Impact of Lateral Transfers on the Genomes of Lepidoptera


Abstract
:1. Introduction
2. Bacterial Sequences Acquired in Lepidopteran Genomes
3. Transfer of Mobile Elements in Lepidoptera
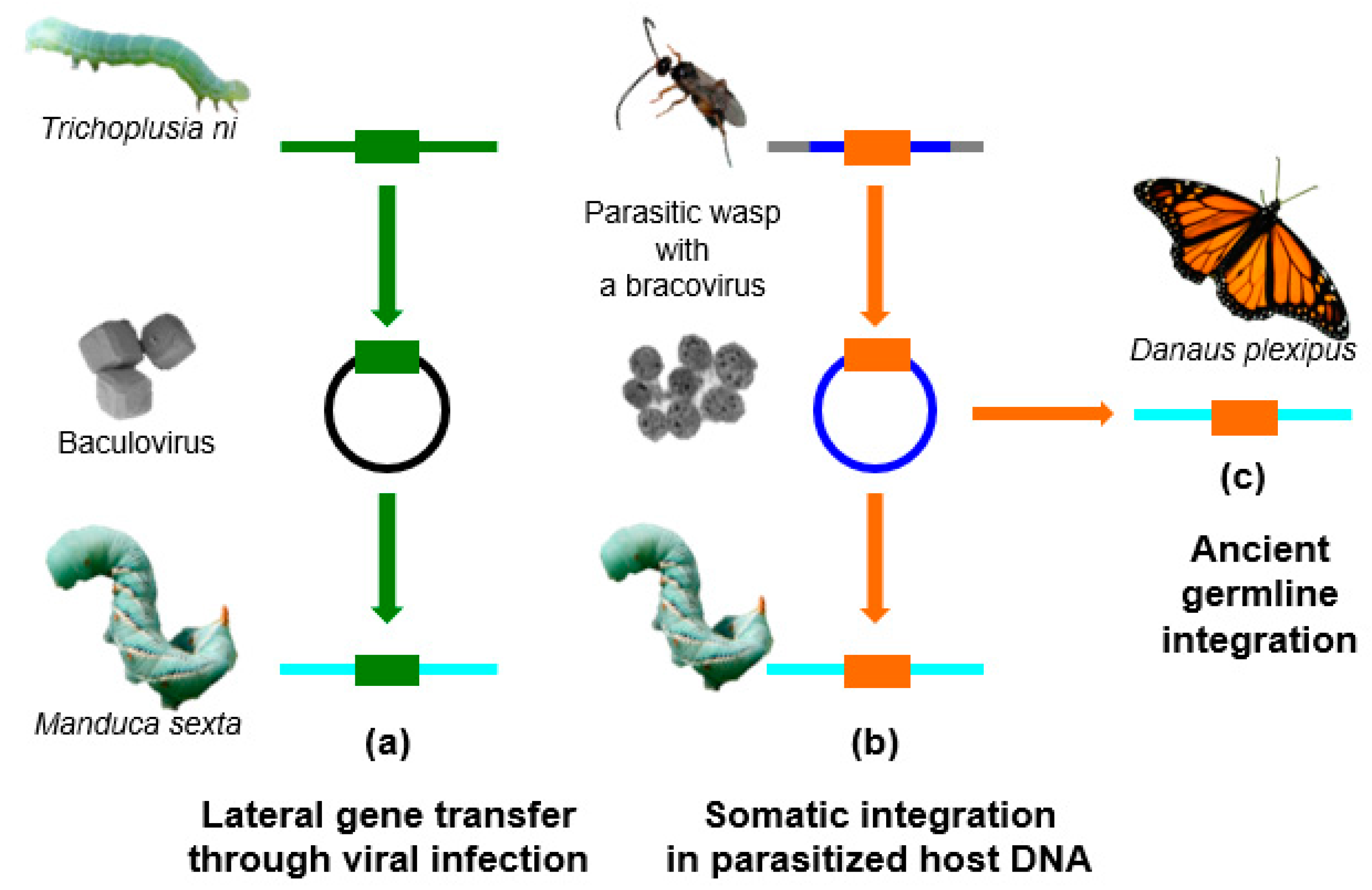
4. Baculoviruses and Horizontal Transfers in Lepidoptera
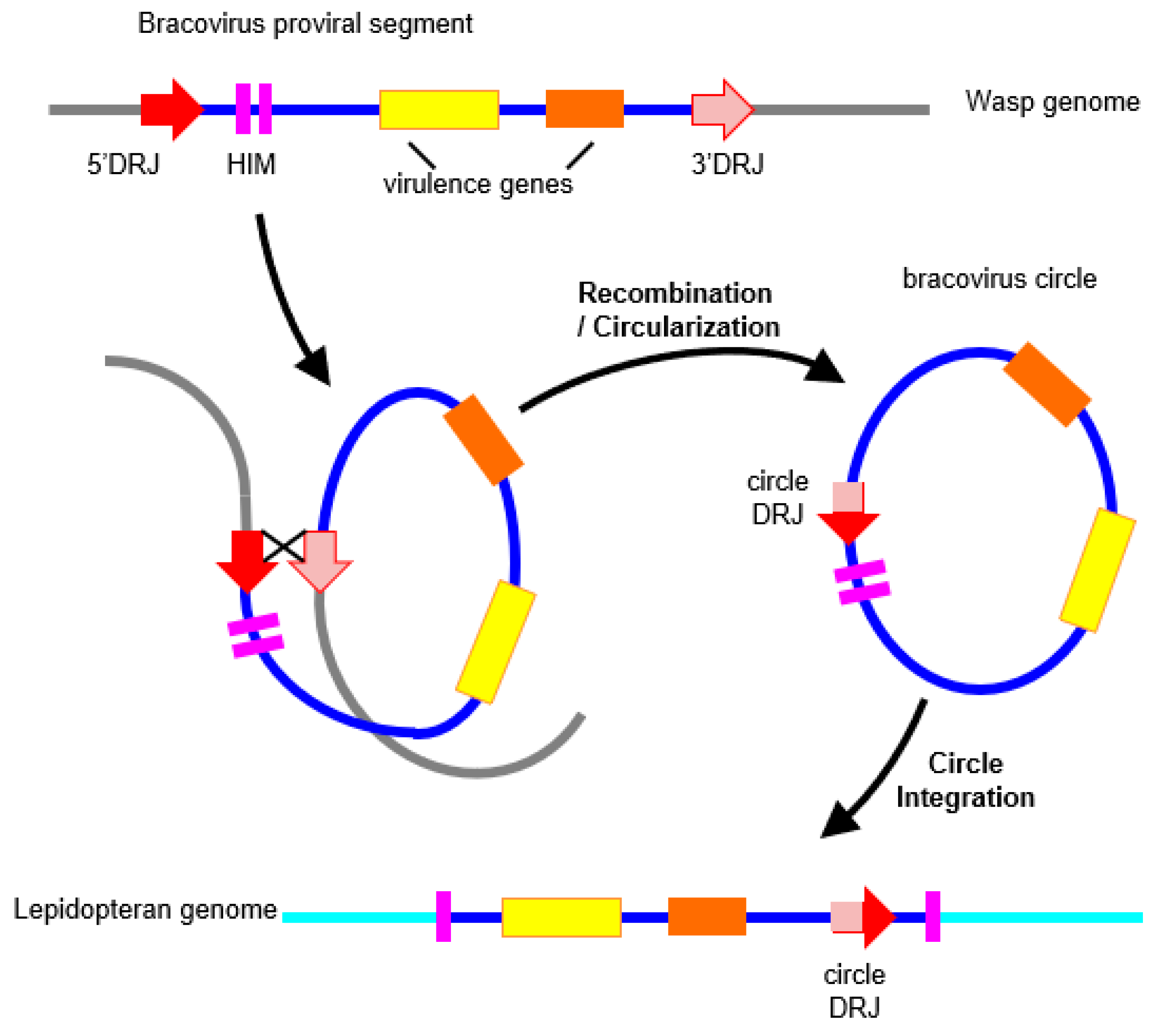
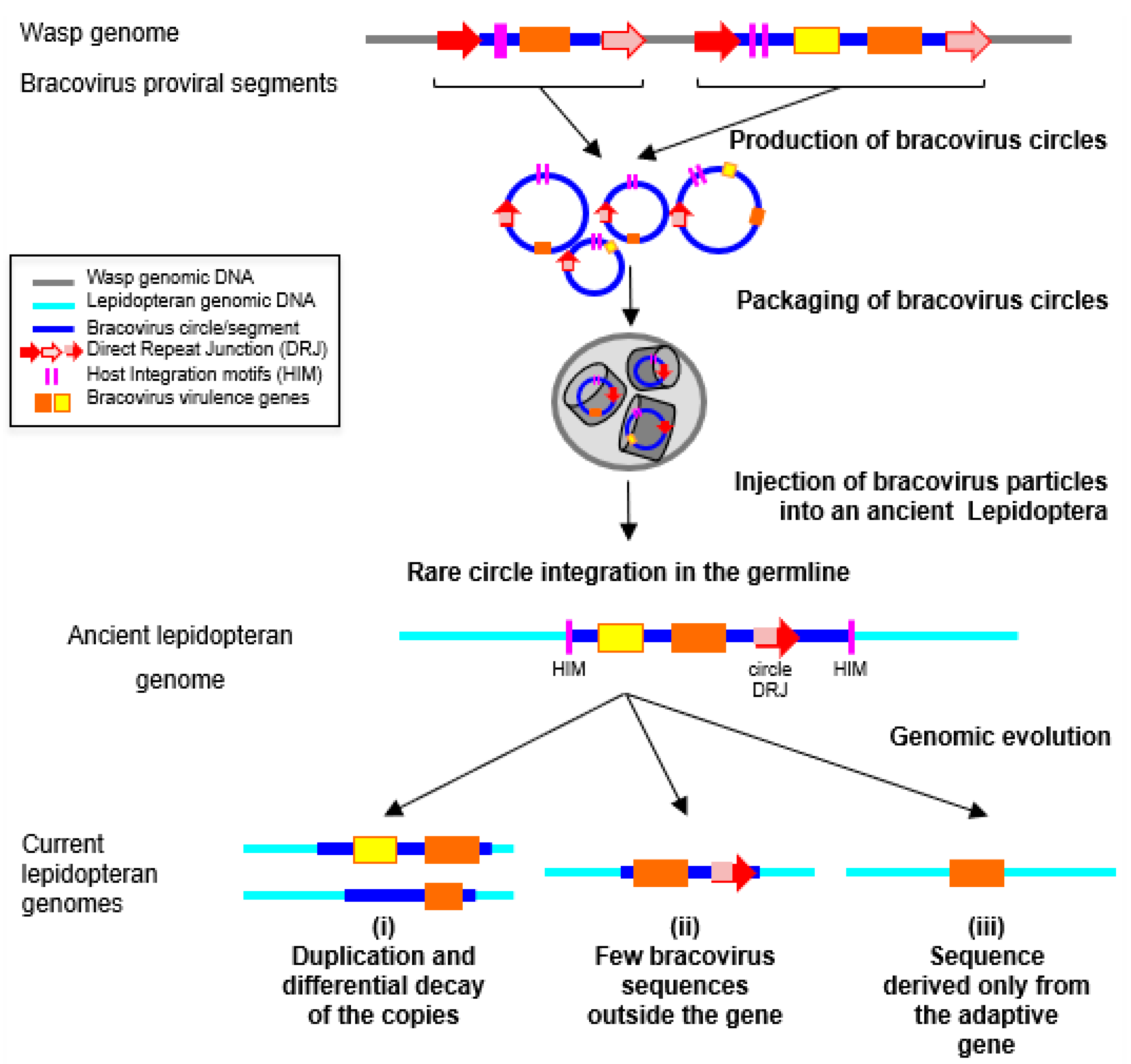
5. Bracoviruses and HGTs in Lepidoptera
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Boto, L. Horizontal gene transfer in the acquisition of novel traits by metazoans. Proc. Biol. Sci. 2014, 281, 20132450. [Google Scholar] [CrossRef] [PubMed]
- Crisp, A.; Boschetti, C.; Perry, M.; Tunnacliffe, A.; Micklem, G. Expression of multiple horizontally acquired genes is a hallmark of both vertebrate and invertebrate genomes. Genome Biol. 2015, 16, 50. [Google Scholar] [CrossRef] [PubMed]
- Keeling, P.J.; Palmer, J.D. Horizontal gene transfer in eukaryotic evolution. Nat. Rev. Genet. 2008, 9, 605–618. [Google Scholar] [CrossRef] [PubMed]
- Syvanen, M. Evolutionary implications of horizontal gene transfer. Annu. Rev. Genet. 2012, 46, 341–358. [Google Scholar] [CrossRef] [PubMed]
- Danchin, E.G. Lateral gene transfer in eukaryotes: Tip of the iceberg or of the ice cube? BMC Biol. 2016, 14, 101. [Google Scholar] [CrossRef] [PubMed]
- Ronsseray, S.; Lehmann, M.; Periquet, G. Comparison of the regulation of P elements in M and M′ strains of Drosophila melanogaster. Genet. Res. 1989, 54, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Feschotte, C.; Gilbert, C. Endogenous viruses: Insights into viral evolution and impact on host biology. Nat. Rev. Genet. 2012, 13, 283–296. [Google Scholar] [CrossRef] [PubMed]
- Thézé, J.; Leclercq, S.; Moumen, B.; Cordaux, R.; Gilbert, C. Remarkable diversity of endogenous viruses in a crustacean genome. Genome Biol. Evol. 2014, 6, 2129–2140. [Google Scholar] [CrossRef] [PubMed]
- Katzourakis, A.; Gifford, R.J. Endogenous viral elements in animal genomes. PLoS Genet. 2010, 6, e1001191. [Google Scholar] [CrossRef] [PubMed]
- Ryabov, E.V. Invertebrate RNA virus diversity from a taxonomic point of view. J. Invertebr. Pathol. 2017, 147, 37–50. [Google Scholar] [CrossRef] [PubMed]
- Lavialle, C.; Cornelis, G.; Dupressoir, A.; Esnault, C.; Heidmann, O.; Vernochet, C.; Heidmann, T. Paleovirology of ‘syncytins’, retroviral env genes exapted for a role in placentation. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2013, 368, 20120507. [Google Scholar] [CrossRef] [PubMed]
