
mRNA Expression and DNA Methylation Analysis of Serotonin Receptor 2A (HTR2A) in the Human Schizophrenic Brain

Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples
2.2. mRNA Data Generation
2.3. mRNA Data Transformation
2.4. Genotyping
2.5. CpG Site Methylation Data Generation and Transformation
2.6. Data Analysis
3. Results
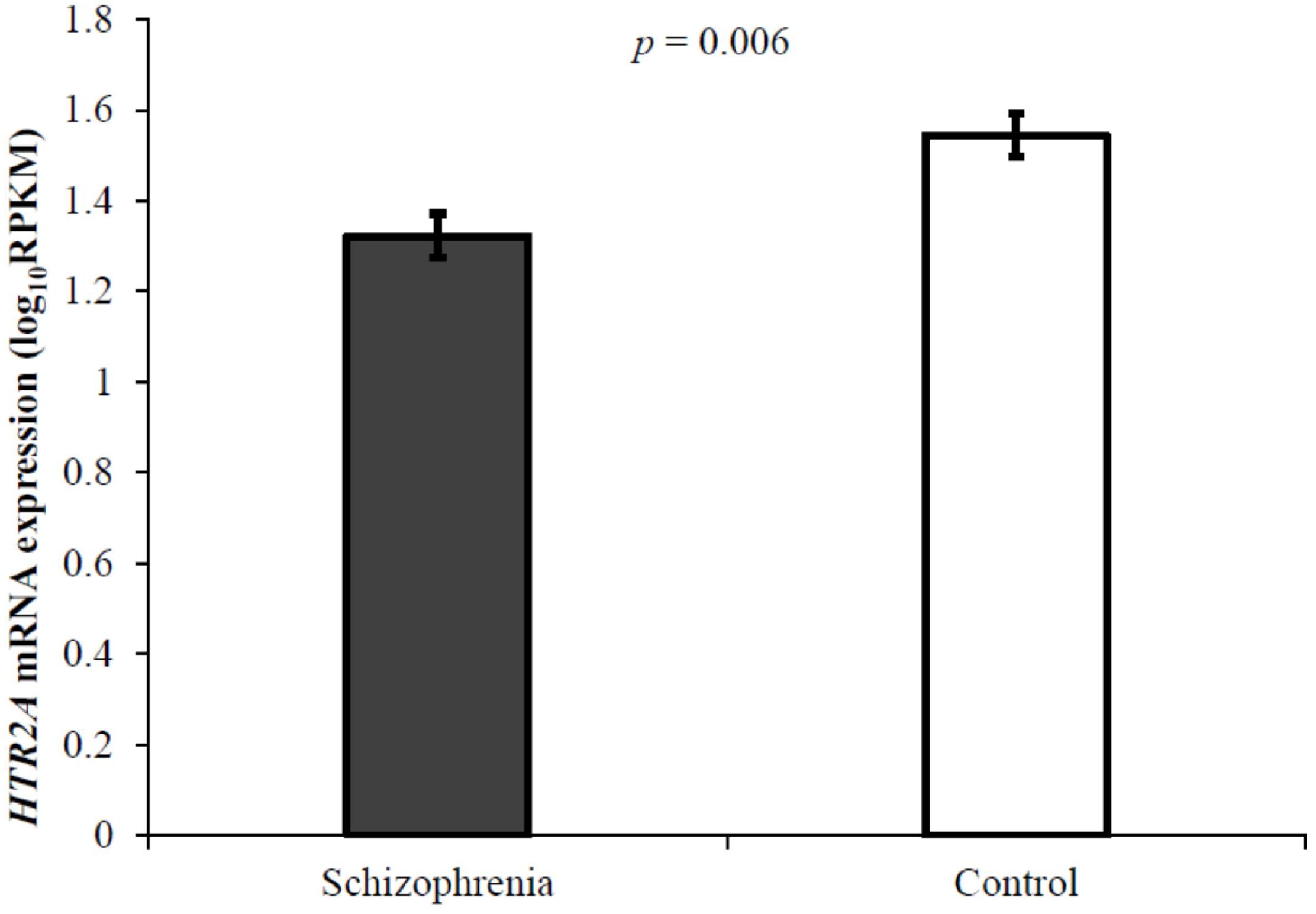
3.1. Schizophrenia and HTR2A mRNA Expression
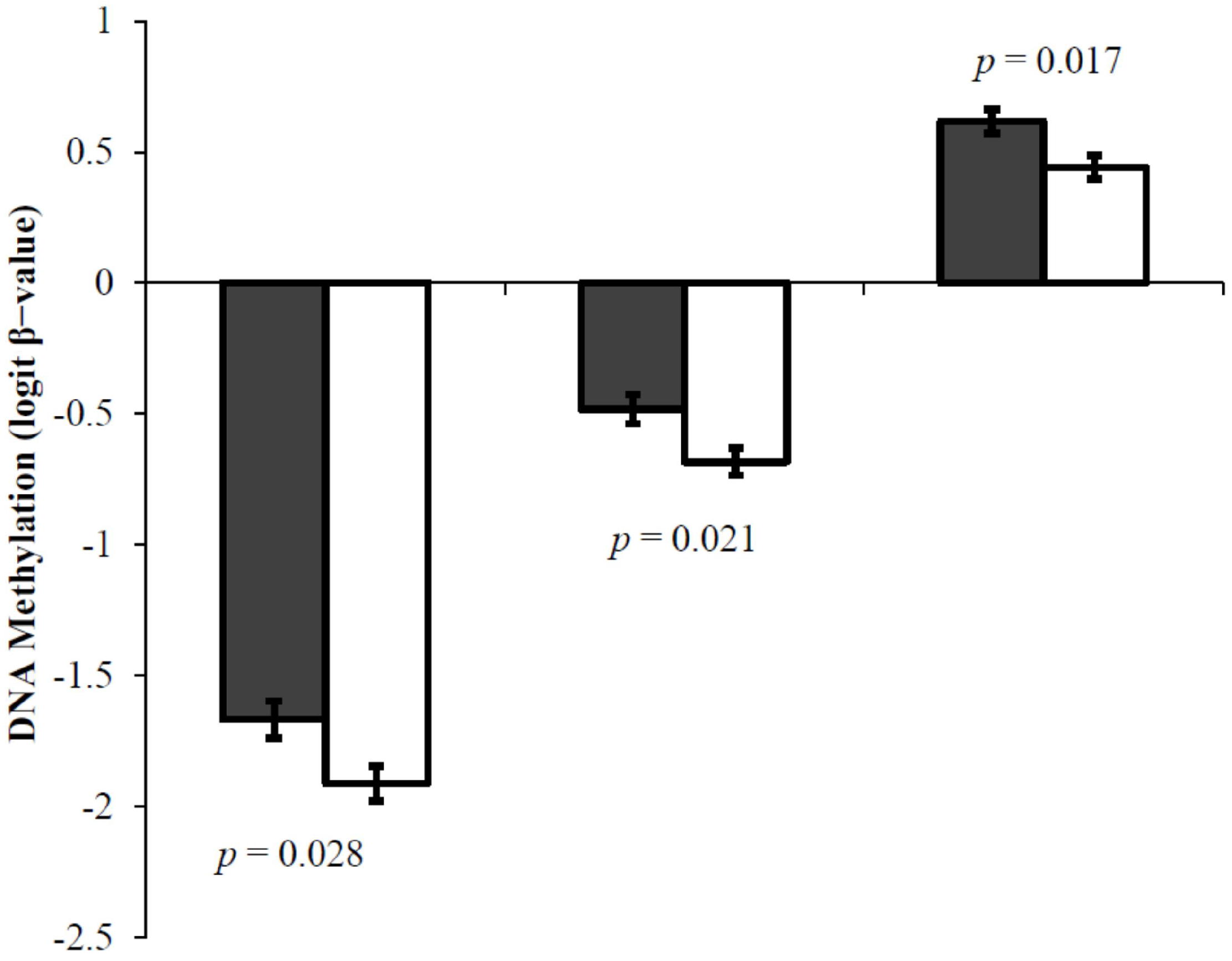
3.2. Schizophrenia and DNA Methylation
3.3. rs6314 and rs6313 Genotypes
3.4. HTR2A mRNA Expression and Genotypes
3.5. DNA Methylation and Genotypes
3.6. Correlation between mRNA Expression and DNA Methylation
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Buhot, M.C. Serotonin receptors in cognitive behaviors. Curr. Opin. Neurobiol. 1997, 7, 243–254. [Google Scholar] [CrossRef]
- Fink, K.B.; Gothert, M. 5-HT receptor regulation of neurotransmitter release. Pharmacol. Rev. 2007, 59, 360–417. [Google Scholar] [CrossRef] [PubMed]
- Jakab, R.L.; Goldman-Rakic, P.S. Segregation of serotonin 5-HT2A and 5-HT3 receptors in inhibitory circuits of the primate cerebral cortex. J. Comp. Neurol. 2000, 417, 337–348. [Google Scholar] [CrossRef]
- Willins, D.L.; Deutch, A.Y.; Roth, B.L. Serotonin 5-HT2A receptors are expressed on pyramidal cells and interneurons in the rat cortex. Synapse 1997, 27, 79–82. [Google Scholar] [CrossRef]
- Kim, Y.K.; Yoon, H.K. Effect of serotonin-related gene polymorphisms on pathogenesis and treatment response in korean schizophrenic patients. Behav. Genet. 2011, 41, 709–715. [Google Scholar] [CrossRef] [PubMed]
- Lang, U.E.; Puls, I.; Muller, D.J.; Strutz-Seebohm, N.; Gallinat, J. Molecular mechanisms of schizophrenia. Cell. Physiol. Biochem. 2007, 20, 687–702. [Google Scholar] [CrossRef] [PubMed]
- Harvey, J.A. Role of the serotonin 5-HT(2A) receptor in learning. Learn. Mem. 2003, 10, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Weickert, T.W.; Goldberg, T.E.; Gold, J.M.; Bigelow, L.B.; Egan, M.F.; Weinberger, D.R. Cognitive impairments in patients with schizophrenia displaying preserved and compromised intellect. Arch. Gen. Psychiatry 2000, 57, 907–913. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, S.; Duncan, G.E.; Marx, C.E.; Lieberman, J.A. Treatments for schizophrenia: A critical review of pharmacology and mechanisms of action of antipsychotic drugs. Mol. Psychiatry 2005, 10, 79–104. [Google Scholar] [CrossRef] [PubMed]
