
Burkholderia cepacia Complex Regulation of Virulence Gene Expression: A Review




Abstract
:1. Introduction
2. Genomics and Transcriptomics Analysis in the Study of Bcc Pathogenesis
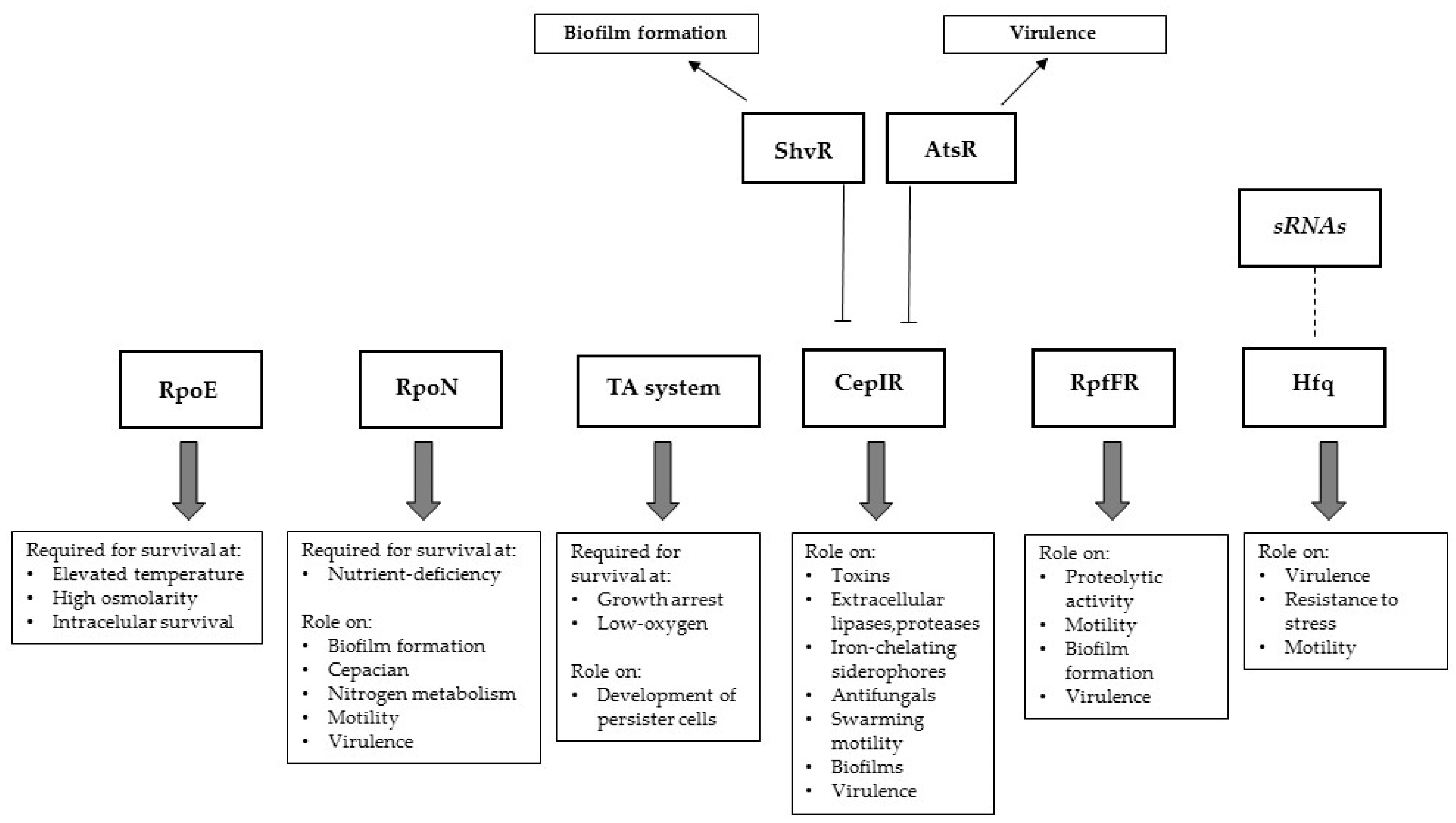
3. Known Regulators of Bcc Virulence Factors
3.1. Sigma Factors and Related Proteins
3.2. Quorum Sensing
3.3. Iron Acquisition and Bcc Virulence
3.4. Activators and Repressors
3.5. Two-Component Regulatory Systems
3.6. Toxin-Antitoxin
3.7. sRNA and Hfq
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Holden, M.T.; Seth-Smith, H.M.; Crossman, L.C.; Sebaihia, M.; Bentley, S.D.; Cerdeno-Tarraga, A.M.; Thomson, N.R.; Bason, N.; Quail, M.A.; Sharp, S.; et al. The Genome of Burkholderia cenocepacia J2315, an Epidemic Pathogen of Cystic Fibrosis Patients. J. Bacteriol. 2009, 191, 261–277. [Google Scholar] [CrossRef] [PubMed]
- Ussery, D.W.; Kiil, K.; Lagesen, K.; Sicheritz-Pont-eacuten, T.; Bohlin, J.; Wassenaar, T.M. The Genus Burkholderia: Analysis of 56 Genomic Sequences. In Microbial Pathogenomics; KARGER: Basel, Switzerland, 2009; Volume 6, pp. 140–157. [Google Scholar]
- Depoorter, E.; Bull, M.J.; Peeters, C.; Coenye, T.; Vandamme, P.; Mahenthiralingam, E. Burkholderia: An update on taxonomy and biotechnological potential as antibiotic producers. Appl. Microbiol. Biotechnol. 2016, 100, 5215–5229. [Google Scholar] [CrossRef] [PubMed]
- Mahenthiralingam, E.; Urban, T.A.; Goldberg, J.B. The multifarious, multireplicon Burkholderia cepacia complex. Nat. Rev. Microbiol. 2005, 3, 144–156. [Google Scholar] [CrossRef] [PubMed]
- Leitão, J.H.; Sousa, S.A.; Ferreira, A.S.; Ramos, C.G.; Silva, I.N.; Moreira, L.M. Pathogenicity, virulence factors, and strategies to fight against Burkholderia cepacia complex pathogens and related species. Appl. Microbiol. Biotechnol. 2010, 87, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Lipuma, J.J. The changing microbial epidemiology in cystic fibrosis. Clin. Microbiol. Rev. 2010, 23, 299–323. [Google Scholar] [CrossRef] [PubMed]
- Leitão, J.H.; Sousa, S.A.; Cunha, M.V.; Salgado, M.J.; Melo-Cristino, J.; Barreto, M.C.; Sá-Correia, I. Variation of the antimicrobial susceptibility profiles of Burkholderia cepacia complex clonal isolates obtained from chronically infected cystic fibrosis patients: A five-year survey in the major Portuguese treatment center. Eur. J. Clin. Microbiol. Infect. Dis. 2008, 27, 1101–1111. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.M.; Dodd, M.E.; Webb, A.K. Burkholderia cepacia: Current clinical issues, environmental controversies and ethical dilemmas. Eur. Respir. J. 2001, 17, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Loutet, S.A.; Valvano, M.A. A decade of Burkholderia cenocepacia virulence determinant research. Infect. Immun. 2010, 78, 4088–4100. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Jelsbak, L.; Molin, S. Microbial ecology and adaptation in cystic fibrosis airways. Environ. Microbiol. 2011, 13, 1682–1689. [Google Scholar] [CrossRef] [PubMed]
- Sass, A.M.; Schmerk, C.; Agnoli, K.; Norville, P.J.; Eberl, L.; Valvano, M.A.; Mahenthiralingam, E. The unexpected discovery of a novel low-oxygen-activated locus for the anoxic persistence of Burkholderia cenocepacia. ISME J. 2013, 14, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saldías, M.S.; Valvano, M.A. Interactions of Burkholderia cenocepacia and other Burkholderia cepacia complex bacteria with epithelial and phagocytic cells. Microbiology 2009, 155, 2809–2817. [Google Scholar] [CrossRef] [PubMed]
- Lieberman, T.D.; Michel, J.-B.; Aingaran, M.; Potter-Bynoe, G.; Roux, D.; Davis, M.R.; Skurnik, D.; Leiby, N.; LiPuma, J.J.; Goldberg, J.B.; et al. Parallel bacterial evolution within multiple patients identifies candidate pathogenicity genes. Nat. Genet. 2011, 43, 1275–1280. [Google Scholar] [CrossRef] [PubMed]
