
The Satellite DNA PcH-Sat, Isolated and Characterized in the Limpet Patella caerulea (Mollusca, Gastropoda), Suggests the Origin from a Nin-SINE Transposable Element





Abstract
:1. Introduction
2. Material and Methods
2.1. DNA Extraction
2.2. Southern Blot
2.3. Quantitative Dot Blot
2.4. Genome Size Estimation
2.5. Fluorescence In Situ Hybridization (FISH)
2.6. Bioinformatic Analysis
3. Results
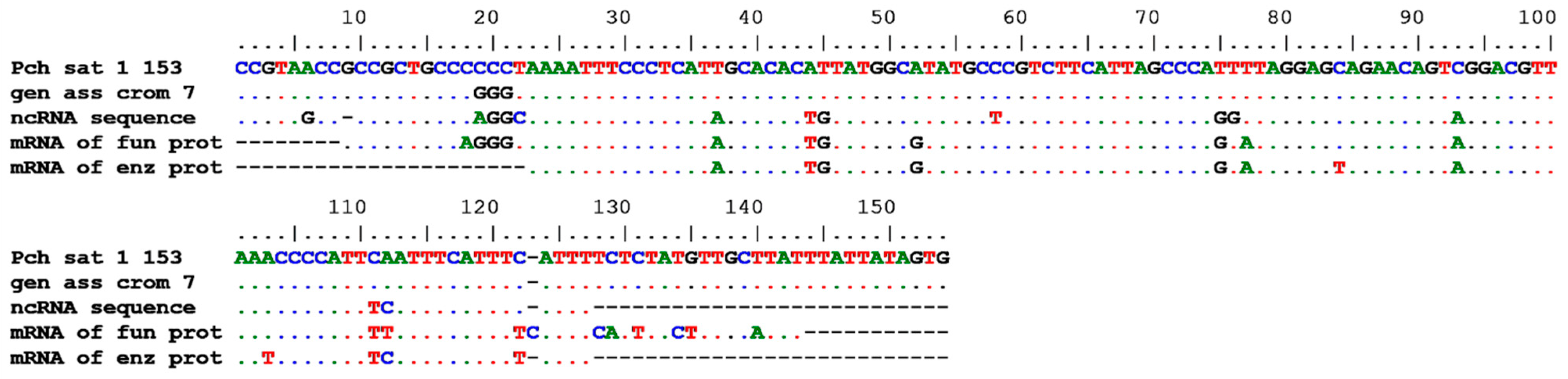
3.1. Restriction Enzyme and Sequence Analysis
3.2. Southern Blot
3.3. Quantitative Dot Blot
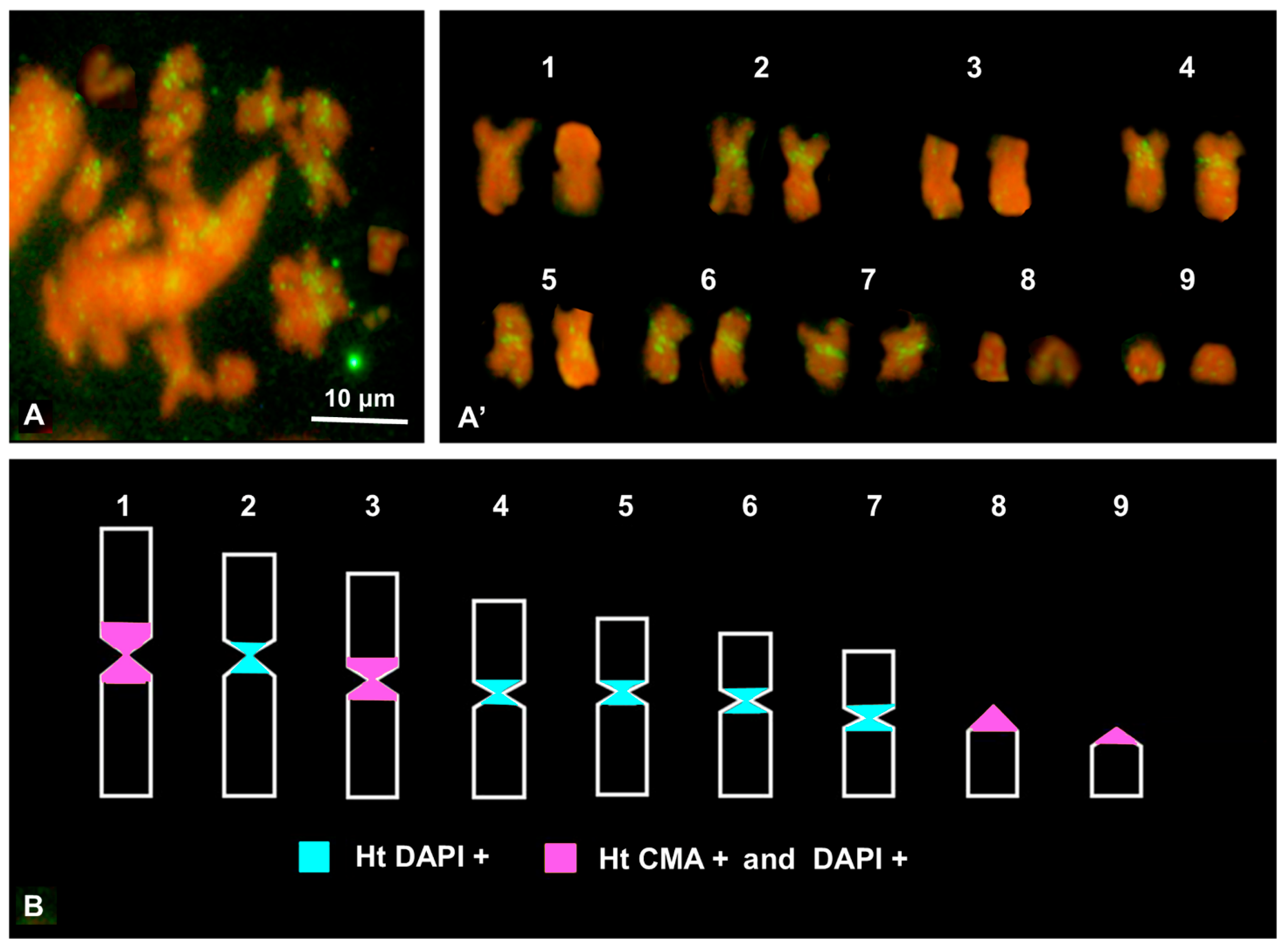
3.4. FISH
3.5. Repeatmasker and Repbase Queries
3.6. BLAST Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WoRMS. Editorial Board. World Register of Marine Species. Available online: https://www.marinespecies.org (accessed on 14 March 2023).
- Petraccioli, A.; Guarino, F.M.; Maio, N.; Odierna, G. Molecular cytogenetic study of three common Mediterranean limpets, Patella caerulea, P. rustica and P. ulyssiponensis, Archaeogastropoda, Mollusca. Genetica 2010, 138, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Cascales-Soler, A.; Ramos-Espla, A.A.; Forcada, A. Reviewing the knowledge on the genus Patella in the Mediterranean Sea: Testing the effect of protection on the mean abundance and size of Patella ferruginea. Front. Mar. Sci. 2023, 10, 158470. [Google Scholar] [CrossRef]
- Guallart, J.; Templado, J. Patella ferruginea. En: VV.AA., Bases ecológicas preliminares para la conservación de las especies de interés comunitario en España: Invertebrados. In Ases Ecológicas Preliminares para la Conservación de las Especies de Interés Comunitario en España: Invertebrados; Ministerio de Agricultura, Alimentación y Medio Ambiente: Madrid, Spain, 2012; p. 86. [Google Scholar]
- UNEP/MAP; SPA/RAC. Annex II: List of Endangered or Threatened Species; SPA-BD Protocol; SAP/RAC: Tunis, Tunisia, 2018. [Google Scholar]
- Guerra-García, J.M.; Corzo, J.; Espinosa, F.; García-Gómez, J.C. Assessing habitat use of the endangered marine mollusc Patella ferruginea (Gastropoda, Patellidae) in northern Africa: Preliminary results and implications for conservation. Biol. Conserv. 2004, 116, 319–326. [Google Scholar] [CrossRef]
