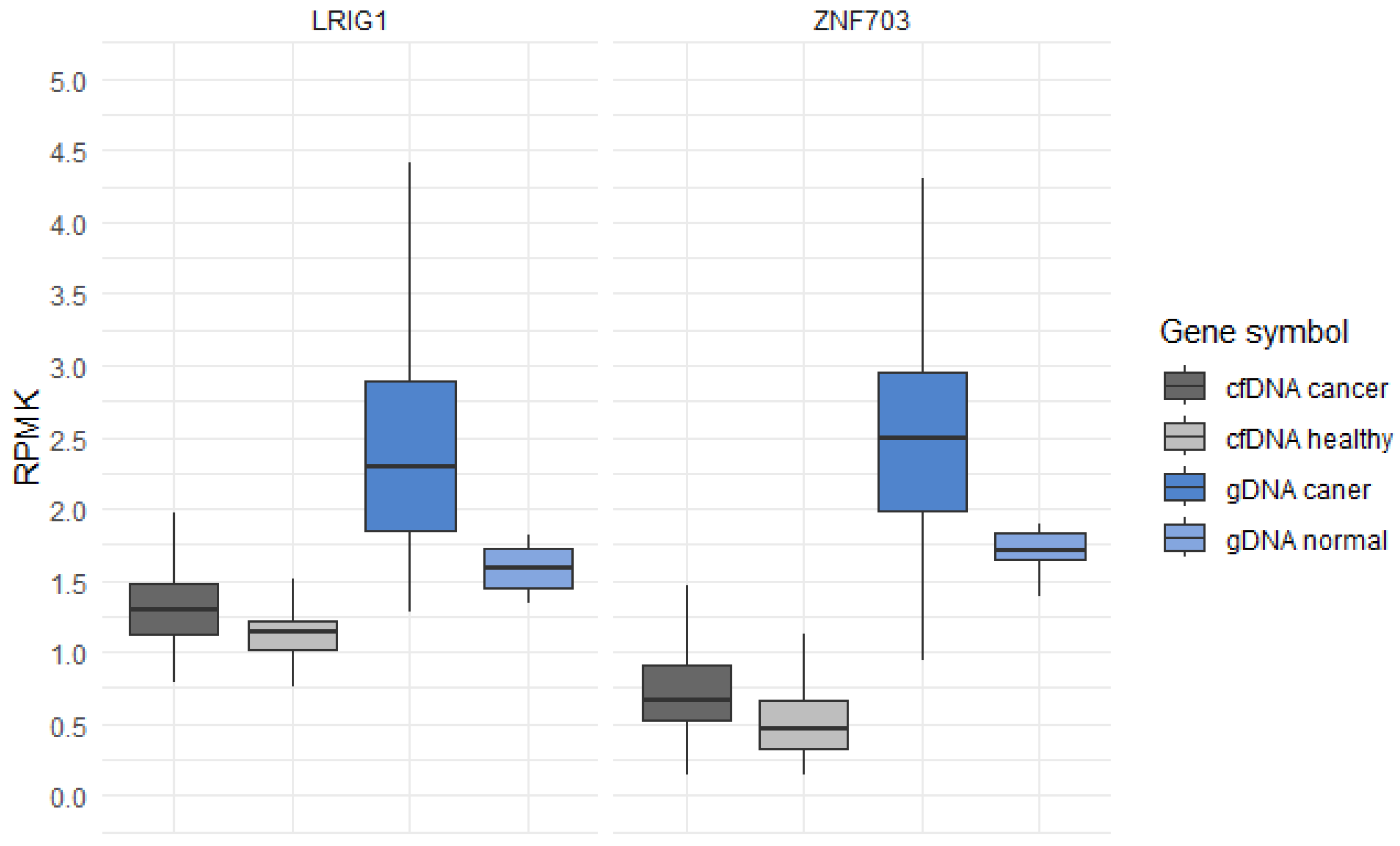
3.1. 5hmC Profiles in cfDNA and gDNA of Glioma Patients
3.2. Interrelation between 5hmC Levels and Gene Expression in Glioma Patients
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lin, I.H.; Chen, Y.F.; Hsu, M.T. Correlated 5-Hydroxymethylcytosine (5hmC) and Gene Expression Profiles Underpin Gene and Organ-Specific Epigenetic Regulation in Adult Mouse Brain and Liver. PLoS ONE 2017, 12, e0170779. [Google Scholar] [CrossRef]
- Nestor, C.E.; Ottaviano, R.; Reddington, J.; Sproul, D.; Reinhardt, D.; Dunican, D.; Katz, E.; Dixon, J.M.; Harrison, D.J.; Meehan, R.R. Tissue type is a major modifier of the 5-hydroxymethylcytosine content of human genes. Genome Res. 2012, 22, 467–477. [Google Scholar] [CrossRef]
- Cai, J.; Zeng, C.; Hua, W.; Qi, Z.; Song, Y.; Lu, X.; Li, D.; Zhang, Z.; Cui, X.; Zhang, X.; et al. An integrative analysis of genome-wide 5-hydroxymethylcytosines in circulating cell-free DNA detects noninvasive diagnostic markers for gliomas. Neurooncol Adv. 2021, 3, vdab049. [Google Scholar] [CrossRef]
- Tang, F.; Wang, H.; Chen, E.; Bian, E.; Xu, Y.; Ji, X.; Yang, Z.; Hua, X.; Zhang, Y.; Zhao, B. LncRNA-ATB promotes TGF-β-induced glioma cells invasion through NF-κB and P38/MAPK pathway. J. Cell. Physiol. 2019, 234, 23302–23314. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, Y.; Wang, X.; Zhang, H.; Wang, S. The up-regulation of LRIG1 expression inhibits the proliferation, apoptosis and invasion of glioma cells. Am. J. Transl. Res. 2022, 14, 788–797. [Google Scholar]
- Ye, F.; Gao, Q.; Xu, T.; Zeng, L.; Ou, Y.; Mao, F.; Wang, H.; He, Y.; Wang, B.; Yang, Z.; et al. Upregulation of LRIG1 suppresses malignant glioma cell growth by attenuating EGFR activity. J. Neurooncol. 2009, 94, 183–194. [Google Scholar] [CrossRef]
- Crake, R.L.I.; Burgess, E.R.; Wiggins, G.A.R.; Magon, N.J.; Das, A.B.; Vissers, M.C.M.; Morrin, H.R.; Royds, J.A.; Slatter, T.L.; Robinson, B.A.; et al. Ascorbate content of clinical glioma tissues is related to tumour grade and to global levels of 5-hydroxymethyl cytosine. Sci. Rep. 2022, 12, 14845. [Google Scholar] [CrossRef]
- Zeng, C.; Song, X.; Zhang, Z.; Cai, Q.; Cai, J.; Horbinski, C.; Hu, B.; Cheng, S.Y.; Zhang, W. Dissection of transcriptomic and epigenetic heterogeneity of grade 4 gliomas: Implications for prognosis. Acta Neuropathol. Commun. 2023, 11, 133. [Google Scholar] [CrossRef]