- Lieberman, T.D.; Flett, K.B.; Yelin, I.; Martin, T.R.; McAdam, A.J.; Priebe, G.P.; Kishony, R. Genetic variation of a bacterial pathogen within individuals with cystic fibrosis provides a record of selective pressures. Nat. Genet. 2014, 46, 82–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, I.N.; Santos, P.M.; Santos, M.R.; Zlosnik, J.E.A.; Speert, D.P.; Buskirk, S.W.; Bruger, E.L.; Waters, C.M.; Cooper, V.S.; Moreira, L.M. Long-Term Evolution of Burkholderia multivorans during a Chronic Cystic Fibrosis Infection Reveals Shifting Forces of Selection. mSystems 2016, 1. [Google Scholar] [CrossRef] [PubMed]
- Nunvar, J.; Kalferstova, L.; Bloodworth, R.A.M.; Kolar, M.; Degrossi, J.; Lubovich, S.; Cardona, S.T.; Drevinek, P.; Bevivino, A. Understanding the Pathogenicity of Burkholderia contaminans, an Emerging Pathogen in Cystic Fibrosis. PLoS ONE 2016, 11, e0160975. [Google Scholar] [CrossRef] [PubMed]
- Mira, N.P.; Madeira, A.; Moreira, A.S.; Coutinho, C.P.; Sá-Correia, I. Genomic Expression Analysis Reveals Strategies of Burkholderia cenocepacia to Adapt to Cystic Fibrosis Patients’ Airways and Antimicrobial Therapy. PLoS ONE 2011, 6, e28831. [Google Scholar] [CrossRef] [PubMed]
- Sass, A.; Marchbank, A.; Tullis, E.; Lipuma, J.J.; Mahenthiralingam, E. Spontaneous and evolutionary changes in the antibiotic resistance of Burkholderia cenocepacia observed by global gene expression analysis. BMC Genom. 2011, 12, 373. [Google Scholar] [CrossRef] [PubMed]
- Kalferstova, L.; Kolar, M.; Fila, L.; Vavrova, J.; Drevinek, P. Gene expression profiling of Burkholderia cenocepacia at the time of cepacia syndrome: Loss of motility as a marker of poor prognosis? J. Clin. Microbiol. 2015, 53, 1515–1522. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.E.; Buckley, D.G.; Wu, Z.; Saenphimmachak, C.; Hoffman, L.R.; D’Argenio, D.A.; Miller, S.I.; Ramsey, B.W.; Speert, D.P.; Moskowitz, S.M.; et al. Genetic adaptation by Pseudomonas aeruginosa to the airways of cystic fibrosis patients. Proc. Natl. Acad. Sci. USA 2006, 103, 8487–8492. [Google Scholar] [CrossRef] [PubMed]
- Deitsch, K.W.; Lukehart, S.A.; Stringer, J.R. Common strategies for antigenic variation by bacterial, fungal and protozoan pathogens. Nat. Rev. Microbiol. 2009, 7, 493–503. [Google Scholar] [CrossRef] [PubMed]
- Rau, M.H.; Hansen, S.K.; Johansen, H.K.; Thomsen, L.E.; Workman, C.T.; Nielsen, K.F.; Jelsbak, L.; Høiby, N.; Yang, L.; Molin, S. Early adaptive developments of Pseudomonas aeruginosa after the transition from life in the environment to persistent colonization in the airways of human cystic fibrosis hosts. Environ. Microbiol. 2010, 12, 1643–1658. [Google Scholar] [PubMed]
- Crosson, S.; McGrath, P.T.; Stephens, C.; McAdams, H.H.; Shapiro, L. Conserved modular design of an oxygen sensory/signaling network with species-specific output. Proc. Natl. Acad. Sci. USA 2005, 102, 8018–8023. [Google Scholar] [CrossRef] [PubMed]
- Worlitzsch, D.; Tarran, R.; Ulrich, M.; Schwab, U.; Cekici, A.; Meyer, K.C.; Birrer, P.; Bellon, G.; Berger, J.; Weiss, T.; et al. Effects of reduced mucus oxygen concentration in airway Pseudomonas infections of cystic fibrosis patients. J. Clin. Investig. 2002, 109, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Govan, J.R.; Brown, A.R.; Jones, A.M. Evolving epidemiology of Pseudomonas aeruginosa and the Burkholderia cepacia complex in cystic fibrosis lung infection. Future Microbiol. 2007, 2, 153–164. [Google Scholar] [CrossRef] [PubMed]
- Martina, P.; Feliziani, S.; Juan, C.; Bettiol, M.; Gatti, B.; Yantorno, O.; Smania, A.M.; Oliver, A.; Bosch, A. Hypermutation in Burkholderia cepacia complex is mediated by DNA mismatch repair inactivation and is highly prevalent in cystic fibrosis chronic respiratory infection. Int. J. Med. Microbiol. 2014, 304, 1182–1191. [Google Scholar] [CrossRef] [PubMed]
