
GhCLCc-1, a Chloride Channel Gene from Upland Cotton, Positively Regulates Salt Tolerance by Modulating the Accumulation of Chloride Ions

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Treatments
2.2. Cloning and Sequence Analyses
2.3. Quantitative Real-Time PCR (qRT-PCR)
2.4. Subcellular Localization of GhCLCc-1A/D
2.5. Genetic Transformation
2.6. Virus-Induced Silencing (VIGS) of GhCLCc-1 in Upland Cotton
2.7. Biochemical Index Measurement
2.8. Analysis of Ion Contents
3. Results
3.1. GhCLCc-1 Transcript Abundance Is Induced by Chloride Salt Stress
3.2. GhCLCc-1 Localizes to the Tonoplast
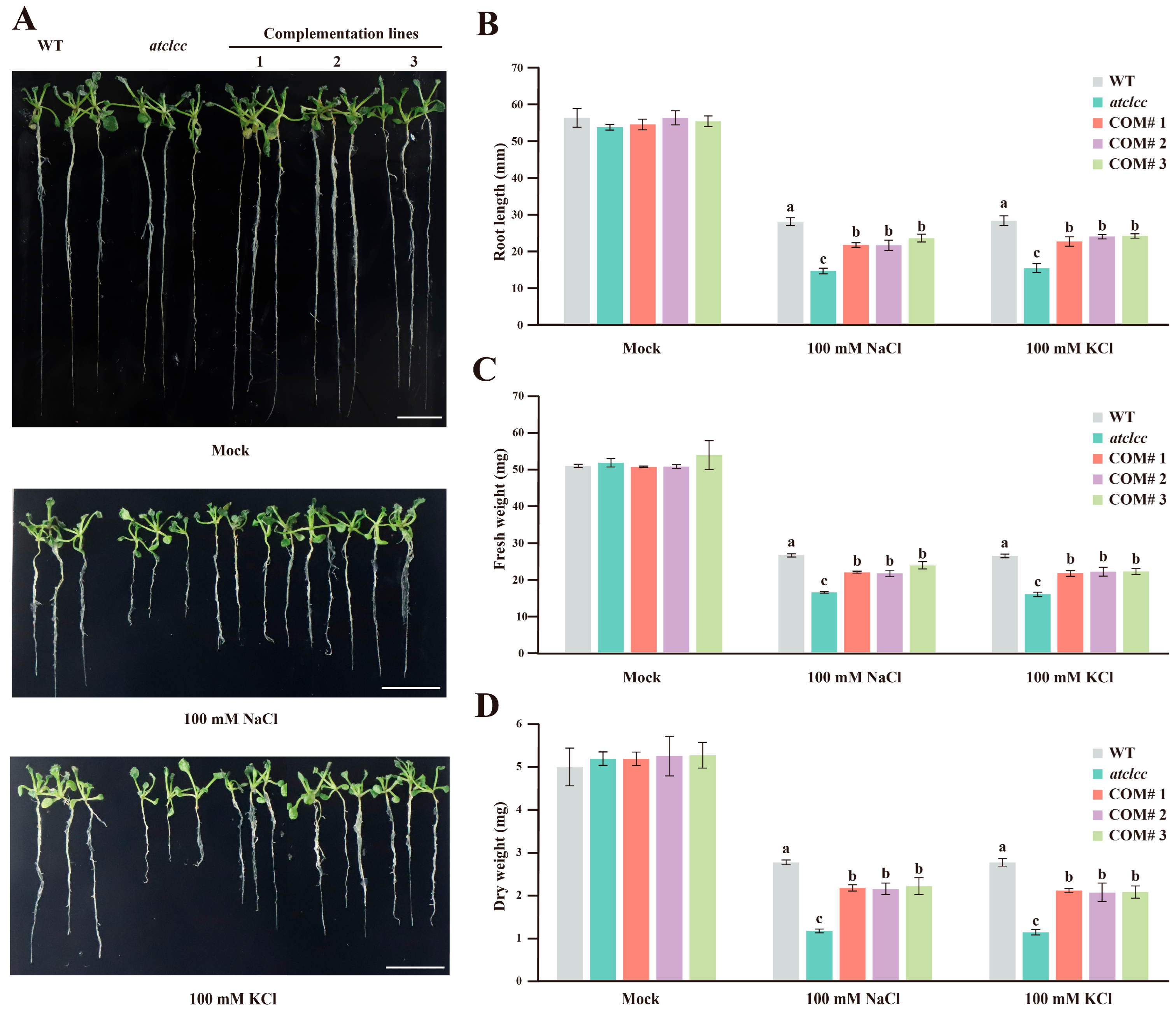
3.3. GhCLCc-1 Rescues Chlorine Salt Tolerance in Arabidopsis Mutant Atclcc
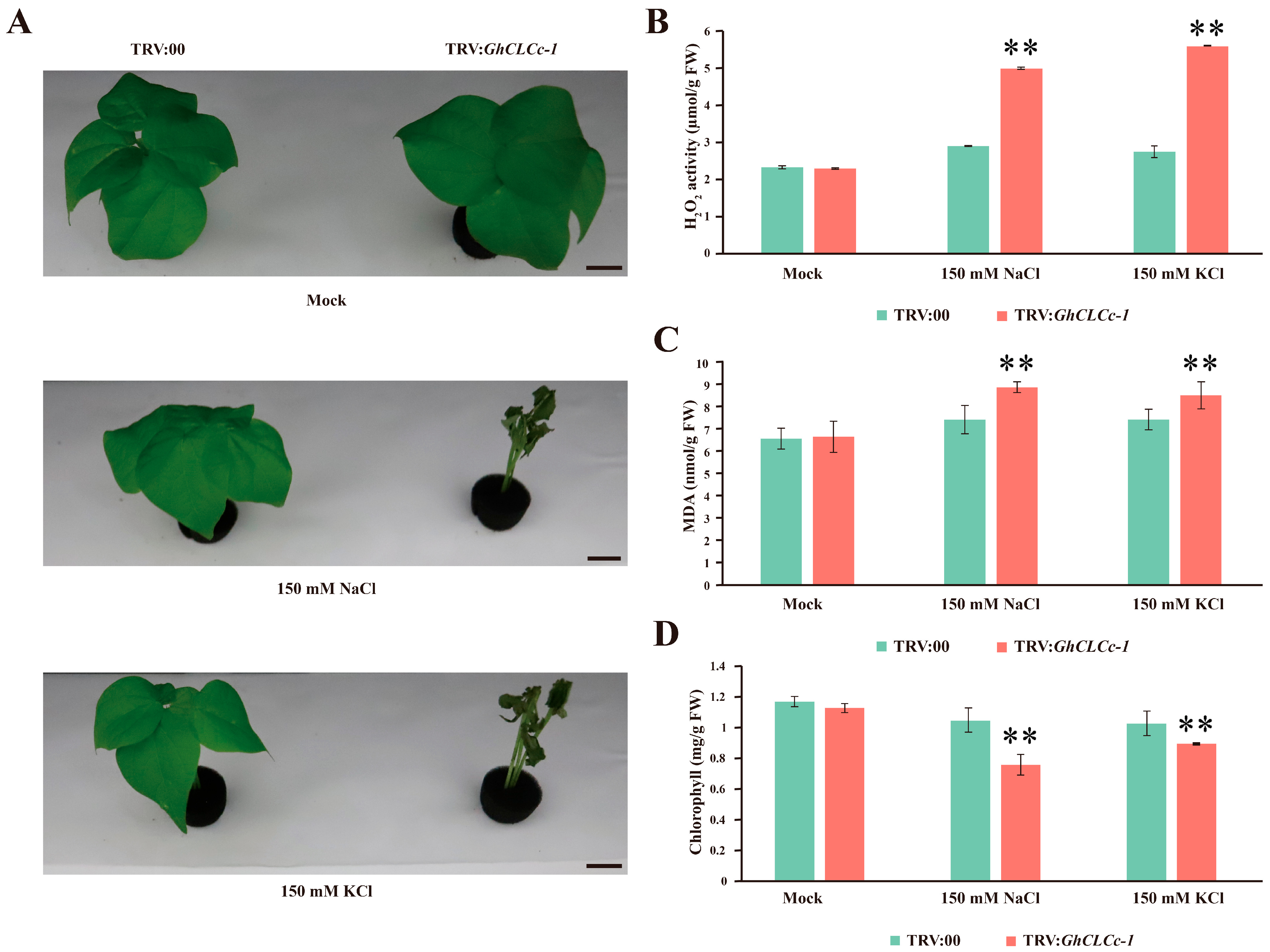
3.4. Silencing GhCLCc-1 in Cotton Reduces Chlorine Salt Tolerance
3.5. GhCLCc-1 Ectopic Expression Enhances Chloride Salt Tolerance in Arabidopsis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Balasubramaniam, T.; Shen, G.; Esmaeili, N.; Zhang, H. Plants’ response mechanisms to salinity stress. Plants 2023, 12, 2253. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, M.S.; Sami, S.; Bano, M.; Rukh, M.; Khan, Q.; Anwar, Z.; Ijaz, A.; Ahmed, T. Impact of salt stress on cotton. Int. J. Agric. Biosci. 2023, 12, 98–103. [Google Scholar]
- dos Santos, T.B.; Ribas, A.F.; de Souza, S.G.H.; Budzinski, I.G.F.; Domingues, D.S. Physiological responses to drought, salinity, and heat stress in plants: A review. Stresses 2022, 2, 113–135. [Google Scholar] [CrossRef]
- Kaushal, S.S. Increased salinization decreases safe drinking water. Environ. Sci. Technol. 2016, 50, 2765–2766. [Google Scholar] [CrossRef] [PubMed]
- Kenzhebayeva, A.; Bakbolat, B.; Sultanov, F.; Daulbayev, C.; Mansurov, Z. A mini-review on recent developments in anti-icing methods. Polymers 2021, 13, 4149. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.; Chen, P.; Yu, S. The cytochrome P450 gene GhCYP94C1 is involved in drought stress in upland cotton (Gossypium hirsutum L.). Czech J. Genet. Plant Breed. 2023, 59, 189–195. [Google Scholar] [CrossRef]
- Wang, C.; Yang, Y.; Wang, H.; Ran, X.; Li, B.; Zhang, J.; Zhang, H. Ectopic expression of a cytochrome P450 monooxygenase gene PtCYP714A3 from Populus trichocarpa reduces shoot growth and improves tolerance to salt stress in transgenic rice. Plant Biotechnol. J. 2016, 14, 1838–1851. [Google Scholar] [CrossRef]