- Bresler, V.; Abelson, A.; Fishelson, L.; Feldestein, T.; Rosenfeld, M.; Mokady, O. Marine molluscs in environmental monitoring. I. Cellular and molecular responses. Helgol. Mar. Res. 2003, 57, 157–165. [Google Scholar] [CrossRef]
- Cravo, A.; Bebianno, M.J. Bioaccumulation of metals in the soft tissue of Patella aspera: Application of metal/shell weight indices. Estuar. Coast. Mar. Sci. 2005, 65, 571–586. [Google Scholar] [CrossRef]
- Pérez, S.; Sánchez-Marín, P.; Bellasc, J.; Viñas, L.; Besada, V.; Fernández, N. Limpets (Patella spp. Mollusca, Gastropoda) as model organisms for biomonitoring environmental quality. Ecol. Indic. 2019, 101, 150–162. [Google Scholar] [CrossRef]
- Koufopanou, V.; Reid, D.G.; Ridgway, S.A.; Thomas, R.H. A molecular phylogeny of the patellid limpets (Gastropoda: Patellidae) and its implications for the origins of their antitropical distribution. Mol. Phylogenet. Evol. 1999, 11, 138–156. [Google Scholar] [CrossRef]
- Sá-Pinto, A.; Branco, M.; Harris, D.-J.; Alexandrino, P. Phylogeny and phylogeography of the genus Patella based on mitochondrial DNA sequence data. J. Exp. Mar. Biol. Ecol. 2005, 325, 95–110. [Google Scholar] [CrossRef]
- Sá-Pinto, A.; Branco, M.; Sayanda, D.; Alexandrino, P. Patterns of colonization, evolution and gene flow in species of the genus Patella in the Macaronesian Islands. Mol. Ecol. 2008, 17, 519–532. [Google Scholar] [CrossRef]
- Casu, M.; Casu, D.; Lai, T.; Cossu, P.; Curini-Galletti, M. Inter-simple sequence repeat markers reveal strong genetic differentiation among populations of the endangered mollusc Patella ferruginea (Gastropoda: Patellidae) from two Sardinian marine protected areas. Mar. Biol. 2006, 149, 1163–1174. [Google Scholar] [CrossRef]
- Casu, M.; Rivera-Ingraham, G.A.; Cossu, P.; Lai, T.; Sanna, D.; Dedola, G.L.; Sussarellu, R.; Sella, G.; Cristo, B.; Curini-Galletti, M.; et al. Patterns of spatial genetic structuring in the endangered limpet Patella ferruginea: Implications for the conservation of a Mediterranean endemic. Genetica 2011, 139, 1293–1308. [Google Scholar] [CrossRef] [PubMed]