- Oliver, A.; Mena, A. Bacterial hypermutation in cystic fibrosis, not only for antibiotic resistance. Clin. Microbiol. Infect. 2010, 16, 798–808. [Google Scholar] [CrossRef] [PubMed]
- Coenye, T.; Drevinek, P.; Mahenthiralingam, E.; Shah, S.A.; Gill, R.T.; Vandamme, P.; Ussery, D.W. Identification of putative noncoding RNA genes in the Burkholderia cenocepacia J2315 genome. FEMS Microbiol. Lett. 2007, 276, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Drevinek, P.; Holden, M.T.G.; Ge, Z.; Jones, A.M.; Ketchell, I.; Gill, R.T.; Mahenthiralingam, E. Gene expression changes linked to antimicrobial resistance, oxidative stress, iron depletion and retained motility are observed when Burkholderia cenocepacia grows in cystic fibrosis sputum. BMC Infect. Dis. 2008, 8, 121. [Google Scholar] [CrossRef] [PubMed]
- Yoder-Himes, D.R.; Chain, P.S.G.; Zhu, Y.; Wurtzel, O.; Rubin, E.M.; Tiedje, J.M.; Sorek, R. Mapping the Burkholderia cenocepacia niche response via high-throughput sequencing. Proc. Natl. Acad. Sci. USA 2009, 106, 3976–3981. [Google Scholar] [CrossRef] [PubMed]
- Yoder-Himes, D.R.; Konstantinidis, K.T.; Tiedje, J.M. Identification of potential therapeutic targets for Burkholderia cenocepacia by comparative transcriptomics. PLoS ONE 2010, 5, e8724. [Google Scholar] [CrossRef] [PubMed]
- O’Grady, E.P.; Viteri, D.F.; Malott, R.J.; Sokol, P.A. Reciprocal regulation by the CepIR and CciIR quorum sensing systems in Burkholderia cenocepacia. BMC Genom. 2009, 10, 441. [Google Scholar] [CrossRef] [PubMed]
- Inhülsen, S.; Aguilar, C.; Schmid, N.; Suppiger, A.; Riedel, K.; Eberl, L. Identification of functions linking quorum sensing with biofilm formation in Burkholderia cenocepacia H111. Microbiologyopen 2012, 1, 225–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmid, N.; Pessi, G.; Deng, Y.; Aguilar, C.; Carlier, A.L.; Grunau, A.; Omasits, U.; Zhang, L.H.; Ahrens, C.H.; Eberl, L. The AHL- and BDSF-Dependent Quorum Sensing Systems Control Specific and Overlapping Sets of Genes in Burkholderia cenocepacia H111. PLoS ONE 2012, 7, e49966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bazzini, S.; Udine, C.; Sass, A.; Pasca, M.R.; Longo, F.; Emiliani, G.; Fondi, M.; Perrin, E.; Decorosi, F.; Viti, C.; et al. Deciphering the role of rnd efflux transporters in Burkholderia cenocepacia. PLoS ONE 2011, 6, e18902. [Google Scholar] [CrossRef] [PubMed]
- Peeters, E.; Sass, A.; Mahenthiralingam, E.; Nelis, H.; Coenye, T. Transcriptional response of Burkholderia cenocepacia J2315 sessile cells to treatments with high doses of hydrogen peroxide and sodium hypochlorite. BMC Genom. 2010, 11, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Acker, H.; Sass, A.; Dhondt, I.; Nelis, H.J.; Coenye, T. Involvement of toxin-antitoxin modules in Burkholderia cenocepacia biofilm persistence. Pathog. Dis. 2014, 71, 326–335. [Google Scholar] [CrossRef] [PubMed]
- Sass, A.M.; van Acker, H.; Förstner, K.U.; van Nieuwerburgh, F.; Deforce, D.; Vogel, J.; Coenye, T. Genome-wide transcription start site profiling in biofilm-grown Burkholderia cenocepacia J2315. BMC Genom. 2015, 16, 775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coenye, T.; van Acker, H.; Peeters, E.; Sass, A.; Buroni, S.; Riccardi, G.; Mahenthiralingam, E. Molecular mechanisms of chlorhexidine tolerance in Burkholderia cenocepacia biofilms. Antimicrob. Agents Chemother. 2011, 55, 1912–1919. [Google Scholar] [CrossRef] [PubMed]
- Menard, A.; de los Santos, P.E.; Arnault Graindorge, V.; Cournoyer, B.; Genomics, B.; Cournoyer, A. Architecture of Burkholderia cepacia complex sigma 70 gene family: Evidence of alternative primary and clade-specific factors, and genomic instability Architecture 70 alternative. BMC Genom. 2007, 8, 308. [Google Scholar] [CrossRef] [PubMed]