- Abdolmaleky, H.M.; Yaqubi, S.; Papageorgis, P.; Lambert, A.W.; Ozturk, S.; Sivaraman, V.; Thiagalingam, S. Epigenetic dysregulation of HTR2A in the brain of patients with schizophrenia and bipolar disorder. Schizophr. Res. 2011, 129, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Abi-Dargham, A. Alterations of serotonin transmission in schizophrenia. Int. Rev. Neurobiol. 2007, 78, 133–164. [Google Scholar] [PubMed]
- Polesskaya, O.O.; Sokolov, B.P. Differential expression of the “C” and “T” alleles of the 5-HT2A receptor gene in the temporal cortex of normal individuals and schizophrenics. J. Neurosci. Res. 2002, 67, 812–822. [Google Scholar] [CrossRef] [PubMed]
- Selvaraj, S.; Arnone, D.; Cappai, A.; Howes, O. Alterations in the serotonin system in schizophrenia: A systematic review and meta-analysis of postmortem and molecular imaging studies. Neurosci. Biobehav. Rev. 2014, 45, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Maeso, J.; Ang, R.L.; Yuen, T.; Chan, P.; Weisstaub, N.V.; Lopez-Gimenez, J.F.; Zhou, M.; Okawa, Y.; Callado, L.F.; Milligan, G.; et al. Identification of a serotonin/glutamate receptor complex implicated in psychosis. Nature 2008, 452, 93–97. [Google Scholar] [CrossRef] [PubMed]
- Muguruza, C.; Moreno, J.L.; Umali, A.; Callado, L.F.; Meana, J.J.; Gonzalez-Maeso, J. Dysregulated 5-HT(2A) receptor binding in postmortem frontal cortex of schizophrenic subjects. Eur. Neuropsychopharmacol. 2013, 23, 852–864. [Google Scholar] [CrossRef] [PubMed]
- Maple, A.M.; Zhao, X.; Elizalde, D.I.; McBride, A.K.; Gallitano, A.L. Htr2a expression responds rapidly to environmental stimuli in an Egr3-dependent manner. ACS Chem. Neurosci. 2015, 6, 1137–1142. [Google Scholar] [CrossRef] [PubMed]
- Schmid, C.L.; Streicher, J.M.; Meltzer, H.Y.; Bohn, L.M. Clozapine acts as an agonist at serotonin 2A receptors to counter MK-801-induced behaviors through a betaarrestin2-independent activation of AKT. Neuropsychopharmacology 2014, 39, 1902–1913. [Google Scholar] [CrossRef] [PubMed]