- Klasson, L.; Kumar, N.; Bromley, R.; Sieber, K.; Flowers, M.; Ott, S.H.; Tallon, L.J.; Andersson, S.G.; Dunning Hotopp, J.C. Extensive duplication of the Wolbachia DNA in chromosome four of Drosophila ananassae. BMC Genom. 2014, 15, 1097. [Google Scholar] [CrossRef] [PubMed]
- Leclercq, S.; Thézé, J.; Chebbi, M.A.; Giraud, I.; Moumen, B.; Ernenwein, L.; Greve, P.; Gilbert, C.; Cordaux, R. Birth of a W sex chromosome by horizontal transfer of Wolbachia bacterial symbiont genome. Proc. Natl. Acad. Sci. USA 2016, 113, 15036–15041. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, C.; Peccoud, J.; Chateigner, A.; Moumen, B.; Cordaux, R.; Herniou, E.A. Continuous influx of genetic material from host to virus populations. PLoS Genet. 2016, 12, e1005838. [Google Scholar] [CrossRef] [PubMed]
- Haegeman, A.; Jones, J.T.; Danchin, E.G. Horizontal gene transfer in nematodes: A catalyst for plant parasitism? Mol. Plant Microbe Interact. 2011, 24, 879–887. [Google Scholar] [CrossRef] [PubMed]
- Kirsch, R.; Gramzow, L.; Theissen, G.; Siegfried, B.D.; Ffrench-Constant, R.H.; Heckel, D.G.; Pauchet, Y. Horizontal gene transfer and functional diversification of plant cell wall degrading polygalacturonases: Key events in the evolution of herbivory in beetles. Insect Biochem. Mol. Biol. 2014, 52, 33–50. [Google Scholar] [CrossRef] [PubMed]
- Bézier, A.; Annaheim, M.; Herbinière, J.; Wetterwald, C.; Gyapay, G.; Bernard-Samain, S.; Wincker, P.; Roditi, I.; Heller, M.; Belghazi, M.; et al. Polydnaviruses of braconid wasps derive from an ancestral nudivirus. Science 2009, 323, 926–930. [Google Scholar] [CrossRef] [PubMed]
- Herniou, E.A.; Huguet, E.; Thézé, J.; Bézier, A.; Periquet, G.; Drezen, J.-M. When parasitc wasps hijacked viruses: Genomic and functionnal evolution of polydnaviruses. Philos. Trans. Royal Soc. B 2013, 368, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Strand, M.R.; Burke, G.R. Polydnavirus-wasp associations: Evolution, genome organization, and function. Curr. Opin. Virol. 2013, 3, 587–594. [Google Scholar] [CrossRef] [PubMed]
- Drezen, J.M.; Leobold, M.; Bézier, A.; Huguet, E.; Volkoff, A.N.; Herniou, E.A. Endogenous viruses of parasitic wasps: Variations on a common theme. Curr. Opin. Virol. 2017, 25, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, J.; Drezen, J.M.; Herniou, E.A. The recurrent domestication of polydnaviruses: Major transitions in parasitic wasps. Parasitology 2017, in press. [Google Scholar]
- Beck, M.H.; Zhang, S.; Bitra, K.; Burke, G.R.; Strand, M.R. The encapsidated genome of Microplitis demolitor bracovirus integrates into the host Pseudoplusia includens. J. Virol. 2011, 85, 11685–11696. [Google Scholar] [CrossRef] [PubMed]