- Wang, X.; Wei, C.; Huang, H.; Kang, J.; Long, R.; Chen, L.; Li, M.; Yang, Q. The GARP family transcription factor MtHHO3 negatively regulates salt tolerance in Medicago truncatula. Plant Physiol. Biochem. 2024, 209, 108542. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Xu, D.; Zhou, L.; Chen, H.; Peng, Z.; Chen, G.; Wang, L.; Cao, H.; Peng, Y.; Geng, S.; et al. The critical role of CmCIPK1-CmRbohD1/D2 complexes in generating H2O2 signals for enhancing salt tolerance in pumpkins. Hortic. Plant J. 2024. [Google Scholar] [CrossRef]
- Du, L.; Ding, L.; Huang, X.; Tang, D.; Chen, B.; Tian, H.; Kang, Z.; Mao, H. Natural variation in a K(+)-preferring HKT transporter contributes to wheat shoot K(+) accumulation and salt tolerance. Plant Cell Environ. 2024, 47, 540–556. [Google Scholar] [CrossRef]
- Zhu, J.K. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef]
- Deinlein, U.; Stephan, A.B.; Horie, T.; Luo, W.; Xu, G.; Schroeder, J.I. Plant salt-tolerance mechanisms. Trends Plant Sci. 2014, 19, 371–379. [Google Scholar] [CrossRef]
- van Zelm, E.; Zhang, Y.; Testerink, C. Salt tolerance mechanisms of plants. Annu. Rev. Plant Biol. 2020, 71, 403–433. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Zhang, Q.; Liu, M.; Zhou, H.; Ma, C.; Wang, P. Regulation of plant responses to salt stress. Int. J. Mol. Sci. 2021, 22, 4609. [Google Scholar] [CrossRef]
- Zhou, X.; Li, J.; Wang, Y.; Liang, X.; Zhang, M.; Lu, M.; Guo, Y.; Qin, F.; Jiang, C. The classical SOS pathway confers natural variation of salt tolerance in maize. New Phytol. 2022, 236, 479–494. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhou, J.; Ni, X.; Wang, Q.; Jia, Y.; Xu, X.; Wu, H.; Fu, P.; Wen, H.; Guo, Y.; et al. Structural basis for the activity regulation of Salt Overly Sensitive 1 in Arabidopsis salt tolerance. Nat. Plants 2023, 9, 1915–1923. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.Y.; Tang, L.H.; Nie, J.W.; Zhang, C.R.; Han, X.; Li, Q.Y.; Qin, L.; Wang, M.H.; Huang, X.; Yu, F.; et al. Structure and activation mechanism of the rice Salt Overly Sensitive 1 (SOS1) Na+/H+ antiporter. Nat. Plants 2023, 9, 1924–1936. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Wang, X.; Zhang, Z.; Yang, L.; Zhang, Q.; Li, Y.; He, H.; Chen, D.; Zhang, B.; Zheng, C.; et al. Uncovering key salt-tolerant regulators through a combined eQTL and GWAS analysis using the super pan-genome in rice. Natl. Sci. Rev. 2024, 11, nwae043. [Google Scholar] [CrossRef]