- Hazelwood, L.A.; Sanders-Bush, E. His452Tyr polymorphism in the human 5-HT2A receptor destabilizes the signaling conformation. Mol. Pharmacol. 2004, 66, 1293–1300. [Google Scholar] [PubMed]
- Ozaki, N.; Manji, H.; Lubierman, V.; Lu, S.J.; Lappalainen, J.; Rosenthal, N.E.; Goldman, D. A naturally occurring amino acid substitution of the human serotonin 5-HT2A receptor influences amplitude and timing of intracellular calcium mobilization. J. Neurochem. 1997, 68, 2186–2193. [Google Scholar] [CrossRef] [PubMed]
- Serretti, A.; Drago, A.; De Ronchi, D. HTR2A gene variants and psychiatric disorders: A review of current literature and selection of snps for future studies. Curr. Med. Chem. 2007, 14, 2053–2069. [Google Scholar] [CrossRef] [PubMed]
- Blasi, G.; De Virgilio, C.; Papazacharias, A.; Taurisano, P.; Gelao, B.; Fazio, L.; Ursini, G.; Sinibaldi, L.; Andriola, I.; Masellis, R.; et al. Converging evidence for the association of functional genetic variation in the serotonin receptor 2A gene with prefrontal function and olanzapine treatment. JAMA Psychiatry 2013, 70, 921–930. [Google Scholar] [CrossRef] [PubMed]
- Filippini, N.; Scassellati, C.; Boccardi, M.; Pievani, M.; Testa, C.; Bocchio-Chiavetto, L.; Frisoni, G.B.; Gennarelli, M. Influence of serotonin receptor 2A His452Tyr polymorphism on brain temporal structures: A volumetric MR study. Eur. J. Hum. Genet. 2006, 14, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Schott, B.H.; Seidenbecher, C.I.; Richter, S.; Wustenberg, T.; Debska-Vielhaber, G.; Schubert, H.; Heinze, H.J.; Richardson-Klavehn, A.; Duzel, E. Genetic variation of the serotonin 2A receptor affects hippocampal novelty processing in humans. PLoS ONE 2011, 6, e15984. [Google Scholar] [CrossRef] [PubMed]
- De Quervain, D.J.; Henke, K.; Aerni, A.; Coluccia, D.; Wollmer, M.A.; Hock, C.; Nitsch, R.M.; Papassotiropoulos, A. A functional genetic variation of the 5-HT2A receptor affects human memory. Nat. Neurosci. 2003, 6, 1141–1142. [Google Scholar] [CrossRef] [PubMed]