- Gasmi, L.; Boulain, H.; Gauthier, J.; Hua-Van, A.; Musset, K.; Jakubowska, A.K.; Aury, J.M.; Volkoff, A.N.; Huguet, E.; Herrero, S.; et al. Recurrent domestication by lepidoptera of genes from their parasites mediated by bracoviruses. PLoS Genet. 2015, 11, e1005470. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.W.; Shen, Y.H.; Xiang, Z.H.; Zhang, Z. Pathogen-origin horizontally transferred genes contribute to the evolution of lepidopteran insects. BMC Evol. Biol. 2011, 11, 356. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Lou, M.M.; Xie, G.L.; Zhang, G.Q.; Zhou, X.P.; Li, B.; Jin, G.L. Horizontal gene transfer in silkworm, Bombyx mori. BMC Genom. 2011, 12, 248. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.F.; Xiao, J.H.; He, S.M.; Liu, L.; Murphy, R.W.; Huang, D.W. Multiple ancient horizontal gene transfers and duplications in lepidopteran species. Insect Mol. Biol. 2013, 22, 72–87. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, D.; Redding, A.J.; Werren, J.H. Characterization of an ancient lepidopteran lateral gene transfer. PLoS ONE 2013, 8, e59262. [Google Scholar] [CrossRef] [PubMed]
- Wybouw, N.; Dermauw, W.; Tirry, L.; Stevens, C.; Grbic, M.; Feyereisen, R.; Van Leeuwen, T. A gene horizontally transferred from bacteria protects arthropods from host plant cyanide poisoning. eLIFE 2014, 3, e02365. [Google Scholar] [CrossRef] [PubMed]
- Hilgenboecker, K.; Hammerstein, P.; Schlattmann, P.; Telschow, A.; Werren, J.H. How many species are infected with Wolbachia? a statistical analysis of current data. FEMS Microbiol. Lett. 2008, 281, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.Z.; Breinholt, J.W.; Kawahara, A.Y. Evidence for common horizontal transmission of Wolbachia among butterflies and moths. BMC Evol. Biol. 2016, 16, 118. [Google Scholar] [CrossRef] [PubMed]
- Kageyama, D.; Nishimura, G.; Hoshizaki, S.; Ishikawa, Y. Feminizing Wolbachia in an insect, Ostrinia furnacalis (Lepidoptera: Crambidae). Heredity 2002, 88, 444–449. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, T.; Ishikawa, H.; Sasaki, T. Infection density of Wolbachia and level of cytoplasmic incompatibility in the mediterranean flour moth, Ephestia kuehniella. J. Invertebr. Pathol. 2003, 84, 1–5. [Google Scholar] [CrossRef]
- Dyson, E.A.; Hurst, G.D. Persistence of an extreme sex-ratio bias in a natural population. Proc. Natl. Acad. Sci. USA 2004, 101, 6520–6523. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.J.; Vavre, F.; Beukeboom, L.W. Manipulation of arthropod sex determination by endosymbionts: Diversity and molecular mechanisms. Sex. Dev. 2014, 8, 59–73. [Google Scholar] [CrossRef] [PubMed]