- Raina, S.; Missiakas, D.; Georgopoulos, C. The rpoE gene encoding the sigma E (sigma 24) heat shock sigma factor of Escherichia coli. EMBO J. 1995, 14, 1043–1055. [Google Scholar] [PubMed]
- Flannagan, R.S.; Valvano, M.A. Burkholderia cenocepacia requires RpoE for growth under stress conditions and delay of phagolysosomal fusion in macrophages. Microbiology 2008, 154, 643–653. [Google Scholar] [CrossRef] [PubMed]
- Lardi, M.; Aguilar, C.; Pedrioli, A.; Omasits, U.; Suppiger, A.; Cárcamo-Oyarce, G.; Schmid, N.; Ahrens, C.H.; Eberl, L.; Pessi, G. σ54-Dependent Response to Nitrogen Limitation and Virulence in Burkholderia cenocepacia Strain H111. Appl. Environ. Microbiol. 2015, 81, 4077–4089. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, A.S.; Leitão, J.H.; Silva, I.N.; Pinheiros, P.F.; Sousa, S.A.; Ramos, C.G.; Moreira, L.M. Distribution of cepacian biosynthesis genes among environmental and clinical Burkholderia strains and role of cepacian exopolysaccharide in resistance to stress conditions. Appl. Environ. Microbiol. 2010, 76, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Conway, B.-A.D.; Chu, K.K.; Bylund, J.; Altman, E.; Speert, D.P. Production of exopolysaccharide by Burkholderia cenocepacia results in altered cell-surface interactions and altered bacterial clearance in mice. J. Infect. Dis. 2004, 190, 957–966. [Google Scholar] [CrossRef] [PubMed]
- Sousa, S.A.; Ulrich, M.; Bragonzi, A.; Burke, M.; Worlitzsch, D.; Leitão, J.H.; Meisner, C.; Eberl, L.; Sá-correia, I.; Döring, G. Virulence of Burkholderia cepacia complex strains in gp91phox-/-mice. Cell. Microbiol. 2007, 9, 2817–2825. [Google Scholar] [CrossRef] [PubMed]
- Bylund, J.; Burgess, L.-A.; Cescutti, P.; Ernst, R.K.; Speert, D.P. Exopolysaccharides from Burkholderia cenocepacia inhibit neutrophil chemotaxis and scavenge reactive oxygen species. J. Biol. Chem. 2006, 281, 2526–2532. [Google Scholar] [CrossRef] [PubMed]
- Cunha, M.V.; Sousa, S.A.; Leitão, J.H.; Moreira, L.M.; Videira, P.A.; Sá-Correia, I. Studies on the involvement of the exopolysaccharide produced by cystic fibrosis-associated isolates of the Burkholderia cepacia complex in biofilm formation and in persistence of respiratory infections. J. Clin. Microbiol. 2004, 42, 3052–3058. [Google Scholar] [CrossRef] [PubMed]
- Saldías, M.S.; Lamothe, J.; Wu, R.; Valvano, M.A. Burkholderia cenocepacia requires the RpoN sigma factor for biofilm formation and intracellular trafficking within macrophages. Infect. Immun. 2008, 76, 1059–1067. [Google Scholar] [CrossRef] [PubMed]
- Hunt, T.A.; Kooi, C.; Sokol, P.A.; Valvano, M.A. Identification of Burkholderia cenocepacia genes required for bacterial survival in vivo. Infect. Immun. 2004, 72, 4010–4022. [Google Scholar] [CrossRef] [PubMed]
- Eberl, L. Quorum sensing in the genus Burkholderia. Int. J. Med. Microbiol. 2006, 296, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Lewenza, S.; Conway, B.; Greenberg, E.P.; Sokol, P.A. Quorum sensing in Burkholderia cepacia: Identification of the LuxRI homologs CepRI. J. Bacteriol. 1999, 181, 748–756. [Google Scholar] [PubMed]
- Gotschlich, A.; Huber, B.; Geisenberger, O.; Tögl, A.; Steidle, A.; Riedel, K.; Hill, P.; Tümmler, B.; Vandamme, P.; Middleton, B.; et al. Synthesis of Multiple N-Acylhomoserine Lactones is Wide-spread Among the Members of the Burkholderia cepacia Complex. Syst. Appl. Microbiol. 2001, 24, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Lutter, E.; Lewenza, S.; Dennis, J.J.; Visser, M.B.; Sokol, P.A. Distribution of Quorum-Sensing Genes in the Burkholderia cepacia Complex. Infect. Immun. 2001, 69, 4661–4666. [Google Scholar] [CrossRef] [PubMed]
- Uehlinger, S.; Schwager, S.; Bernier, S.P.; Riedel, K.; Nguyen, D.T.; Sokol, P.A.; Eberl, L. Identification of specific and universal virulence factors in Burkholderia cenocepacia strains by using multiple infection hosts. Infect. Immun. 2009, 77, 4102–4110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sokol, P.A.; Sajjan, U.; Visser, M.B.; Gingues, S.; Forstner, J.; Kooi, C. The CepIR quorum-sensing system contributes to the virulence of Burkholderia cenocepacia respiratory infections. Microbiology 2003, 149, 3649–3658. [Google Scholar] [CrossRef] [PubMed]