- Arranz, M.J.; Collier, D.A.; Munro, J.; Sham, P.; Kirov, G.; Sodhi, M.; Roberts, G.; Price, J.; Kerwin, R.W. Analysis of a structural polymorphism in the 5-HT2A receptor and clinical response to clozapine. Neurosci. Lett. 1996, 217, 177–178. [Google Scholar] [CrossRef]
- Arranz, M.J.; Munro, J.; Owen, M.J.; Spurlock, G.; Sham, P.C.; Zhao, J.; Kirov, G.; Collier, D.A.; Kerwin, R.W. Evidence for association between polymorphisms in the promoter and coding regions of the 5-HT2A receptor gene and response to clozapine. Mol. Psychiatry 1998, 3, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Abdolmaleky, H.M.; Faraone, S.V.; Glatt, S.J.; Tsuang, M.T. Meta-analysis of association between the T102C polymorphism of the 5ht2a receptor gene and schizophrenia. Schizophr. Res. 2004, 67, 53–62. [Google Scholar] [CrossRef]
- Khait, V.D.; Huang, Y.Y.; Zalsman, G.; Oquendo, M.A.; Brent, D.A.; Harkavy-Friedman, J.M.; Mann, J.J. Association of serotonin 5-HT2A receptor binding and the T102C polymorphism in depressed and healthy caucasian subjects. Neuropsychopharmacology 2005, 30, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Yildiz, S.H.; Akilli, A.; Bagcioglu, E.; Ozdemir Erdogan, M.; Coskun, K.S.; Alpaslan, A.H.; Subasi, B.; Arikan Terzi, E.S. Association of schizophrenia with T102C (rs6313) and 1438 a/g (rs6311) polymorphisms of HTR2A gene. Acta Neuropsychiatr. 2013, 25, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Ucok, A.; Alpsan, H.; Cakir, S.; Saruhan-Direskeneli, G. Association of a serotonin receptor 2a gene polymorphism with cognitive functions in patients with schizophrenia. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2007, 144B, 704–707. [Google Scholar] [CrossRef] [PubMed]
- Vyas, N.S.; Lee, Y.; Ahn, K.; Ternouth, A.; Stahl, D.R.; Al-Chalabi, A.; Powell, J.F.; Puri, B.K. Association of a serotonin receptor 2A gene polymorphism with visual sustained attention in early-onset schizophrenia patients and their non-psychotic siblings. Aging Dis. 2012, 3, 291–300. [Google Scholar] [PubMed]
