
Characterization of the Apoptotic and Antimicrobial Activities of Two Initiator Caspases of Sea Cucumber Apostichopus japonicus


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Gene Cloning
2.2. Bioinformatics and Sequence Analysis
2.3. Transient Transfection
2.4. Flow Cytometry
2.5. Lactate Dehydrogenase (LDH) Assay
2.6. Immunoblotting
2.7. Caspase Activity Analysis
2.8. Recombinant Protein Expression and Purification
2.9. Enzyme-Linked Immunosorbent Assay (ELISA) to Examine Protein–Bacteria Interaction
2.10. Effect of Caspase on Bacterial Growth
2.11. Quantitative Real Time PCR (qRT-PCR)
2.12. Bacterial Infection
2.13. Statistical Analysis
3. Results
3.1. Characterization of Two Caspases from A. japonicus
3.2. AjCASPX1 and AjCASPX2 Possess Unique Structural Features in the CASc Domain
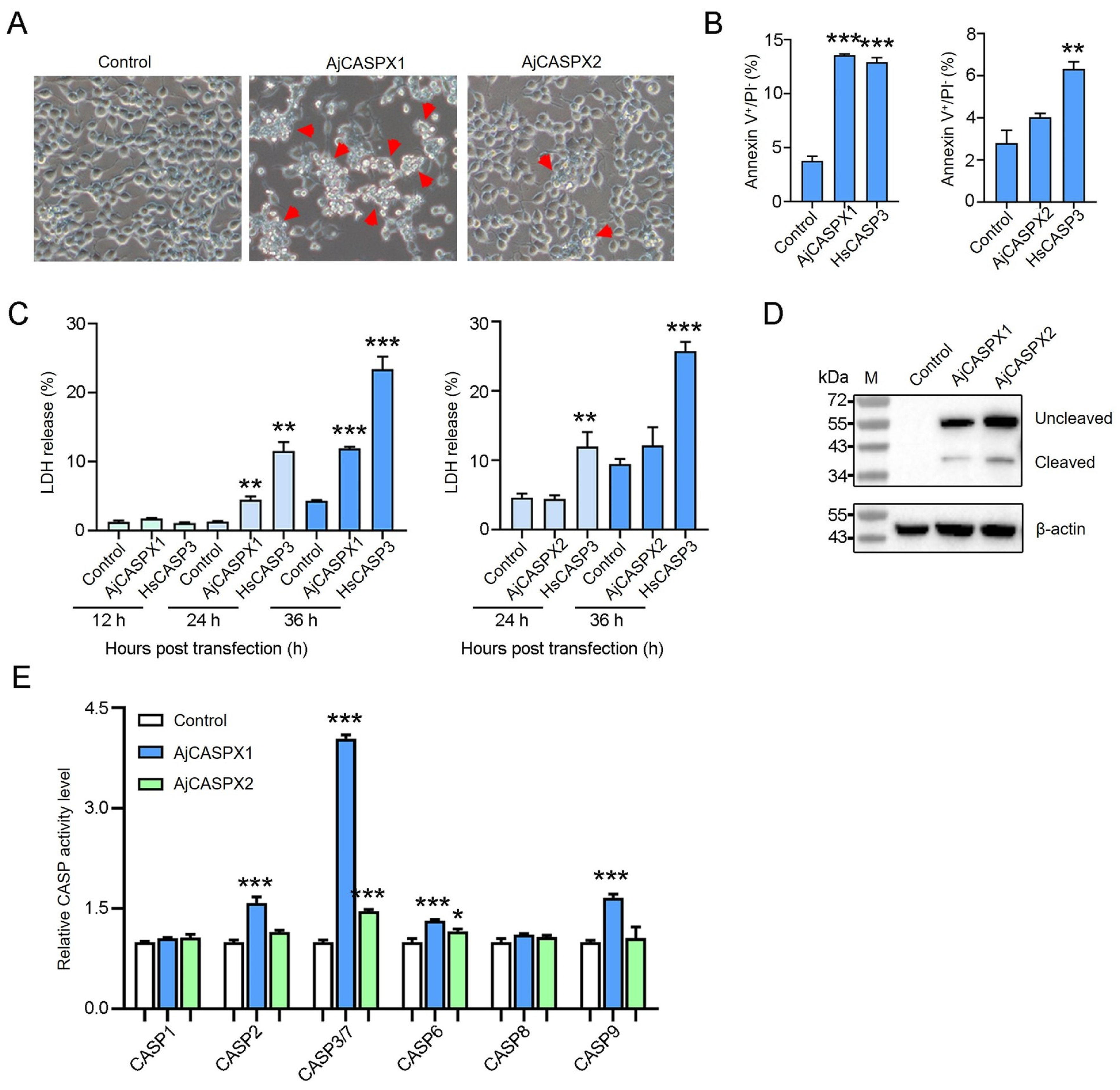
3.3. AjCASPX1 Is Activated and Induces Apoptosis in HEK293T
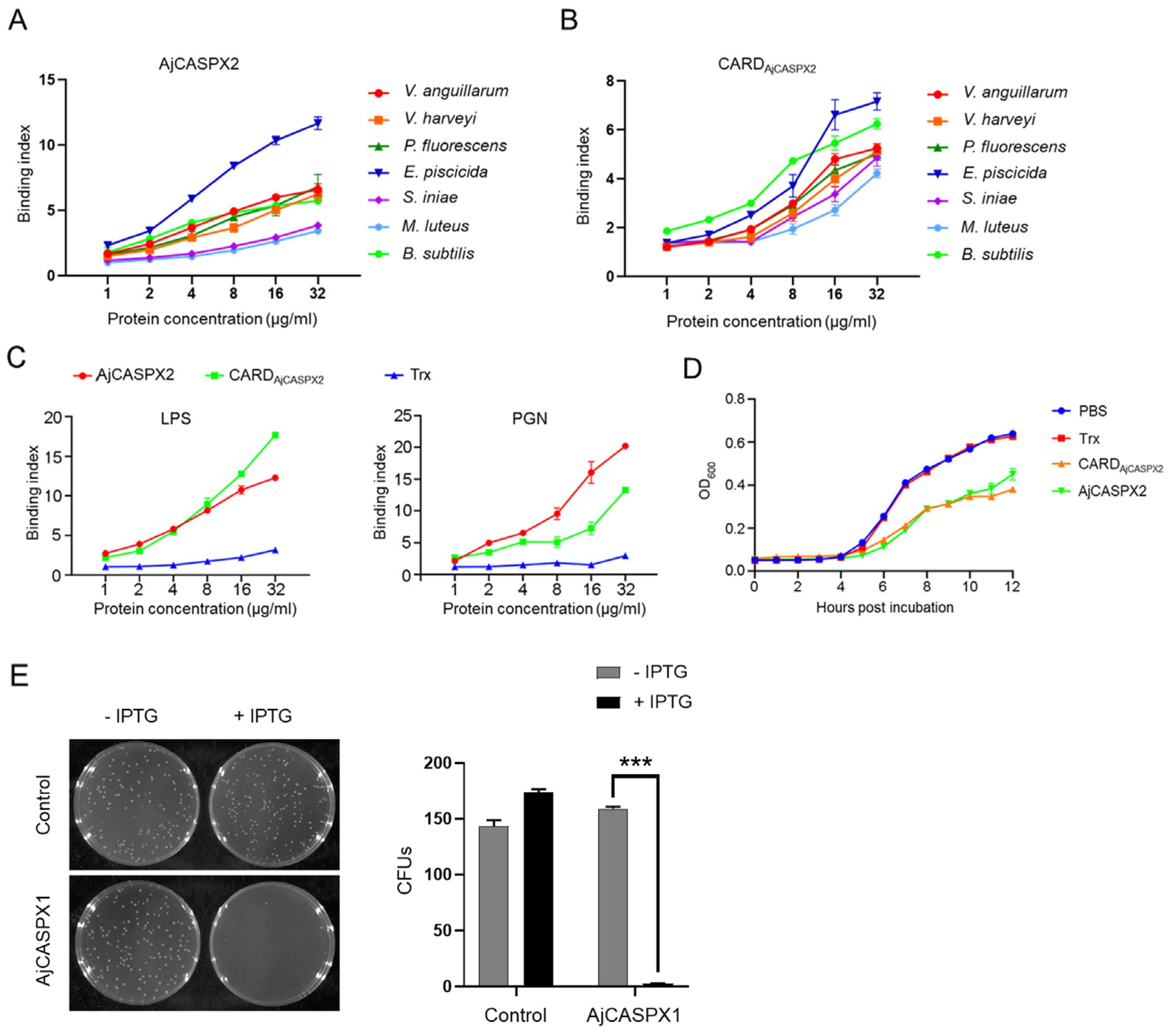
3.4. AjCASPX2 and AjCASPX1 Exhibit Antibacterial Activity
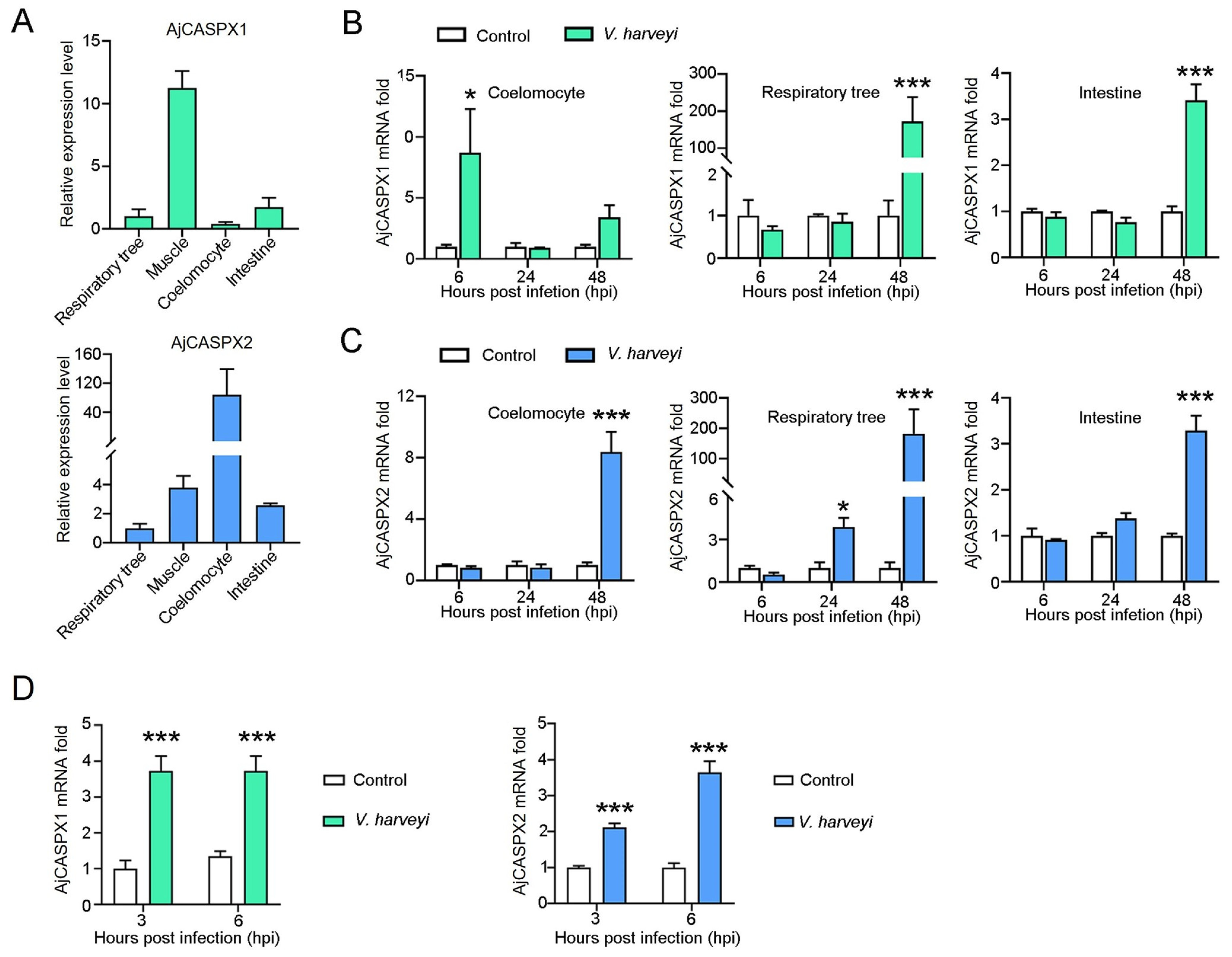
3.5. AjCASPX1 and AjCASPX2 Are Involved in V. harveyi Infection
4. Discussion
5. Conclusions
6. Limitations of the Study
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nagata, S. Apoptosis and Clearance of Apoptotic Cells. Annu. Rev. Immunol. 2018, 36, 489–517. [Google Scholar] [CrossRef] [PubMed]
- Riedl, S.J.; Salvesen, G.S. The Apoptosome: Signalling Platform of Cell Death. Nat. Rev. Mol. Cell Biol. 2007, 8, 405–413. [Google Scholar] [CrossRef]
- Sun, E.W.; Shi, Y.F. Apoptosis: The Quiet Death Silences the Immune System. Pharmacol. Ther. 2001, 92, 135–145. [Google Scholar] [CrossRef]