- Nikoh, N.; Tanaka, K.; Shibata, F.; Kondo, N.; Hizume, M.; Shimada, M.; Fukatsu, T. Wolbachia genome integrated in an insect chromosome: Evolution and fate of laterally transferred endosymbiont genes. Genome Res. 2008, 18, 272–280. [Google Scholar] [CrossRef] [PubMed]
- Klasson, L.; Kambris, Z.; Cook, P.E.; Walker, T.; Sinkins, S.P. Horizontal gene transfer between Wolbachia and the mosquito Aedes aegypti. BMC Genom. 2009, 10, 33. [Google Scholar] [CrossRef] [PubMed]
- Doudoumis, V.; Tsiamis, G.; Wamwiri, F.; Brelsfoard, C.; Alam, U.; Aksoy, E.; Dalaperas, S.; Abd-Alla, A.; Ouma, J.; Takac, P.; et al. Detection and characterization of Wolbachia infections in laboratory and natural populations of different species of tsetse flies (genus Glossina). BMC Microbiol. 2012. [Google Scholar] [CrossRef] [PubMed]
- Aikawa, T.; Anbutsu, H.; Nikoh, N.; Kikuchi, T.; Shibata, F.; Fukatsu, T. Longicorn beetle that vectors pinewood nematode carries many Wolbachia genes on an autosome. Proc. Biol. Sci. 2009, 276, 3791–3798. [Google Scholar] [CrossRef] [PubMed]
- Dunning Hotopp, J.C.; Clark, M.E.; Oliveira, D.C.; Foster, J.M.; Fischer, P.; Munoz Torres, M.C.; Giebel, J.D.; Kumar, N.; Ishmael, N.; Wang, S.; et al. Widespread lateral gene transfer from intracellular bacteria to multicellular eukaryotes. Science 2007, 317, 1753–1756. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Creasy, T.; Sun, Y.; Flowers, M.; Tallon, L.J.; Dunning Hotopp, J.C. Efficient subtraction of insect rRNAprior to transcriptome analysis of Wolbachia-Drosophila lateral gene transfer. BMC Res. Notes 2012, 5, 230. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.Y.; Bubnell, J.E.; Aquadro, C.F. Population genomics of infectious and integrated Wolbachia pipientis genomes in Drosophila ananassae. Genome Biol. Evol. 2015, 7, 2362–2382. [Google Scholar] [CrossRef] [PubMed]
- Kanost, M.R.; Arrese, E.L.; Cao, X.; Chen, Y.R.; Chellapilla, S.; Goldsmith, M.R.; Grosse-Wilde, E.; Heckel, D.G.; Herndon, N.; Jiang, H.; et al. Multifaceted biological insights from a draft genome sequence of the tobacco hornworm moth, Manduca sexta. Insect Biochem. Mol. Biol. 2016, 76, 118–147. [Google Scholar] [CrossRef] [PubMed]
- Wybouw, N.; Pauchet, Y.; Heckel, D.G.; Van Leeuwen, T. Horizontal gene transfer contributes to the evolution of arthropod herbivory. Genome Biol. Evol. 2016, 8, 1785–1801. [Google Scholar] [CrossRef] [PubMed]