- Turecki, G.; Sequeira, A.; Gingras, Y.; Seguin, M.; Lesage, A.; Tousignant, M.; Chawky, N.; Vanier, C.; Lipp, O.; Benkelfat, C.; et al. Suicide and serotonin: Study of variation at seven serotonin receptor genes in suicide completers. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2003, 118B, 36–40. [Google Scholar] [CrossRef] [PubMed]
- Parsons, M.J.; D’Souza, U.M.; Arranz, M.J.; Kerwin, R.W.; Makoff, A.J. The -1438a/g polymorphism in the 5-hydroxytryptamine type 2A receptor gene affects promoter activity. Biol. Psychiatry 2004, 56, 406–410. [Google Scholar] [CrossRef] [PubMed]
- Wockner, L.F.; Noble, E.P.; Lawford, B.R.; Young, R.M.; Morris, C.P.; Whitehall, V.L.; Voisey, J. Genome-wide DNA methylation analysis of human brain tissue from schizophrenia patients. Transl. Psychiatry 2014, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paquette, A.G.; Lesseur, C.; Armstrong, D.A.; Koestler, D.C.; Appleton, A.A.; Lester, B.M.; Marsit, C.J. Placental HTR2A methylation is associated with infant neurobehavioral outcomes. Epigenetics 2013, 8, 796–801. [Google Scholar] [CrossRef] [PubMed]
- Ghadirivasfi, M.; Nohesara, S.; Ahmadkhaniha, H.R.; Eskandari, M.R.; Mostafavi, S.; Thiagalingam, S.; Abdolmaleky, H.M. Hypomethylation of the serotonin receptor type-2A gene (HTR2A) at T102C polymorphic site in DNA derived from the saliva of patients with schizophrenia and bipolar disorder. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2011, 156B, 536–545. [Google Scholar] [CrossRef] [PubMed]
- De Luca, V.; Viggiano, E.; Dhoot, R.; Kennedy, J.L.; Wong, A.H. Methylation and qtdt analysis of the 5-HT2A receptor 102C allele: Analysis of suicidality in major psychosis. J. Psychiatr. Res. 2009, 43, 532–537. [Google Scholar] [CrossRef] [PubMed]
- Polesskaya, O.O.; Aston, C.; Sokolov, B.P. Allele c-specific methylation of the 5-HT2A receptor gene: Evidence for correlation with its expression and expression of DNA methylase DNMT1. J. Neurosci. Res. 2006, 83, 362–373. [Google Scholar] [CrossRef] [PubMed]