- Geilfus, C.M. Chloride: From nutrient to toxicant. Plant Cell Physiol. 2018, 59, 877–886. [Google Scholar] [CrossRef]
- Tavakkoli, E.; Fatehi, F.; Coventry, S.; Rengasamy, P.; McDonald, G.K. Additive effects of Na+ and Cl− ions on barley growth under salinity stress. J. Exp. Bot. 2011, 62, 2189–2203. [Google Scholar] [CrossRef]
- Wu, H.; Li, Z. The importance of Cl− exclusion and vacuolar Cl− sequestration: Revisiting the role of Cl− transport in plant salt tolerance. Front. Plant Sci. 2019, 10, 1418. [Google Scholar] [CrossRef] [PubMed]
- Henderson, S.W.; Baumann, U.; Blackmore, D.H.; Walker, A.R.; Walker, R.R.; Gilliham, M. Shoot chloride exclusion and salt tolerance in grapevine is associated with differential ion transporter expression in roots. BMC Plant Biol. 2014, 14, 273. [Google Scholar] [CrossRef]
- Yin, P.; Liang, X.; Zhao, H.; Xu, Z.; Chen, L.; Yang, X.; Qin, F.; Zhang, J.; Jiang, C. Cytokinin signaling promotes salt tolerance by modulating shoot chloride exclusion in maize. Mol. Plant 2023, 16, 1031–1047. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, C.T.; Agorio, A.; Jossier, M.; Depré, S.; Thomine, S.; Filleur, S. Characterization of the chloride channel-like, AtCLCg, involved in chloride tolerance in Arabidopsis thaliana. Plant Cell Physiol. 2016, 57, 764–775. [Google Scholar] [CrossRef] [PubMed]
- Nedelyaeva, O.I.; Shuvalov, A.V.; Balnokin, Y.V. Chloride channels and transporters of the CLC family in plants. Russ. J. Plant Physiol. 2020, 67, 767–784. [Google Scholar] [CrossRef]
- Lv, Q.; Tang, R.; Liu, H.; Gao, X.; Li, Y.; Zheng, H.; Zhang, H. Cloning and molecular analyses of the Arabidopsis thaliana chloride channel gene family. Plant Sci. 2009, 176, 650–661. [Google Scholar] [CrossRef]
- Jossier, M.; Kroniewicz, L.; Dalmas, F.; Le Thiec, D.; Ephritikhine, G.; Thomine, S.; Barbier-Brygoo, H.; Vavasseur, A.; Filleur, S.; Leonhardt, N. The Arabidopsis vacuolar anion transporter, AtCLCc, is involved in the regulation of stomatal movements and contributes to salt tolerance. Plant J. 2010, 64, 563–576. [Google Scholar] [CrossRef] [PubMed]