- Faria, J.; Pita, A.; Rivas, M.; Martins, G.M.; Hawkins, S.J.; Ribeiro, P.; Neto, A.I.; Presa, P. A multiplex microsatellite tool for conservation genetics of the endemic limpet Patella candei in the Macaronesian archipelagos. Aquat. Cons. 2016, 26, 775–781. [Google Scholar] [CrossRef]
- Quinteiro, J.; ·González-Lorenzo, G.; Hernández-Reyes, D.; Quinteiro, L.; Herrera-Pérez, R.; Martínez-Barrio, J.; González, J.-A.; Rey-Méndez, M.; González-Henríquez, N. Genetic differentiation of a critically endangered population of the limpet Patella candei candei d’Orbigny, 1840, in the Canary Islands. Conserv. Genet. 2022, 23, 807–819. [Google Scholar] [CrossRef]
- Simakov, O.; Marletaz, F.; Cho, S.J.; Edsinger-Gonzales, E.; Havlak, P.; Hellsten, U.; Kuo, D.H.; Larsson, T.; Lv, J.; Arendt, D.; et al. Insights into bilaterian evolution from three spiralian genomes. Nature 2012, 493, 526–531. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chu, J.; Cheng, H.; Li, H. De novo reconstruction of satellite repeat units from sequence data. Genome Res. 2023, 33, 1994–2001. [Google Scholar] [CrossRef] [PubMed]
- Cervella, P.; Ramella, L.; Robotti, C.A.; Sella, G. Chromosome analysis of three species of Patella (Archaeogastropoda). Genetica 1988, 77, 97–103. [Google Scholar] [CrossRef]
- John, B.E.; Miklos, G.L.G. Functional aspects of satellite DNA and heterochromatin. Int. Rev. Cytol. 1979, 58, 1–114. [Google Scholar] [CrossRef]
- Singer, M.F. Highly repeated sequences in mammalian genomes. Int. Rev. Cytol. 1982, 67, 67–112. [Google Scholar] [CrossRef] [PubMed]
- Mezzasalma, M.; Andreone, F.; Glaw, F.; Guarino, F.M.; Odierna, G.; Petraccioli, A.; Picariello, O. Changes in heterochromatin content and ancient chromosome fusion in the endemic Malagasy boid snakes Sanzinia and Acrantophis (Squamata: Serpentes). Salamandra 2019, 55, 140–144. [Google Scholar]
- Šatović-Vukšić, E.; Plohl, M. Satellite DNAs—From localized to highly dispersed genome components. Genes 2023, 14, 742. [Google Scholar] [CrossRef]
- Miklos, G.L.G. Localized highly repetitive DNA sequences in vertebrate and invertebrate genomes. In Molecular Evolutionary Genetics; Maclntyre, R.J., Ed.; Plenum Press: New York, NY, USA, 1985; pp. 241–321. [Google Scholar]