- Elmore, S. Apoptosis: A Review of Programmed Cell Death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Fuentes-Prior, P.; Salvesen, G.S. The Protein Structures That Shape Caspase Activity, Specificity, Activation and Inhibition. Biochem. J. 2004, 384 Pt 2, 201–232. [Google Scholar] [CrossRef] [PubMed]
- Grutter, M.G. Caspases: Key Players in Programmed Cell Death. Curr. Opin. Struct. Biol. 2000, 10, 649–655. [Google Scholar] [CrossRef]
- Earnshaw, W.C.; Martins, L.M.; Kaufmann, S.H. Mammalian Caspases: Structure, Activation, Substrates, and Functions during Apoptosis. Annu. Rev. Biochem. 1999, 68, 383–424. [Google Scholar] [CrossRef] [PubMed]
- Fan, T.J.; Han, L.H.; Cong, R.S.; Liang, J. Caspase Family Proteases and Apoptosis. Acta Biochim. Biophys. Sin. 2005, 37, 719–727. [Google Scholar] [CrossRef]
- Bolkent, Ş.; Füsün, Ö.; Gezginci-Oktayoglu, S.; Oktayoğlu, S.; Karatuğ, A. A Matter of Regeneration and Repair: Caspases as the Key Molecules. Turk. J. Biol. 2016, 40, 333–352. [Google Scholar] [CrossRef]
- Brown-Suedel, A.N.; Bouchier-Hayes, L. Caspase-2 Substrates: To Apoptosis, Cell Cycle Control, and Beyond. Front. Cell Dev. Biol. 2020, 8, 610022. [Google Scholar] [CrossRef]
- Li, P.; Zhou, L.; Zhao, T.; Liu, X.; Zhang, P.; Liu, Y.; Zheng, X.; Li, Q. Caspase-9: Structure, Mechanisms and Clinical Application. Oncotarget 2017, 8, 23996–24008. [Google Scholar] [CrossRef]
- Cohen, G.M. Caspases: The Executioners of Apoptosis. Biochem. J. 1997, 326 Pt 1, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Boyce, M.; Degterev, A.; Yuan, J. Caspases: An Ancient Cellular Sword of Damocles. Cell Death Differ. 2004, 11, 29–37. [Google Scholar] [CrossRef]
- Boice, A.; Bouchier-Hayes, L. Targeting Apoptotic Caspases in Cancer. Biochim. Biophys. Acta Mol. Cell Res. 2020, 1867, 118688. [Google Scholar] [CrossRef] [PubMed]