- Subramoni, S.; Sokol, P.A. Quorum sensing systems influence Burkholderia cenocepacia virulence. Future Microbiol. 2012, 7, 1373–1387. [Google Scholar] [CrossRef] [PubMed]
- Venturi, V.; Friscina, A.; Bertani, I.; Devescovi, G.; Aguilar, C. Quorum sensing in the Burkholderia cepacia complex. Res. Microbiol. 2004, 155, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Suppiger, A.; Schmid, N.; Aguilar, C.; Pessi, G.; Eberl, L. Two quorum sensing systems control biofilm formation and virulence in members of the Burkholderia cepacia complex. Virulence 2013, 4, 400–409. [Google Scholar] [CrossRef] [PubMed]
- Conway, B.-A.; Greenberg, E.P. Quorum-sensing signals and quorum-sensing genes in Burkholderia vietnamiensis. J. Bacteriol. 2002, 184, 1187–1191. [Google Scholar] [CrossRef] [PubMed]
- Boon, C.; Deng, Y.; Wang, L.-H.; He, Y.; Xu, J.-L.; Fan, Y.; Pan, S.Q.; Zhang, L.-H. A novel DSF-like signal from Burkholderia cenocepacia interferes with Candida albicans morphological transition. ISME J. 2008, 2, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Wu, J.; Eberl, L.; Zhang, L.-H. Structural and Functional Characterization of Diffusible Signal Factor Family Quorum-Sensing Signals Produced by Members of the Burkholderia cepacia Complex. Appl. Environ. Microbiol. 2010, 76, 4675–4683. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Schmid, N.; Wang, C.; Wang, J.; Pessi, G.; Wu, D.; Lee, J.; Aguilar, C.; Ahrens, C.H.; Chang, C.; et al. Cis-2-dodecenoic acid receptor RpfR links quorum-sensing signal perception with regulation of virulence through cyclic dimeric guanosine monophosphate turnover. Proc. Natl. Acad. Sci. USA 2012, 109, 15479–15484. [Google Scholar] [CrossRef] [PubMed]
- Ryan, R.P.; Dow, J.M. Communication with a growing family: Diffusible signal factor (DSF) signaling in bacteria. Trends Microbiol. 2011, 19, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Twomey, K.B.; O’Connell, O.J.; McCarthy, Y.; Dow, J.M.; O’Toole, G.A.; Plant, B.J.; Ryan, R.P. Bacterial cis-2-unsaturated fatty acids found in the cystic fibrosis airway modulate virulence and persistence of Pseudomonas aeruginosa. ISME J. 2012, 6, 939–950. [Google Scholar] [CrossRef] [PubMed]
- Malott, R.J.; Baldwin, A.; Mahenthiralingam, E.; Sokol, P.A. Characterization of the cciIR Quorum-Sensing System in Burkholderia cenocepacia. Infect. Immun. 2005, 73, 4982–4992. [Google Scholar] [CrossRef] [PubMed]
- Ryan, G.T.; Wei, Y.; Winans, S.C. A LuxR-type repressor of Burkholderia cenocepacia inhibits transcription via antiactivation and is inactivated by its cognate acylhomoserine lactone. Mol. Microbiol. 2013, 87, 94–111. [Google Scholar] [CrossRef] [PubMed]
- Bernier, S.P.; Nguyen, D.T.; Sokol, P.A. A LysR-type transcriptional regulator in Burkholderia cenocepacia influences colony morphology and virulence. Infect. Immun. 2008, 76, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Aubert, D.F.; O’Grady, E.P.; Hamad, M.A.; Sokol, P.A.; Valvano, M.A. The Burkholderia cenocepacia sensor kinase hybrid AtsR is a global regulator modulating quorum-sensing signalling. Environ. Microbiol. 2013, 15, 372–385. [Google Scholar] [CrossRef] [PubMed]
- Khodai-Kalaki, M.; Aubert, D.F.; Valvano, M.A. Characterization of the AtsR hybrid sensor kinase phosphorelay pathway and identification of its response regulator in Burkholderia cenocepacia. J. Biol. Chem. 2013, 288, 30473–30484. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.S. Iron acquisition mechanisms of the Burkholderia cepacia complex. BioMetals 2007, 20, 431–452. [Google Scholar] [CrossRef] [PubMed]