- Wei, P.; Che, B.; Shen, L.; Cui, Y.; Wu, S.; Cheng, C.; Liu, F.; Li, M.W.; Yu, B.; Lam, H.M. Identification and functional characterization of the chloride channel gene, GsCLC-c2 from wild soybean. BMC Plant Biol. 2019, 19, 121. [Google Scholar] [CrossRef]
- Song, J.; Han, M.; Zhu, X.; Li, H.; Ning, Y.; Zhang, W.; Yang, H. MhCLC-c1, a Cl channel c homolog from Malus hupehensis, alleviates NaCl-induced cell death by inhibiting intracellular Cl− accumulation. BMC Plant Biol. 2023, 23, 306. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.; Liu, Y.; Zhou, G.; Li, Q.; Yang, C.; Peng, S. Overexpression of CsCLCc, a chloride channel gene from Poncirus trifoliata, enhances salt tolerance in Arabidopsis. Plant Mol. Biol. Rep. 2013, 31, 1548–1557. [Google Scholar] [CrossRef]
- Liu, W.; Feng, J.; Ma, W.; Zhou, Y.; Ma, Z. GhCLCg-1, a vacuolar chloride channel, contributes to salt tolerance by regulating ion accumulation in upland cotton. Front. Plant Sci. 2021, 12, 765173. [Google Scholar] [CrossRef] [PubMed]
- Alonso, J.M.; Stepanova, A.N.; Leisse, T.J.; Kim, C.J.; Chen, H.; Shinn, P.; Stevenson, D.K.; Zimmerman, J.; Barajas, P.; Cheuk, R.; et al. Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 2003, 301, 653–657. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, W.; Ma, Z.; Zhu, W.; Jia, L. Transcriptional characterization and response to defense elicitors of mevalonate pathway genes in cotton (Gossypium arboreum L.). PeerJ 2019, 7, e8123. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.J.; Sreedasyam, A.; Ando, A.; Song, Q.; De Santiago, L.M.; Hulse-Kemp, A.M.; Ding, M.; Ye, W.; Kirkbride, R.C.; Jenkins, J.; et al. Genomic diversifications of five Gossypium allopolyploid species and their impact on cotton improvement. Nat. Genet. 2020, 52, 525–533. [Google Scholar] [CrossRef] [PubMed]
- Rychlik, W. OLIGO 7 primer analysis software. Methods Mol. Biol. 2007, 402, 35–60. [Google Scholar] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Ma, W.; Ren, Z.; Zhou, Y.; Zhao, J.; Zhang, F.; Feng, J.; Liu, W.; Ma, X. Genome-wide identification of the Gossypium hirsutum NHX genes reveals that the endosomal-type GhNHX4A is critical for the salt tolerance of cotton. Int. J. Mol. Sci. 2020, 21, 7712. [Google Scholar] [CrossRef]
- Rotmistrovsky, K.; Jang, W.; Schuler, G.D. A web server for performing electronic PCR. Nucleic Acids Res. 2004, 32, W108–W112. [Google Scholar] [CrossRef]