- Falkenberg, V.R.; Gurbaxani, B.M.; Unger, E.R.; Rajeevan, M.S. Functional genomics of serotonin receptor 2a (HTR2A): Interaction of polymorphism, methylation, expression and disease association. Neuromol. Med. 2011, 13, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Paquette, A.G.; Marsit, C.J. The developmental basis of epigenetic regulation of HTR2A and psychiatric outcomes. J. Cell. Biochem. 2014, 115, 2065–2072. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-Seq experiments with tophat and cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Smyth, G.K.; Shi, W. The subread aligner: Fast, accurate and scalable read mapping by seed-and-vote. Nucleic Acids Res. 2013, 41. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, D.J.; Chen, Y.; Smyth, G.K. Differential expression analysis of multifactor RNA-Seq experiments with respect to biological variation. Nucleic Acids Res. 2012, 40, 4288–4297. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; Oshlack, A. A scaling normalization method for differential expression analysis of RNA-Seq data. Genome Biol. 2010, 11. [Google Scholar] [CrossRef] [PubMed]
- Schirmer, M.; Ijaz, U.Z.; D'Amore, R.; Hall, N.; Sloan, W.T.; Quince, C. Insight into biases and sequencing errors for amplicon sequencing with the illumina miseq platform. Nucleic Acids Res. 2015, 43. [Google Scholar] [CrossRef] [PubMed]
- Wockner, L.F.; Morris, C.P.; Noble, E.P.; Lawford, B.R.; Whitehall, V.L.; Young, R.M.; Voisey, J. Brain-specific epigenetic markers of schizophrenia. Transl. Psychiatry 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Guhathakurta, S.; Singh, A.S.; Sinha, S.; Chatterjee, A.; Ahmed, S.; Ghosh, S.; Usha, R. Analysis of serotonin receptor 2A gene (HTR2A): Association study with autism spectrum disorder in the Indian population and investigation of the gene expression in peripheral blood leukocytes. Neurochem. Int. 2009, 55, 754–759. [Google Scholar] [CrossRef] [PubMed]
- Dammann, G.; Teschler, S.; Haag, T.; Altmuller, F.; Tuczek, F.; Dammann, R.H. Increased DNA methylation of neuropsychiatric genes occurs in borderline personality disorder. Epigenetics 2011, 6, 1454–1462. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Control (n = 23) | Schizophrenia (n = 22) | t-Test/Chi-Square p-Value | |
|---|---|---|---|
| Age, mean (s.d.) | 70.2 (9.2) | 52.5 (22.7) | 0.0022 |
| PMI, mean (s.d.) | 14.1 (3.2) | 24 (10.6) | 0.0003 |
| Sex, male (%) | 18 (78) | 14 (64) | 0.2778 |