- Redi, C.A.; Garagna, S.; Zacharias, H.; Zuccotti, M.; Capanna, E. The other chromatin. Chromosoma 2001, 110, 136–147. [Google Scholar] [CrossRef] [PubMed]
- Feliciello, I.; Pezer, Ž.; Kordiš, D.; Bruvo Mađarić, B.; Urđica, Đ. Evolutionary History of Alpha Satellite DNA Repeats Dispersed within human genome euchromatin. Genome Biol. Evol. 2020, 12, 2125–2138. [Google Scholar] [CrossRef] [PubMed]
- Altemose, N.; Logsdon, G.-A.; Bzikadze, A.V.; Sidhwani, P.; Langley, S.A.; Caldas, G.V.; Hoyt, S.J.; Uralsky, L.; Ryabov, F.D.; Shew, C.J.; et al. Complete genomic and epigenetic maps of human centromeres. Science 2022, 376, eabl4178. [Google Scholar] [CrossRef] [PubMed]
- Doolittle, W.F.; Sapienza, C. Selfish genes, the phenotype paradigm and genome evolution. Nature 1980, 284, 601–603. [Google Scholar] [CrossRef] [PubMed]
- Orgel, L.E.; Crick, F.H.C. Selfish DNA: The ultimate parasite. Nature 1980, 284, 604–607. [Google Scholar] [CrossRef] [PubMed]
- Phillippy, A.M. New advances in sequence assembly. Genome Res. 2017, 27, xi–xiii. [Google Scholar] [CrossRef] [PubMed]
- Lower, S.S.; McGurk, M.P.; Clark, A.G.; Barbash, D.A. Satellite DNA evolution: Old ideas, new approaches. Curr. Opin. Genet. Dev. 2018, 49, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Sedlazeck, F.J.; Lee, H.; Darby, C.A.; Schatz, M.C. Piercing the Dark Matter: Bioinformatics of Long-Range Sequencing and Mapping. Nat. Rev. Genet. 2018, 19, 329–346. [Google Scholar] [CrossRef] [PubMed]
- Tunjić Cvitanić, M.; Vojvoda Zeljko, T.; Pasantes, J.J.; García-Souto, D.; Gržan, T.; Despot-Slade, E.; Plohl, M.; Šatović, E. Sequence composition underlying centromeric and heterochromatic genome compartments of the Pacific Oyster Crassostrea gigas. Genes 2020, 11, 695. [Google Scholar] [CrossRef]
- Tunjić Cvitanić, M.; Pasantes, J.J.; García-Souto, D.; Cvitanić, T.; Plohl, M.; Šatović -Vukšić, E. Satellitome Analysis of the Pacific Oyster Crassostrea gigas reveals new pattern of Satellite DNA organization, highly scattered across the Genome. Int. J. Mol. Sci. 2021, 22, 6798. [Google Scholar] [CrossRef]