- Sheen, J. Signal transduction in maize and Arabidopsis mesophyll protoplasts. Plant Physiol. 2001, 127, 1466–1475. [Google Scholar] [CrossRef]
- Zhang, X.; Henriques, R.; Lin, S.S.; Niu, Q.W.; Chua, N.H. Agrobacterium-mediated transformation of Arabidopsis thaliana using the floral dip method. Nat. Protoc. 2006, 1, 641–646. [Google Scholar] [CrossRef] [PubMed]
- An, C.; Wang, C.; Mou, Z. The Arabidopsis Elongator complex is required for nonhost resistance against the bacterial pathogens Xanthomonas citri subsp. citri and Pseudomonas syringae pv. phaseolicola NPS3121. New Phytol. 2017, 214, 1245–1259. [Google Scholar] [PubMed]
- Amo, J.; Lara, A.; Martínez-Martínez, A.; Martínez, V.; Rubio, F.; Nieves-Cordones, M. The protein kinase SlCIPK23 boosts K+ and Na+ uptake in tomato plants. Plant Cell Environ. 2021, 44, 3589–3605. [Google Scholar] [CrossRef] [PubMed]
- White, P.J.; Broadley, M.R. Chloride in soils and its uptake and movement within the plant: A review. Ann. Bot. 2001, 88, 967–988. [Google Scholar] [CrossRef]
- Jentsch, T.J.; Pusch, M.; Rehfeldt, A.; Steinmeyer, K. The ClC family of voltage-gated chloride channels: Structure and function. Ann. N. Y. Acad. Sci. 1993, 707, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Accardi, A. Structure and gating of CLC channels and exchangers. J. Physiol. 2015, 593, 4129–4138. [Google Scholar] [CrossRef]
- Zhang, X.K.; Zhou, Q.H.; Cao, J.H.; Yu, B.J. Differential Cl−/salt tolerance and NaCl-induced alternations of tissue and cellular ion fluxes in Glycine max, Glycine soja and their hybrid seedlings. J. Agron. Crop Sci. 2011, 197, 329–339. [Google Scholar] [CrossRef]
- Wissing, F.; Smith, J.A. Vacuolar chloride transport in Mesembryanthemum crystallinum L. measured using the fluorescent dye lucigenin. J. Membr. Biol. 2000, 177, 199–208. [Google Scholar] [CrossRef]
- Brini, F.; Masmoudi, K. Ion transporters and abiotic stress tolerance in plants. ISRN Mol. Biol. 2012, 2012, 927436. [Google Scholar] [CrossRef]
- Li, W.Y.; Wong, F.L.; Tsai, S.N.; Phang, T.H.; Shao, G.; Lam, H.M. Tonoplast-located GmCLC1 and GmNHX1 from soybean enhance NaCl tolerance in transgenic bright yellow (BY)-2 cells. Plant Cell Environ. 2006, 29, 1122–1137. [Google Scholar] [CrossRef]
- Zhou, G.A.; Qiu, L.J. Identification and functional analysis on abiotic stress response of soybean Cl− channel gene GmCLCnt. Agric. Sci. China 2010, 9, 199–206. [Google Scholar] [CrossRef]