- Daimon, T.; Taguchi, T.; Meng, Y.; Katsuma, S.; Mita, K.; Shimada, T. β-Fructofuranosidase genes of the silkworm, Bombyx mori: Insights into enzymatic adaptation of B. mori to toxic alkaloids in mulberry latex. J. Biol. Chem. 2008, 283, 15271–15279. [Google Scholar] [CrossRef] [PubMed]
- Grbic, M.; Van Leeuwen, T.; Clark, R.M.; Rombauts, S.; Rouze, P.; Grbic, V.; Osborne, E.J.; Dermauw, W.; Ngoc, P.C.; Ortego, F.; et al. The genome of Tetranychus urticae reveals herbivorous pest adaptations. Nature 2011, 479, 487–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keeling, C.I.; Yuen, M.M.; Liao, N.Y.; Docking, T.R.; Chan, S.K.; Taylor, G.A.; Palmquist, D.L.; Jackman, S.D.; Nguyen, A.; Li, M.; et al. Draft genome of the mountain pine beetle, Dendroctonus ponderosae hopkins, a major forest pest. Genome Biol. 2013, 14, R27. [Google Scholar] [CrossRef] [PubMed]
- van Ohlen, M.; Herfurth, A.M.; Kerbstadt, H.; Wittstock, U. Cyanide detoxification in an insect herbivore: Molecular identification of β-cyanoalanine synthases from Pieris rapae. Insect Biochem. Mol. Biol. 2016, 70, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.; Katsuma, S.; Daimon, T.; Banno, Y.; Uchino, K.; Sezutsu, H.; Tamura, T.; Mita, K.; Shimada, T. The silkworm mutant lemon (lemon lethal) is a potential insect model for human sepiapterin reductase deficiency. J. Biol. Chem. 2009, 284, 11698–11705. [Google Scholar] [CrossRef] [PubMed]
- Wicker, T.; Sabot, F.; Hua-Van, A.; Bennetzen, J.L.; Capy, P.; Chalhoub, B.; Flavell, A.; Leroy, P.; Morgante, M.; Panaud, O.; et al. A unified classification system for eukaryotic transposable elements. Nat. Rev. Genet. 2007, 8, 973–982. [Google Scholar] [CrossRef] [PubMed]
- Finnegan, D.J. Eukaryotic transposable elements and genome evolution. Trends Genet. 1989, 5, 103–107. [Google Scholar] [CrossRef]
- Feschotte, C.; Pritham, E.J. DNA transposons and the evolution of eukaryotic genomes. Annu. Rev. Genet. 2007, 41, 331–368. [Google Scholar] [CrossRef] [PubMed]
- Cordaux, R.; Batzer, M.A. The impact of retrotransposons on human genome evolution. Nat. Rev. Genet. 2009, 10, 691–703. [Google Scholar] [CrossRef] [PubMed]
- Hua-Van, A.; Le Rouzic, A.; Boutin, T.S.; Filee, J.; Capy, P. The struggle for life of the genome’s selfish architects. Biol. Direct 2011, 6, 19. [Google Scholar] [CrossRef] [PubMed]
- Petrov, D.A.; Fiston-Lavier, A.S.; Lipatov, M.; Lenkov, K.; Gonzalez, J. Population genomics of transposable elements in Drosophila melanogaster. Mol. Biol. Evol. 2011, 28, 1633–1644. [Google Scholar] [CrossRef] [PubMed]