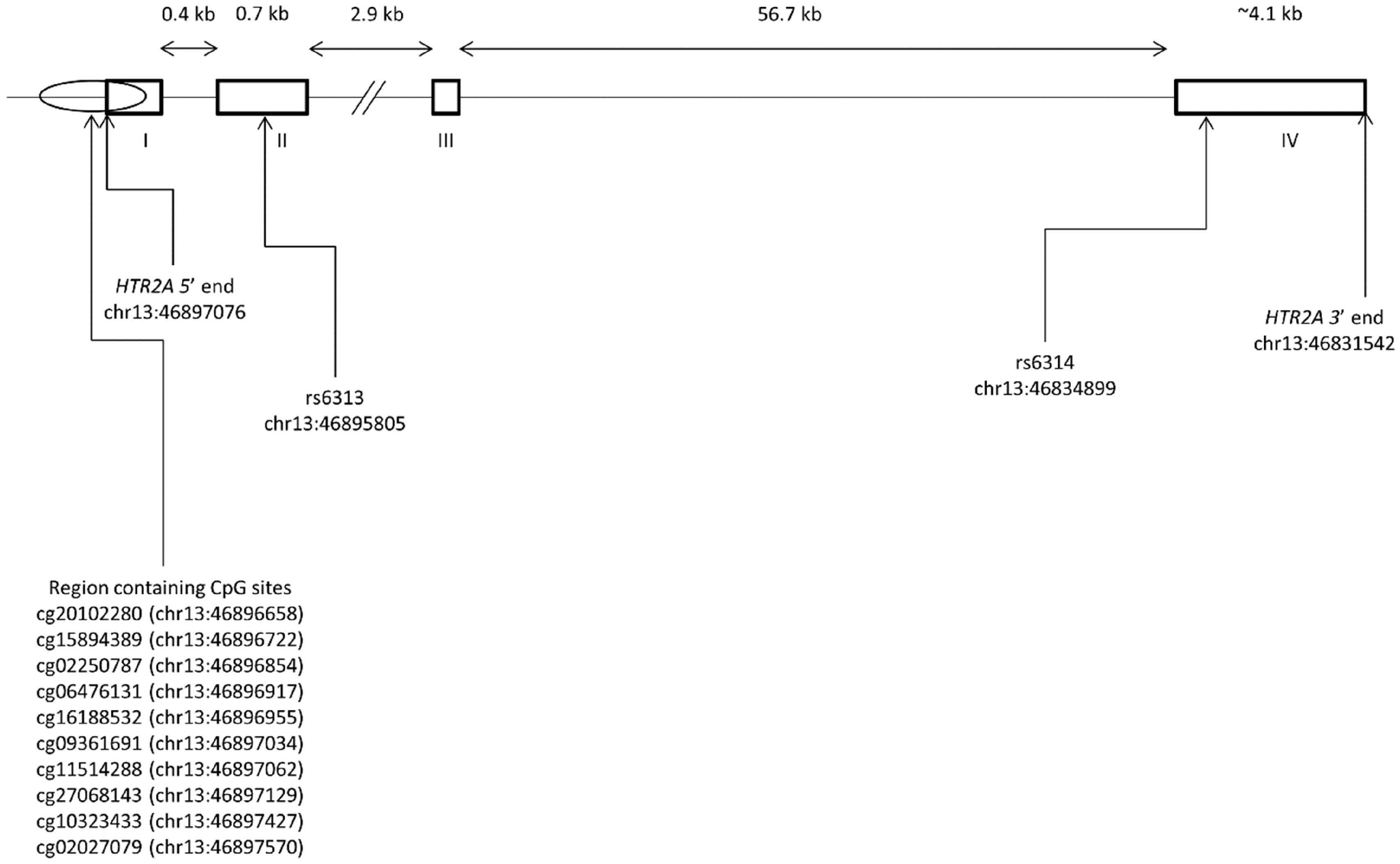
| CpG Site | Illumina CpG Site Name | Feature | Locus |
|---|---|---|---|
| cg1 | cg20102280 | Exon I | chr13:46896658 |
| cg2 | cg15894389 | Exon I | chr13:46896722 |
| cg3 | cg02250787 | Exon I | chr13:46896854 |
| cg4 | cg06476131 | Exon I | chr13:46896917 |
| cg5 | cg16188532 | Exon I | chr13:46896955 |
| cg6 | cg09361691 | Exon I | chr13:46897034 |
| cg7 | cg11514288 | Exon I | chr13:46897062 |
| cg8 | cg27068143 | Promoter | chr13:46897129 |
| cg9 | cg10323433 | Promoter | chr13:46897427 |
| cg10 | cg02027079 | Promoter | chr13:46897570 |
| Genotype/Allele | Schizophrenia | Control | Chi-Square p-Value | Odds Ratio | |
|---|---|---|---|---|---|
| rs6314 Genotype | C/C | 10 | 19 | 0.008 | 5.7 |
| C/T | 12 | 4 | |||
| T/T | 0 | 0 | |||
| Total | 22 | 23 | |||
| rs6314 Allele | C | 32 | 42 | 0.019 | 3.9 |
| T | 12 | 4 | |||
| Total | 44 | 46 | |||
| rs6313 Genotype | C/C | 6 | 7 | 0.078 | -- |
| C/T | 9 | 14 | |||
| T/T | 6 | 1 | |||
| Total | 21 | 22 | |||
| rs6313 Allele | C | 21 | 28 | 0.201 | -- |
| T | 21 | 16 | |||
| Total | 42 | 44 |
© 2017 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheah, S.-Y.; Lawford, B.R.; Young, R.M.; Morris, C.P.; Voisey, J. mRNA Expression and DNA Methylation Analysis of Serotonin Receptor 2A (HTR2A) in the Human Schizophrenic Brain. Genes 2017, 8, 14. https://doi.org/10.3390/genes8010014
Cheah S-Y, Lawford BR, Young RM, Morris CP, Voisey J. mRNA Expression and DNA Methylation Analysis of Serotonin Receptor 2A (HTR2A) in the Human Schizophrenic Brain. Genes. 2017; 8(1):14. https://doi.org/10.3390/genes8010014
Chicago/Turabian StyleCheah, Sern-Yih, Bruce R. Lawford, Ross McD. Young, Charles P. Morris, and Joanne Voisey. 2017. "mRNA Expression and DNA Methylation Analysis of Serotonin Receptor 2A (HTR2A) in the Human Schizophrenic Brain" Genes 8, no. 1: 14. https://doi.org/10.3390/genes8010014
APA StyleCheah, S. -Y., Lawford, B. R., Young, R. M., Morris, C. P., & Voisey, J. (2017). mRNA Expression and DNA Methylation Analysis of Serotonin Receptor 2A (HTR2A) in the Human Schizophrenic Brain. Genes, 8(1), 14. https://doi.org/10.3390/genes8010014