- You, J.; Chan, Z. ROS regulation during abiotic stress responses in crop plants. Front. Plant Sci. 2015, 6, 1092. [Google Scholar] [CrossRef] [PubMed]
- Davey, M.W.; Stals, E.; Panis, B.; Keulemans, J.; Swennen, R.L. High-throughput determination of malondialdehyde in plant tissues. Anal. Biochem. 2005, 347, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotoxicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef] [PubMed]
- Stefanov, M.A.; Rashkov, G.D.; Apostolova, E.L. Assessment of the photosynthetic apparatus functions by chlorophyll fluorescence and P(700) absorbance in C3 and C4 plants under physiological conditions and under salt stress. Int. J. Mol. Sci. 2022, 23, 3768. [Google Scholar] [CrossRef] [PubMed]
- Nedelyaeva, O.I.; Popova, L.G.; Volkov, V.S.; Balnokin, Y.V. Molecular Cloning and characterization of SaCLCd, SaCLCf, and SaCLCg, novel proteins of the chloride channel family (CLC) from the halophyte Suaeda altissima (L.) Pall. Plants 2022, 11, 409. [Google Scholar] [CrossRef]
- Teakle, N.L.; Tyerman, S.D. Mechanisms of Cl− transport contributing to salt tolerance. Plant Cell Environ. 2010, 33, 566–589. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, W.; Gao, S.; Zhao, Y.; Wu, Y.; Li, X.; Li, J.; Zhu, W.; Ma, Z.; Liu, W. GhCLCc-1, a Chloride Channel Gene from Upland Cotton, Positively Regulates Salt Tolerance by Modulating the Accumulation of Chloride Ions. Genes 2024, 15, 555. https://doi.org/10.3390/genes15050555
Li W, Gao S, Zhao Y, Wu Y, Li X, Li J, Zhu W, Ma Z, Liu W. GhCLCc-1, a Chloride Channel Gene from Upland Cotton, Positively Regulates Salt Tolerance by Modulating the Accumulation of Chloride Ions. Genes. 2024; 15(5):555. https://doi.org/10.3390/genes15050555
Chicago/Turabian StyleLi, Wenhao, Siqi Gao, Yinghao Zhao, Yuchen Wu, Xiaona Li, Jianing Li, Wei Zhu, Zongbin Ma, and Wei Liu. 2024. "GhCLCc-1, a Chloride Channel Gene from Upland Cotton, Positively Regulates Salt Tolerance by Modulating the Accumulation of Chloride Ions" Genes 15, no. 5: 555. https://doi.org/10.3390/genes15050555
APA StyleLi, W., Gao, S., Zhao, Y., Wu, Y., Li, X., Li, J., Zhu, W., Ma, Z., & Liu, W. (2024). GhCLCc-1, a Chloride Channel Gene from Upland Cotton, Positively Regulates Salt Tolerance by Modulating the Accumulation of Chloride Ions. Genes, 15(5), 555. https://doi.org/10.3390/genes15050555