- Schaack, S.; Gilbert, C.; Feschotte, C. Promiscuous DNA: Horizontal transfer of transposable elements and why it matters for eukaryotic evolution. Trends Ecol. Evol. 2010, 25, 537–546. [Google Scholar] [CrossRef] [PubMed]
- Daniels, S.B.; Peterson, K.R.; Strausbaugh, L.D.; Kidwell, M.G.; Chovnick, A. Evidence for horizontal transmission of the P transposable element between Drosophila species. Genetics 1990, 124, 339–355. [Google Scholar] [PubMed]
- Dotto, B.R.; Carvalho, E.L.; Silva, A.F.; Duarte Silva, L.F.; Pinto, P.M.; Ortiz, M.F.; Wallau, G.L. HTT-db: Horizontally transferred transposable elements database. Bioinformatics 2015, 31, 2915–2917. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.C.; Loreto, E.L.; Clark, J.B. Factors that affect the horizontal transfer of transposable elements. Curr. Issues Mol. Biol. 2004, 6, 57–71. [Google Scholar] [PubMed]
- Lampe, D.J.; Churchill, M.E.; Robertson, H.M. A purified Mariner transposase is sufficient to mediate transposition in vitro. EMBO J. 1996, 15, 5470–5479. [Google Scholar] [PubMed]
- Zhang, H.H.; Feschotte, C.; Han, M.J.; Zhang, Z. Recurrent horizontal transfers of Chapaev transposons in diverse invertebrate and vertebrate animals. Genome Biol. Evol. 2014, 6, 1375–1386. [Google Scholar] [CrossRef] [PubMed]
- Peccoud, J.; Loiseau, V.; Cordaux, R.; Gilbert, C. Massive horizontal transfer of transposable elements in insects. Proc. Natl. Acad. Sci. USA 2017, 114, 4721–4726. [Google Scholar] [CrossRef] [PubMed]
- Loreto, E.L.; Carareto, C.M.; Capy, P. Revisiting horizontal transfer of transposable elements in Drosophila. Heredity 2008, 100, 545–554. [Google Scholar] [CrossRef] [PubMed]
- Novikova, O.; Sliwinska, E.; Fet, V.; Settele, J.; Blinov, A.; Woyciechowski, M. Cr1 clade of non-LTR retrotransposons from Maculinea butterflies (lepidoptera: Lycaenidae): Evidence for recent horizontal transmission. BMC Evol. Biol. 2007, 7, 93. [Google Scholar] [CrossRef] [PubMed]
- Sormacheva, I.; Smyshlyaev, G.; Mayorov, V.; Blinov, A.; Novikov, A.; Novikova, O. Vertical evolution and horizontal transfer of cr1 non-LTR retrotransposons and Tc1/Mariner DNA transposons in lepidoptera species. Mol. Biol. Evol. 2012, 29, 3685–3702. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, A.; Han, Z.; Zhang, Z.; Li, F.; Li, X. Characterization of three novel sine families with unusual features in Helicoverpa armigera. PLoS ONE 2012, 7, e31355. [Google Scholar] [CrossRef] [PubMed]
- Yoshiyama, M.; Tu, Z.; Kainoh, Y.; Honda, H.; Shono, T.; Kimura, K. Possible horizontal transfer of a transposable element from host to parasitoid. Mol. Biol. Evol. 2001, 18, 1952–1958. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Zhang, H.H.; Huang, K.; Zhang, X.G.; Han, M.J.; Zhang, Z. Repeated horizontal transfers of four DNA transposons in invertebrates and bats. Mob. DNA 2015, 6, 3. [Google Scholar] [CrossRef] [PubMed]