- Murray, T.V.; McMahon, J.M.; Howley, B.A.; Stanley, A.; Ritter, T.; Mohr, A.; Zwacka, R.; Fearnhead, H.O. A Non-Apoptotic Role for Caspase-9 in Muscle Differentiation. J. Cell Sci. 2008, 121 Pt 22, 3786–3793. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Che, Z.; Li, C.; Zhang, W.; Zhao, X.; Guo, M. A Novel Caspase-1 Mediates Inflammatory Responses and Pyroptosis in Sea Cucumber Apostichopus japonicus. Aquaculture 2019, 513, 734399. [Google Scholar]
- Ye, S.; Gao, Y.; Wang, S.; Li, Q.; Li, R.; Li, H. Characterization and Expression Analysis of a Caspase-2 in an Invertebrate Echinoderm Sea Cumber Apostichopus japonicus. Fish Shellfish Immunol. 2016, 48, 266–272. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Li, C.; Zhang, W.; Duan, X.; Li, Y.; Jin, C.; Xiong, J.; Qiu, Q. Molecular Cloning and Characterization of Four Caspases Members in Apostichopus japonicus. Fish Shellfish Immunol. 2016, 55, 203–211. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree of Life (Itol) V5: An Online Tool for Phylogenetic Tree Display and Annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Zhao, Y.; Sun, L. Bacillus Cereus Cytotoxin K Triggers Gasdermin D-Dependent Pyroptosis. Cell Death Discov. 2022, 8, 305. [Google Scholar] [CrossRef]
- Xu, H.; Yuan, Z.; Qin, K.; Jiang, S.; Sun, L. The Molecular Mechanism and Evolutionary Divergence of Caspase 3/7-Regulated Gasdermin E Activation. eLife 2024, 12, RP89974. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Jiang, S.; Yu, C.; Yuan, Z.; Sun, L. Gsdmea-Mediated Pyroptosis Is Bi-Directionally Regulated by Caspase and Required for Effective Bacterial Clearance in Teleost. Cell Death Dis. 2022, 13, 491. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Yuan, Z.; Sun, L. A Non-Canonical Teleost Nk-Lysin: Antimicrobial Activity Via Multiple Mechanisms. Int. J. Mol. Sci. 2022, 23, 12722. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Sun, Y.; Sun, L. A Teleost Cxcl10 Is Both an Immunoregulator and an Antimicrobial. Front. Immunol. 2022, 13, 917697. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Sun, L.; Shao, Y.; Zhang, X.; Li, C. Ampk-Mediated Glutaminolysis Maintains Coelomocytes Redox Homeostasis in Vibrio Splendidus-Challenged Apostichopus japonicus. Fish Shellfish Immunol. 2022, 122, 170–180. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Zhou, Z.; Sun, Y.; Zhang, T.; Sun, L. Coral Gasdermin Triggers Pyroptosis. Sci. Immunol. 2020, 5, eabd2591. [Google Scholar] [CrossRef] [PubMed]
- Riedl, S.J.; Shi, Y. Molecular Mechanisms of Caspase Regulation During Apoptosis. Nat. Rev. Mol. Cell Biol. 2004, 5, 897–907. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Chen, T.; Wu, X.; Li, Z.; Zhang, X.; Jiang, X.; Luo, P.; Hu, C.; Wong, N.K.; Ren, C. Evolutionarily Ancient Caspase-9 Sensitizes Immune Effector Coelomocytes to Cadmium-Induced Cell Death in the Sea Cucumber, Holothuria leucospilota. Front. Immunol. 2022, 13, 927880. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Chen, J.; Cui, Y.; Li, S. Mir-2 Contributes to Wssv Infection by Targeting Caspase 2 in Mud Crab (Scylla paramamosain). Dev. Comp. Immunol. 2021, 125, 104217. [Google Scholar] [CrossRef]
- Nicholson, D.W.; Thornberry, N.A. Caspases: Killer Proteases. Trends Biochem. Sci. 1997, 22, 299–306. [Google Scholar] [CrossRef]
- Ando, K.; Kernan, J.L.; Liu, P.H.; Sanda, T.; Logette, E.; Tschopp, J.; Look, A.T.; Wang, J.; Bouchier-Hayes, L.; Sidi, S. Pidd Death-Domain Phosphorylation by Atm Controls Prodeath Versus Prosurvival Piddosome Signaling. Mol. Cell 2012, 47, 681–693. [Google Scholar] [CrossRef] [PubMed]