- Bettegowda, C.; Sausen, M.; Leary, R.J.; Kinde, I.; Wang, Y.; Agrawal, N.; Bartlett, B.R.; Wang, H.; Luber, B.; Alani, R.M.; et al. Detection of Circulating Tumor DNA in Early- and Late-Stage Human Malignancies. Sci. Transl. Med. 2014, 6, 224ra224. [Google Scholar] [CrossRef]
- Smyth, G.K. Linear models and empirical bayes methods for assessing differential expression in microarray experiments. Stat. Appl. Genet. Mol. Biol. 2004, 3, Article3. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2019; Volume 8, 212p. [Google Scholar]
- Ghiglione, C.; Carraway, K.L.; Amundadottir, L.T.; Boswell, R.E.; Perrimon, N.; Duffy, J.B. The Transmembrane Molecule Kekkon 1 Acts in a Feedback Loop to Negatively Regulate the Activity of the Drosophila EGF Receptor during Oogenesis. Cell 1999, 96, 847–856. [Google Scholar] [CrossRef]
- Suzuki, Y.; Sato, N.; Tohyama, M.; Wanaka, A.; Takagi, T. cDNA Cloning of a Novel Membrane Glycoprotein That Is Expressed Specifically in Glial Cells in the Mouse Brain: LIG-1, A PROtein with leucine-rich repeats and immunoglobulin-like domains*. J. Biol. Chem. 1996, 271, 22522–22527. [Google Scholar] [CrossRef]
- Nilsson, J.; Vallbo, C.; Guo, D.; Golovleva, I.; Hallberg, B.; Henriksson, R.; Hedman, H. Cloning, Characterization, and Expression of Human LIG1. Biochem. Biophys. Res. Commun. 2001, 284, 1155–1161. [Google Scholar] [CrossRef]
- Guo, D.; Holmlund, C.; Henriksson, R.; Hedman, H. The LRIG gene family has three vertebrate paralogs widely expressed in human and mouse tissues and a homolog in Ascidiacea. Genomics 2004, 84, 157–165. [Google Scholar] [CrossRef]
- Holmlund, C.; Nilsson, J.; Guo, D.; Starefeldt, A.; Golovleva, I.; Henriksson, R.; Hedman, H. Characterization and tissue-specific expression of human LRIG2. Gene 2004, 332, 35–43. [Google Scholar] [CrossRef]
- Qi, X.-C.; Xie, D.-J.; Yan, Q.-F.; Wang, Y.-R.; Zhu, Y.-X.; Qian, C.; Yang, S.-X. LRIG1 dictates the chemo-sensitivity of temozolomide (TMZ) in U251 glioblastoma cells via down-regulation of EGFR/topoisomerase-2/Bcl-2. Biochem. Biophys. Res. Commun. 2013, 437, 565–572. [Google Scholar] [CrossRef]
- Xie, R.; Yang, H.; Xiao, Q.; Mao, F.; Zhang, S.; Ye, F.; Wan, F.; Wang, B.; Lei, T.; Guo, D. Downregulation of LRIG1 expression by RNA interference promotes the aggressive properties of glioma cells via EGFR/Akt/c-Myc activation. Oncol. Rep. 2013, 29, 177–184. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, Y.; Zhu, K.; Song, L.; Tao, M.; Huang, P.; Pan, Y. Resveratrol inhibits glioma cell growth via targeting LRIG1. J. BUON 2018, 23, 403–409. [Google Scholar]