- Kapitonov, V.V.; Jurka, J. Rolling-circle transposons in eukaryotes. Proc. Natl. Acad. Sci. USA 2001, 98, 8714–8719. [Google Scholar] [CrossRef] [PubMed]
- Coates, B.S. Horizontal transfer of a non-autonomous Helitron among insect and viral genomes. BMC Genom. 2015, 16, 137. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Gao, J.; Li, F.; Wang, J. Evidence of horizontal transfer of non-autonomous lep1 Helitrons facilitated by host-parasite interactions. Sci. Rep. 2014, 4, 5119. [Google Scholar] [CrossRef] [PubMed]
- Thomas, J.; Schaack, S.; Pritham, E.J. Pervasive horizontal transfer of rolling-circle transposons among animals. Genome Biol. Evol. 2010, 2, 656–664. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.H.; Shen, Y.H.; Xiong, X.M.; Han, M.J.; Qi, D.W.; Zhang, X.G. Evidence for horizontal transfer of a recently active Academ transposon. Insect Mol. Biol. 2016, 25, 338–346. [Google Scholar] [CrossRef] [PubMed]
- Venner, S.; Miele, V.; Terzian, C.; Biemont, C.; Daubin, V.; Feschotte, C.; Pontier, D. Ecological networks to unravel the routes to horizontal transposon transfers. PLoS Biol. 2017, 15, e2001536. [Google Scholar] [CrossRef] [PubMed]
- Dupuy, C.; Periquet, G.; Serbielle, C.; Bézier, A.; Louis, F.; Drezen, J.M. Transfer of a chromosomal Maverick to endogenous bracovirus in a parasitoid wasp. Genetica 2011, 139, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, C.; Chateigner, A.; Ernenwein, L.; Barbe, V.; Bézier, A.; Herniou, E.A.; Cordaux, R. Population genomics supports baculoviruses as vectors of horizontal transfer of insect transposons. Nat. Commun. 2014, 5, 3348. [Google Scholar] [CrossRef] [PubMed]
- Murphy, N.; Banks, J.C.; Whitfield, J.B.; Austin, A.D. Phylogeny of the parasitic microgastroid subfamilies (Hymenoptera: Braconidae) based on sequence data from seven genes, with an improved time estimate of the origin of the lineage. Mol. Phylogenet. Evol. 2008, 47, 378–395. [Google Scholar] [CrossRef] [PubMed]
- Slack, J.; Arif, B.M. The baculoviruses occlusion-derived virus: Virion structure and function. Adv. Virus Res. 2007, 69, 99–165. [Google Scholar] [PubMed]
- Rohrmann, G.F. Baculovirus Molecular Biology, 3rd ed.; National Library of Medicine, National Center for Biotechnology Information: Bethesda, MD, USA, 2013. Available online: http://www.ncbi.nlm.nih.gov/books/NBK114593/ (accessed on 12 December 2013).