- Sidi, S.; Sanda, T.; Kennedy, R.D.; Hagen, A.T.; Jette, C.A.; Hoffmans, R.; Pascual, J.; Imamura, S.; Kishi, S.; Amatruda, J.F.; et al. Chk1 Suppresses a Caspase-2 Apoptotic Response to DNA Damage That Bypasses P53, Bcl-2, and Caspase-3. Cell 2008, 133, 864–877. [Google Scholar] [CrossRef]
- Fava, L.L.; Bock, F.J.; Geley, S.; Villunger, A. Caspase-2 at a Glance. J. Cell Sci. 2012, 125 Pt 24, 5911–5915. [Google Scholar] [CrossRef]
- Shi, J.; Zhao, Y.; Wang, Y.; Gao, W.; Ding, J.; Li, P.; Hu, L.; Shao, F. Inflammatory Caspases Are Innate Immune Receptors for Intracellular Lps. Nature 2014, 514, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Matikainen, S.; Nyman, T.A.; Cypryk, W. Function and Regulation of Noncanonical Caspase-4/5/11 Inflammasome. J. Immunol. 2020, 204, 3063–3069. [Google Scholar] [CrossRef]
- Lu, G.; Yu, Z.; Lu, M.; Liu, D.; Wang, F.; Wu, Y.; Liu, Y.; Liu, C.; Wang, L.; Song, L. The Self-Activation and Lps Binding Activity of Executioner Caspase-1 in Oyster Crassostrea Gigas. Dev. Comp. Immunol. 2017, 77, 330–339. [Google Scholar] [CrossRef]
- Uchiyama, R.; Tsutsui, H. Caspases as the Key Effectors of Inflammatory Responses against Bacterial Infection. Arch. Immunol. Ther. Exp. 2015, 63, 1–13. [Google Scholar] [CrossRef]
- Creagh, E.M.; Conroy, H.; Martin, S.J. Caspase-Activation Pathways in Apoptosis and Immunity. Immunol. Rev. 2003, 193, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Faherty, C.S.; Maurelli, A.T. Staying Alive: Bacterial Inhibition of Apoptosis During Infection. Trends Microbiol. 2008, 16, 173–180. [Google Scholar] [CrossRef]