- Kunyavskaya, O.; Dvorkina, T.; Bzikadze, A.V.; Alexandrov, I.A.; Pevzner, P.A. Automated annotation of human centromeres with HORmon. Genome Res. 2022, 32, 1137–1151. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Yang, X.; Guo, H.; Zhao, X.; Wang, B.; Ye, K. HiCAT: A tool for automatic annotation of centromere structure. Genome Biol. 2023, 24, 58. [Google Scholar] [CrossRef]
- Pech, M.; Igo-Kemenes, T.; Zachau, H.G. Nucleotide sequence of a highly repetitive component of rat DNA. Nucleic Acids Res. 1979, 7, 417–432. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Ramos, M.A. Satellite DNA: An evolving topic. Genes 2017, 8, 230. [Google Scholar] [CrossRef]
- Petraccioli, A.; Odierna, G.; Capriglione, T.; Barucca, M.; Forconi, M.; Olmo, E.; Biscotti, M.A. A novel satellite DNA isolated in Pecten jacobaeus shows high sequence similarity among molluscs. Mol. Genet. Genom. 2015, 290, 1717–1725. [Google Scholar] [CrossRef]
- Sokolov, E.P. An improved method for DNA isolation from mucopolysaccharide-rich molluscan tissues. J. Mollusc. Stud. 2000, 66, 573–575. [Google Scholar] [CrossRef]
- Picariello, O.; Odierna, G.; Petraccioli, A.; Amor, N.; Feliciello, I.; Chinali, G. Characterization of two major satellite DNAs specific to the genus Discoglossus (Amphibia, Anura). Ital. J. Zool. 2012, 79, 385–394. [Google Scholar] [CrossRef]
- Hardie, D.C.; Gregory, T.R.; Hebert, P.D.N. From pixels to picograms, a beginners’ guide to genome quantification by Feulgen image analysis densitometry. J. Histochem. Cytochem. 2002, 50, 735–749. [Google Scholar] [CrossRef]
- Gregory, T.R. Animal Genome Size Database. Available online: http://www.genomesize.com (accessed on 14 March 2023).
- Mezzasalma, M.; Andreone, F.; Aprea, G.; Glaw, F.; Odierna, G.; Guarino, F.M. When can chromosomes drive speciation? The peculiar case of the Malagasy tomato frogs (genus Dyscophus). Zool. Anz. 2017, 268, 41–46. [Google Scholar] [CrossRef]
- Bao, W.; Kojima, K.K.; Kohany, O. Repbase Update, a database of repetitive elements in eukaryotic genomes. Mobile DNA 2015, 6, 11. [Google Scholar] [CrossRef]
- Smit, A.F.A.; Hubley, R.; Green, P. RepeatMasker Open-4.0. 2013–2015. Available online: http://www.repeatmasker.org (accessed on 14 March 2023).