- Visser, M.B.; Majumdar, S.; Hani, E.; Sokol, P.A. Importance of the ornibactin and pyochelin siderophore transport systems in Burkholderia cenocepacia lung infections. Infect. Immun. 2004, 72, 2850–2857. [Google Scholar] [CrossRef] [PubMed]
- Sokol, P.A.; Darling, P.; Lewenza, S.; Corbett, C.R.; Kooi, C.D. Identification of a siderophore receptor required for ferric ornibactin uptake in Burkholderia cepacia. Infect. Immun. 2000, 68, 6554–6560. [Google Scholar] [CrossRef] [PubMed]
- Tyrrell, J.; Whelan, N.; Wright, C.; Sá-Correia, I.; McClean, S.; Thomas, M.; Callaghan, M. Investigation of the multifaceted iron acquisition strategies of Burkholderia cenocepacia. BioMetals 2015, 28, 367–380. [Google Scholar] [CrossRef] [PubMed]
- Whitby, P.W.; VanWagoner, T.M.; Springer, J.M.; Morton, D.J.; Seale, T.W.; Stull, T.L. Burkholderia cenocepacia utilizes ferritin as an iron source. J. Med. Microbiol. 2006, 55, 661–668. [Google Scholar] [CrossRef] [PubMed]
- Mathew, A.; Eberl, L.; Carlier, A.L. A novel siderophore-independent strategy of iron uptake in the genus Burkholderia. Mol. Microbiol. 2014, 91, 805–820. [Google Scholar] [CrossRef] [PubMed]
- Bevivino, A.; Dalmastri, C.; Tabacchioni, S.; Chiarini, L.; Belli, M.L.; Piana, S.; Materazzo, A.; Vandamme, P.; Manno, G. Burkholderia cepacia complex bacteria from clinical and environmental sources in Italy: Genomovar status and distribution of traits related to virulence and transmissibility. J. Clin. Microbiol. 2002, 40, 846–851. [Google Scholar] [CrossRef] [PubMed]
- Hutchison, M.L.; Poxton, I.R.; Govan, J.R. Burkholderia cepacia produces a hemolysin that is capable of inducing apoptosis and degranulation of mammalian phagocytes. Infect. Immun. 1998, 66, 2033–2039. [Google Scholar] [PubMed]
- Thomson, E.L.S.; Dennis, J.J. A Burkholderia cepacia complex non-ribosomal peptide-synthesized toxin is hemolytic and required for full virulence. Virulence 2012, 3, 286–298. [Google Scholar] [CrossRef] [PubMed]
- Ramos, J.L.; Martínez-Bueno, M.; Molina-Henares, A.J.; Terán, W.; Watanabe, K.; Zhang, X.; Gallegos, M.T.; Brennan, R.; Tobes, R. The TetR family of transcriptional repressors. Microbiol. Mol. Biol. Rev. 2005, 69, 326–356. [Google Scholar] [CrossRef] [PubMed]
- Winsor, G.L.; Khaira, B.; van Rossum, T.; Lo, R.; Whiteside, M.D.; Brinkman, F.S.L. The Burkholderia Genome Database: Facilitating flexible queries and comparative analyses. Bioinformatics 2008, 24, 2803–2804. [Google Scholar] [CrossRef] [PubMed]
- O’Grady, E.P.; Nguyen, D.T.; Weisskopf, L.; Eberl, L.; Sokol, P.A. The Burkholderia cenocepacia LysR-type transcriptional regulator ShvR influences expression of quorum-sensing, protease, type II secretion, and afc genes. J. Bacteriol. 2011, 193, 163–176. [Google Scholar] [CrossRef] [PubMed]
- Thomson, E.L.S.; Dennis, J.J. Common duckweed (Lemna minor) is a versatile high-throughput infection model for the Burkholderia cepacia complex and other pathogenic bacteria. PLoS ONE 2013, 8, e80102. [Google Scholar] [CrossRef] [PubMed]
- Subramoni, S.; Agnoli, K.; Eberl, L.; Lewenza, S.; Sokol, P.A. Role of Burkholderia cenocepacia afcE and afcF genes in determining lipid-metabolism-associated phenotypes. Microbiology 2013, 159, 603–614. [Google Scholar] [CrossRef] [PubMed]
- Michalska, K.; Chhor, G.; Clancy, S.; Jedrzejczak, R.; Babnigg, G.; Winans, S.C.; Joachimiak, A. RsaM: A transcriptional regulator of Burkholderia spp. with novel fold. FEBS J. 2014, 281, 4293–4306. [Google Scholar] [CrossRef] [PubMed]
- Stock, A.M.; Robinson, V.L.; Goudreau, P.N. Two-Component Signal Transduction. Annu. Rev. Biochem. 2000, 69, 183–215. [Google Scholar] [CrossRef] [PubMed]
- Merry, C.R.; Perkins, M.; Mu, L.; Peterson, B.K.; Knackstedt, R.W.; Weingart, C.L. Characterization of a Novel Two-Component System in Burkholderia cenocepacia. Curr. Microbiol. 2015, 70, 556–561. [Google Scholar] [CrossRef] [PubMed]