- Liu, Y.X.; Zhou, D.Y.; Liu, Z.Q.; Lu, T.; Song, L.; Li, D.M.; Dong, X.P.; Qi, H.; Zhu, B.W.; Shahidi, F. Structural and Biochemical Changes in Dermis of Sea Cucumber (Stichopus japonicus) during Autolysis in Response to Cutting the Body Wall. Food Chem. 2018, 240, 1254–1261. [Google Scholar] [CrossRef]
- Qi, H.; Fu, H.; Dong, X.; Feng, D.; Li, N.; Wen, C.; Nakamura, Y.; Zhu, B. Apoptosis Induction Is Involved in Uva-Induced Autolysis in Sea Cucumber Stichopus japonicus. J. Photochem. Photobiol. B Biol. 2016, 158, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Gu, M.; Ma, H.; Mai, K.; Zhang, W.; Ai, Q.; Wang, X.; Bai, N. Immune Response of Sea Cucumber Apostichopus japonicus Coelomocytes to Several Immunostimulants In Vitro. Aquaculture 2010, 306, 49–56. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, H.; Yuan, Z.; Xu, H.; Sun, L. Characterization of the Apoptotic and Antimicrobial Activities of Two Initiator Caspases of Sea Cucumber Apostichopus japonicus. Genes 2024, 15, 540. https://doi.org/10.3390/genes15050540
Zhu H, Yuan Z, Xu H, Sun L. Characterization of the Apoptotic and Antimicrobial Activities of Two Initiator Caspases of Sea Cucumber Apostichopus japonicus. Genes. 2024; 15(5):540. https://doi.org/10.3390/genes15050540
Chicago/Turabian StyleZhu, Hanshuo, Zihao Yuan, Hang Xu, and Li Sun. 2024. "Characterization of the Apoptotic and Antimicrobial Activities of Two Initiator Caspases of Sea Cucumber Apostichopus japonicus" Genes 15, no. 5: 540. https://doi.org/10.3390/genes15050540
APA StyleZhu, H., Yuan, Z., Xu, H., & Sun, L. (2024). Characterization of the Apoptotic and Antimicrobial Activities of Two Initiator Caspases of Sea Cucumber Apostichopus japonicus. Genes, 15(5), 540. https://doi.org/10.3390/genes15050540