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 2020, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Kenny, N.J.; Namigai, E.K.O.; Marlétaz, F.; Hui, J.H.L.; Shimeld, S.M. Draft genome assemblies and predicted microRNA complements of the intertidal lophotrochozoans Patella vulgata, Mollusca, Patellogastropoda and Spirobranchus (Pomatoceros) lamarcki. Annelida, Serpulida. Mar. Genom. 2015, 24, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Piskurek, O.; Jackson, D.J. Tracking the ancestry of a deeply conserved eumetazoan SINE domain. Mol. Biol. Evol. 2011, 28, 2727–2730. [Google Scholar] [CrossRef] [PubMed]
- Halstead-Nussloch, G.; Signorini, S.G.; Giulio, M.; Crocetta, F.; Munari, M.; Della Torre, C.; Weber, A.A.-T. The genome of the rayed Mediterranean limpet Patella caerulea (Linnaeus, 1758). Genome Biol. Evol. 2024, 1, evae070. [Google Scholar] [CrossRef] [PubMed]
- Šatović, E.; Vojvoda Zeljko, T.; Plohl, M. Characteristics and evolution of satellite DNA sequences in bivalve mollusks. Eur. Zool. J. 2018, 85, 94–103. [Google Scholar] [CrossRef]
- Markova, N.G.; Markov, G.G. Complex organization of a cryptic satellite DNA in the genome of the marine invertebrate Rapana thomasiana Grosse (Gastropoda). Biochim. Biophys. Acta 1983, 741, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Biscotti, M.A.; Canapa, A.; Olmo, E.; Barucca, M.; Teo, C.H.; Schwarzacher, T.; Dennerlein, S.; Richter, R.; Heslop-Harrison, J.S. Repetitive DNA, molecular cytogenetics and genome organization in the king scallop (Pecten maximus). Gene 2007, 406, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Levitsky, V.G.; Babenko, V.N.; Vershinin, A.V. The roles of the monomer length and nucleotide context of plant tandem repeats in nucleosome positioning. J. Biomol. Struct. Dynam. 2014, 32, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, Z.; Guo, X. A Centromeric satellite sequence in the Pacific Oyster (Crassostrea gigas Thunberg) Identified by Fluorescence In Situ Hybridization. Mar. Biotechnol. 2001, 3, 486–492. [Google Scholar] [CrossRef] [PubMed]
- Piskurek, O.; Austin, C.C.; Okada, N. Sauria SINEs: Novel short interspersed retroposable elements that are widespread in reptile genomes. J. Mol. Evol. 2006, 62, 630–644. [Google Scholar] [CrossRef]
- Luchetti, A.; Šatović, E.; Mantovani, B.; Plohl, M. RUDI, a short interspersed element of the V-SINE superfamily widespread in molluscan genomes. Mol. Genet. Genom. 2016, 291, 1419–1429. [Google Scholar] [CrossRef] [PubMed]
- Petraccioli, A.; Guarino, F.M.; Kupriyanova, L.; Mezzasalma, M.; Odierna, G.; Picariello, O.; Capriglione, T. Isolation and characterization of interspersed repeated sequences in the Common Lizard, Zootoca vivipara, and their conservation in Squamata. Cytogenet. Genome Res. 2019, 157, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Paço, A.; Freitas, R.; Vieira-da-Silva, A. Conversion of DNA sequences: From a transposable element to a tandem repeat or to a gene. Genes 2019, 10, 1014. [Google Scholar] [CrossRef] [PubMed]
- McGurk, M.P.; Barbash, D.A. Double insertion of transposable elements provides a substrate for the evolution of satellite DNA. Genome Res. 2018, 28, 714–725. [Google Scholar] [CrossRef] [PubMed]
- Tollis, M.; Boissinot, S. The transposable element profile of the Anolis genome: How a lizard can provide insights into the evolution of vertebrate genome size and structure. Mob. Genet. Elem. 2011, 1, 107–111. [Google Scholar] [CrossRef]
- Mezzasalma, M.; Visone, V.; Petraccioli, A.; Guarino, F.M.; Capriglione, T.; Odierna, G. Non-random accumulation of LINE1 like sequences on differentiated snake W chromosomes. J. Zool. 2016, 300, 65–75. [Google Scholar] [CrossRef]