- Gislason, A.S.; Choy, M.; Bloodworth, R.A.M.; Qu, W.; Stietz, M.S.; Li, X.; Zhang, C.; Cardona, S.T. Competitive growth enhances conditional growth mutant sensitivity to antibiotics and exposes a two-component system as an emerging antibacterial target in Burkholderia cenocepacia. Antimicrob. Agents Chemother. 2016, 61, E00790-16. [Google Scholar] [CrossRef] [PubMed]
- Aubert, D.F.; Valvano, M.A.; Hu, S. Quantification of type VI secretion system activity in macrophages infected with Burkholderia cenocepacia. Microbiology 2015, 161, 2161–2173. [Google Scholar] [CrossRef] [PubMed]
- Flannagan, R.S.; Aubert, D.; Kooi, C.; Sokol, P.A.; Valvano, M.A. Burkholderia cenocepacia requires a periplasmic HtrA protease for growth under thermal and osmotic stress and for survival in vivo. Infect. Immun. 2007, 75, 1679–1689. [Google Scholar] [CrossRef] [PubMed]
- Schaefers, M.M.; Liao, T.L.; Boisvert, N.M.; Roux, D.; Yoder-Himes, D.; Priebe, G.P. An Oxygen-Sensing Two-Component System in the Burkholderia cepacia Complex Regulates Biofilm, Intracellular Invasion, and Pathogenicity. PLoS Pathog. 2017, 13, e1006116. [Google Scholar] [CrossRef] [PubMed]
- Wen, J.; Fozo, E. sRNA Antitoxins: More than One Way to Repress a Toxin. Toxins 2014, 6, 2310–2335. [Google Scholar] [CrossRef] [PubMed]
- Lobato-Márquez, D.; Díaz-Orejas, R.; García-del Portillo, F. Toxin-antitoxins and bacterial virulence. FEMS Microbiol. Rev. 2016, 40, 592–609. [Google Scholar] [CrossRef] [PubMed]
- Gottesman, S. Micros for microbes: Non-coding regulatory RNAs in bacteria. Trends Genet. 2005, 21, 399–404. [Google Scholar] [CrossRef] [PubMed]
- Papenfort, K.; Vogel, J. Regulatory RNA in Bacterial Pathogens. Cell Host Microbe 2010, 8, 116–127. [Google Scholar] [CrossRef] [PubMed]
- Feliciano, J.R.; Grilo, A.M.; Guerreiro, S.I.; Sousa, S.A.; Leitão, J.H. Hfq: A multifaceted RNA chaperone involved in virulence. Future Microbiol. 2016, 11, 137–151. [Google Scholar] [CrossRef] [PubMed]
- Updegrove, T.B.; Zhang, A.; Storz, G. Hfq: The flexible RNA matchmaker. Curr. Opin. Microbiol. 2016, 30, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Sousa, S.A.; Ramos, C.G.; Moreira, L.M.; Leitão, J.H. The hfq gene is required for stress resistance and full virulence of Burkholderia cepacia to the nematode Caenorhabditis elegans. Microbiology 2010, 156, 896–908. [Google Scholar] [CrossRef] [PubMed]
- Ramos, C.G.; Grilo, A.M.; da Costa, P.J.P.; Leitão, J.H. Experimental identification of small non-coding regulatory RNAs in the opportunistic human pathogen Burkholderia cenocepacia J2315. Genomics 2013, 101, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Ramos, C.G.; da Costa, P.J.P.; Döring, G.; Leitão, J.H. The Novel Cis-Encoded Small RNA h2cR Is a Negative Regulator of hfq2 in Burkholderia cenocepacia. PLoS ONE 2012, 7, e47896. [Google Scholar] [CrossRef] [PubMed]
- Ramos, C.G.; Grilo, A.M.; Sousa, S.A.; Feliciano, J.R.; Da Costa, P.J.P.; Leitão, J.H. Regulation of Hfq mRNA and protein levels in Escherichia coli and Pseudomonas aeruginosa by the Burkholderia cenocepacia MtvR sRNA. PLoS ONE 2014, 9, e98813. [Google Scholar] [CrossRef] [PubMed]
- Kimelman, A.; Levy, A.; Sberro, H.; Kidron, S.; Leavitt, A.; Amitai, G.; Yoder-Himes, D.R.; Wurtzel, O.; Zhu, Y.; Rubin, E.M.; et al. A vast collection of microbial genes that are toxic to bacteria. Genome Res. 2012, 22, 802–809. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Experiment | Major Findings | Reference |
|---|---|---|
| Genomics | ||
| Retrospective study of a B. dolosa outbreak among CF patients by sequencing the genomes of a single colony of 112 isolates collected from 14 individuals over 16 years. | 17 bacterial genes acquired non-synonymous mutations in multiple individuals, indicating parallel adaptive evolution. Mutated genes involved in antibiotic resistance, bacterial membrane composition, and oxygen-dependent gene expression regulation. | Lieberman et al., 2011 [13] |