- Chang, L.; Shi, R.; Yang, T.; Li, F.; Li, G.; Guo, Y.; Lang, B.; Yang, W.; Zhuang, Q.; Xu, H. Restoration of LRIG1 suppresses bladder cancer cell growth by directly targeting EGFR activity. J. Exp. Clin. Cancer Res. 2013, 32, 101. [Google Scholar] [CrossRef]
- Ljuslinder, I.; Golovleva, I.; Palmqvist, R.; Oberg, A.; Stenling, R.; Jonsson, Y.; Hedman, H.; Henriksson, R.; Malmer, B. LRIG1 expression in colorectal cancer. Acta Oncol. 2007, 46, 1118–1122. [Google Scholar] [CrossRef]
- Miller, J.K.; Shattuck, D.L.; Ingalla, E.Q.; Yen, L.; Borowsky, A.D.; Young, L.J.; Cardiff, R.D.; Carraway, K.L., 3rd; Sweeney, C. Suppression of the negative regulator LRIG1 contributes to ErbB2 overexpression in breast cancer. Cancer Res. 2008, 68, 8286–8294. [Google Scholar] [CrossRef]
- Morrison, M.M.; Williams, M.M.; Vaught, D.B.; Hicks, D.; Lim, J.; McKernan, C.; Aurisicchio, L.; Ciliberto, G.; Simion, C.; Sweeney, C.; et al. Decreased LRIG1 in fulvestrant-treated luminal breast cancer cells permits ErbB3 upregulation and increased growth. Oncogene 2016, 35, 1143–1152. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, X.; Zhang, Y.; Wu, J.; Zhou, N. Leucine-rich repeats and immunoglobulin-like domains protein 1 and fascin actin-bundling protein 1 expression in nonsmall cell lung cancer. J. Cancer Res. Ther. 2016, 12, C248–C251. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jevšinek Skok, D.; Bolha, L.; Hauptman, N. Preservation of 5-Hydroxymethylcytosine Levels in LRIG1 across Genomic DNA and Cell-Free DNA in Glioma Patients. Genes 2024, 15, 535. https://doi.org/10.3390/genes15050535
Jevšinek Skok D, Bolha L, Hauptman N. Preservation of 5-Hydroxymethylcytosine Levels in LRIG1 across Genomic DNA and Cell-Free DNA in Glioma Patients. Genes. 2024; 15(5):535. https://doi.org/10.3390/genes15050535
Chicago/Turabian StyleJevšinek Skok, Daša, Luka Bolha, and Nina Hauptman. 2024. "Preservation of 5-Hydroxymethylcytosine Levels in LRIG1 across Genomic DNA and Cell-Free DNA in Glioma Patients" Genes 15, no. 5: 535. https://doi.org/10.3390/genes15050535
APA StyleJevšinek Skok, D., Bolha, L., & Hauptman, N. (2024). Preservation of 5-Hydroxymethylcytosine Levels in LRIG1 across Genomic DNA and Cell-Free DNA in Glioma Patients. Genes, 15(5), 535. https://doi.org/10.3390/genes15050535