- Mezzasalma, M.; Capriglione, T.; Kupriyanova, L.; Odierna, G.; Pallotta, M.M.; Petraccioli, A.; Picariello, O.; Guarino, F.M. Characterization of two Transposable Elements and an Ultra-Conserved Element isolated in the genome of Zootoca vivipara (Squamata, Lacertidae). Life 2023, 13, 637. [Google Scholar] [CrossRef]
- Jensen, S.; Gassama, M.P.; Heidmann, T. Taming of transposable elements by homology dependent gene silencing. Nat. Genet. 1999, 21, 200–212. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Locality | Number |
|---|---|---|
| P. caerulea | Pozzuoli, Naples, Italy | 3 |
| Gaeta, Latina, Italy | 2 | |
| P. rustica | Nisida, Naples, Italy | 2 |
| Acciaroli, Salerno, Italy | 1 | |
| P. ulyssiponensis | Gaiola, Naples, Italy | 2 |
| P. vulgata | Baiona, Spain | 2 |
| Porto, Portugal | 1 | |
| P. depressa | Porto, Portugal | 2 |
| P. ferruginea | Porticcio, Corse, France | 1 |
| Species | Nr. Hits | Max Score | % Cover | E Value | % Identity | Orient. | Accession | |
|---|---|---|---|---|---|---|---|---|
| P. caerulea | 29 | 1st hit | 285 | 98% | 3.00 × 10−75 | 94129 | +/− | JAZGQO010000021 |
| Last hits | 233 | 93% | 2.00 × 10−59 | 93.092 | +/− | JAZGQO010000031 | ||
| P. depressa | 340 | 1st hit | 260 | 100% | 3.00 × 10−68 | 97.39% | +/+ | CAOLEZ010000010 |
| Last hit | 81.5 | 72% | 6.00 × 10−14 | 76.58% | +/+ | CAOLEW010000395 | ||
| P. pellucida | 726 | 1st hit | 233 | 99% | 1.00 × 10−59 | 94.08% | +/− | CAKJPO010001416 CAKJPO010001283 |
| Last hit | 59.9 | 70% | 2.00 × 10−7 | 73.08% | +/− | |||
| P. ulyssiponensis | 557 | 1st hit | 279 | 100% | 2.00 × 10−71 | 99.35% | +/− | CAVMBP010000854 |
| Last hit | 85.1 | 70% | 5.00 × 10−15 | 79.59 | +/+ | CAVMBP010000752 | ||
| P. vulgata | 624 | 1st hit | 166 | 75% | 2.00 × 10−39 | 88.49% | +/− | CAKNZQ020000656 |
| Last hit | 88.7 | 75% | 1.00 × 10−15 | 74.50% | +/− | CAKNZQ020001418 | ||
| L. scabra | 111 | 1st hit | 126 | 89% | 2.00 × 10−27 | 85.59% | +/− | JARJEJ010000003 |
| Last hit | 94.2 | 78% | 1.00 × 10−17 | 81.90% | +/+ | JARJEJ010000118 | ||
| L. gigantea | 113 | 1st hit | 113 | 70% | 4.00 × 10−24 | 81.89% | +/+ | AMQO01003418 |
| Last hit | 63.5 | 78% | 6.00 × 10−9 | 71.33% | +/− | AMQO01000500 | ||
| N. schrenckii | 16 | 1st hit | 129 | 86% | 7.00 × 10−29 | 86.84% | +/− | JAUJPP010000003 |
| Last hit | 68.0 | 72% | 2.00 × 10−10 | 78.72% | +/− | JAUJPP010000958 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petraccioli, A.; Maio, N.; Carotenuto, R.; Odierna, G.; Guarino, F.M. The Satellite DNA PcH-Sat, Isolated and Characterized in the Limpet Patella caerulea (Mollusca, Gastropoda), Suggests the Origin from a Nin-SINE Transposable Element. Genes 2024, 15, 541. https://doi.org/10.3390/genes15050541
Petraccioli A, Maio N, Carotenuto R, Odierna G, Guarino FM. The Satellite DNA PcH-Sat, Isolated and Characterized in the Limpet Patella caerulea (Mollusca, Gastropoda), Suggests the Origin from a Nin-SINE Transposable Element. Genes. 2024; 15(5):541. https://doi.org/10.3390/genes15050541
Chicago/Turabian StylePetraccioli, Agnese, Nicola Maio, Rosa Carotenuto, Gaetano Odierna, and Fabio Maria Guarino. 2024. "The Satellite DNA PcH-Sat, Isolated and Characterized in the Limpet Patella caerulea (Mollusca, Gastropoda), Suggests the Origin from a Nin-SINE Transposable Element" Genes 15, no. 5: 541. https://doi.org/10.3390/genes15050541
APA StylePetraccioli, A., Maio, N., Carotenuto, R., Odierna, G., & Guarino, F. M. (2024). The Satellite DNA PcH-Sat, Isolated and Characterized in the Limpet Patella caerulea (Mollusca, Gastropoda), Suggests the Origin from a Nin-SINE Transposable Element. Genes, 15(5), 541. https://doi.org/10.3390/genes15050541