| Analysis of intraspecies diversity in the sputum of five CF patients infected with B. dolosa. | Several bacterial lineages coexisted within the same patient. Genes subject to strongest selection are involved in outer membrane component synthesis, iron scavenging and antibiotic resistance. | Lieberman et al., 2014 [14] |
| Genomic and functional evolution analysis of 22 B. multivorans isolates sequentially collected over 20 years. | Population diversified into four primary, coexisting clades, with distinct evolutionary dynamics. Strong selection of genes involved in B. multivorans populations during long-term colonization of CF patient lung targets adherence, metabolism, and cell envelope changes related to adaptation to the biofilm lifestyle. | Silva et al., 2016 [15] |
| Genome-wide comparative analysis of two B. contaminans isolates, from sputum and from blood culture. | The sputum isolate differs from the bloodstream isolate by over 1400 mutations as a result of a mismatch repair-deficient hypermutable state. | Nunvar et al., 2016 [16] |
| Transcriptomics | ||
| Comparative transcriptomics of two clonal isolates of B. cenocepacia in different stages of chronic infection, recovered from a long-term colonized CF patient deceased with cepacia syndrome. | The later isolate presented upregulated genes involved in translation, iron acquisition, efflux of drugs, and adhesion to respiratory epithelial surface. Alterations related to adaptation to the nutritional environment of the CF lung and to oxygen-limitation. | Mira et al., 2011 [17] |
| Comparative transcriptomics of two pan-resistant ET12 outbreak isolates recovered two decades after J2315. | The outbreak strains exhibited downregulation of genes involved in flagella production and chemotaxis; upregulation of genes involved in transport and efflux, restriction modification, and transposition. | Sass et al., 2011 [18] |
| Comparative transcriptomics of B. cenocepacia isolates from the bloodstream of CF patients with cepacia syndrome with isolates recovered from sputum one to two months before the cepacia syndrome. | Blood isolates presented a higher level of expression of virulence genes involved in type III secretion, exopolysaccharide cepacian biosynthesis, and quorum sensing, and reduced expression of flagellar genes. | Kalferstova et al., 2015 [19] |
| Comparative transcriptomics of two B. contaminans isolates from sputum and blood culture of a CF patient. | Differential expression of quorum sensing-regulated virulence factors, motility, and chemotaxis; the low-oxygen-activated (lxa) locus encoding stress-related proteins; and two clusters responsible for the biosynthesis of the antifungal and hemolytic compounds pyrrolnitrin and occidiofungin. | Nunvar et al., 2016 [16] |
© 2017 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sousa, S.A.; Feliciano, J.R.; Pita, T.; Guerreiro, S.I.; Leitão, J.H. Burkholderia cepacia Complex Regulation of Virulence Gene Expression: A Review. Genes 2017, 8, 43. https://doi.org/10.3390/genes8010043
Sousa SA, Feliciano JR, Pita T, Guerreiro SI, Leitão JH. Burkholderia cepacia Complex Regulation of Virulence Gene Expression: A Review. Genes. 2017; 8(1):43. https://doi.org/10.3390/genes8010043
Chicago/Turabian StyleSousa, Sílvia A., Joana R. Feliciano, Tiago Pita, Soraia I. Guerreiro, and Jorge H. Leitão. 2017. "Burkholderia cepacia Complex Regulation of Virulence Gene Expression: A Review" Genes 8, no. 1: 43. https://doi.org/10.3390/genes8010043
APA StyleSousa, S. A., Feliciano, J. R., Pita, T., Guerreiro, S. I., & Leitão, J. H. (2017). Burkholderia cepacia Complex Regulation of Virulence Gene Expression: A Review. Genes, 8(1), 43. https://doi.org/10.3390/genes8010043