- Herniou, E.A.; Arif, B.M.; Becnel, J.J.; Blissard, G.W.; Bonning, B.; Harrison, R.; Jehle, J.A.; Theilmann, D.A.; Vlak, J.M. Baculoviridae. In Virus Taxonomy, Ninth Report of the International Committee on Taxonomy of Viruses; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Elsevier Academic Press: London, UK, 2012; pp. 163–173. [Google Scholar]
- Krell, P.J. Passage effect of virus infection in insect cells. Cytotechnology 1996, 20, 125–137. [Google Scholar] [CrossRef] [PubMed]
- Fraser, M.J. The TTAA-specific family of transposable elements: Identification, functional characterization, and utility for transformation of insects. In Insect transgenesis: Methods and Applications; Handler, A.M., James, A.A., Eds.; CRC Press: New York, NY, USA, 2000; pp. 249–268. [Google Scholar]
- Hink, W.F.; Vail, P.V. A plaque assay for titration of alfalfa looper nuclear polyhedrosis virus in cabbage looper (TN-368) cell line. J. Invertebr. Pathol. 1973, 22, 168–174. [Google Scholar] [CrossRef]
- Potter, K.N.; Faulkner, P.; MacKinnon, E.A. Strain selection during serial passage of Trichoplusia ni nuclear polyhedrosis virus. J. Virol. 1976, 18, 1040–1050. [Google Scholar] [PubMed]
- Fraser, M.J.; Hink, W.F. The isolation and characterization of the MP and FP plaque variants of Galleria mellonella nuclear polyhedrosis virus. Virology 1982, 117, 366–378. [Google Scholar] [CrossRef]
- Kumar, S.; Miller, L.K. Effects of serial passage of Autographa californica nuclear polyhedrosis virus in cell culture. Virus Res. 1987, 7, 335–349. [Google Scholar] [CrossRef]
- Miller, D.W.; Miller, L.K. A virus mutant with an insertion of a copia-like transposable element. Nature 1982, 299, 562–564. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.; Terzian, C.; Santamaria, P.; Pelisson, A.; Purd’homme, N.; Bucheton, A. Retroviruses in invertebrates: The Gypsy retrotransposon is apparently an infectious retrovirus of Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 1994, 91, 1285–1289. [Google Scholar] [CrossRef] [PubMed]
- Cary, L.C.; Goebel, M.; Corsaro, B.G.; Wang, H.G.; Rosen, E.; Fraser, M.J. Transposon mutagenesis of baculoviruses: Analysis of Trichoplusia ni transposon IFP2 insertions within the fp-locus of nuclear polyhedrosis viruses. Virology 1989, 172, 156–169. [Google Scholar] [CrossRef]
- Wang, H.G.; Fraser, M.J. TTAA serves as the target site for TFP3 lepidopteran transposon insertions in both nuclear polyhedrosis virus and Trichoplusia ni genomes. Insect Mol. Biol. 1993, 1, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Fraser, M.J.; Cary, L.; Boonvisudhi, K.; Wang, H.G. Assay for movement of lepidopteran transposon IFP2 in insect cells using a baculovirus genome as a target DNA. Virology 1995, 211, 397–407. [Google Scholar] [CrossRef] [PubMed]
- Fraser, M.J.; Ciszczon, T.; Elick, T.; Bauser, C. Precise excision of TTAA-specific lepidopteran transposons Piggybac (IFP2) and Tagalong (TFP3) from the baculovirus genome in cell lines from two species of Lepidoptera. Insect Mol. Biol. 1996, 5, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Friesen, P.D.; Rice, W.C.; Miller, D.W.; Miller, L.K. Bidirectional transcription from a solo long terminal repeat of the retrotransposon TED: Symmetrical RNA start sites. Mol. Cell Biol. 1986, 6, 1599–1607. [Google Scholar] [CrossRef] [PubMed]
- Friesen, P.D.; Nissen, M.S. Gene organization and transcription of TED, a lepidopteran retrotransposon integrated within the baculovirus genome. Mol. Cell Biol. 1990, 10, 3067–3077. [Google Scholar] [CrossRef] [PubMed]
- Schetter, C.; Oellig, C.; Doerfler, W. An insertion of insect cell DNA in the 81-map-unit segment of Autographa californica nuclear polyhedrosis virus DNA. J. Virol. 1990, 64, 1844–1850. [Google Scholar] [PubMed]
- Bauser, C.A.; Elick, T.A.; Fraser, M.J. Characterization of hitchhiker, a transposon insertion frequently associated with baculovirus FP mutants derived upon passage in the TN-368 cell line. Virology 1996, 216, 235–237. [Google Scholar] [CrossRef] [PubMed]
- Beames, B.; Summers, M.D. Comparisons of host cell DNA insertions and altered transcription at the site of insertions in few polyhedra baculovirus mutants. Virology 1988, 162, 206–220. [Google Scholar] [CrossRef]
- Beames, B.; Summers, M.D. Sequence comparison of cellular and viral copies of host cell DNA insertions found in Autographa californica nuclear polyhedrosis virus. Virology 1990, 174, 354–363. [Google Scholar] [CrossRef]
- Carstens, E.B. Identification and nucleotide sequence of the regions of Autographa californica nuclear polyhedrosis virus genome carrying insertion elements derived from Spodoptera frugiperda. Virology 1987, 161, 8–17. [Google Scholar] [CrossRef]
- Jehle, J.A.; Fritsch, E.; Nickel, A.; Huber, J.; Backhaus, H. Tcl4.7: A novel lepidopteran transposon found in Cydia pomonella granulosis virus. Virology 1995, 207, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Jehle, J.A. Transmission of insect transposons into baculovirus genomes: An unusual host-pathogen interaction. In Transgenic Organism Biological and Social Implications; Tomiuk, J., Wöhrmann, K., Sentker, A., Eds.; Birkhäuser Verlag: Basel, Switzerland, 1996; pp. 81–97. [Google Scholar]
- Jehle, J.A.; Nickel, A.; Vlak, J.M.; Backhaus, H. Horizontal escape of the novel Tc1-like lepidopteran transposon TCP3.2 into Cydia pomonella granulovirus. J. Mol. Evol. 1998, 46, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Thézé, J.; Cabodevilla, O.; Palma, L.; Williams, T.; Caballero, P.; Herniou, E.A. Genomic diversity in european Spodoptera exigua multiple nucleopolyhedrovirus isolates. J. Gen. Virol. 2014, 95, 2297–2309. [Google Scholar] [CrossRef] [PubMed]
- Fraser, M.J.; Smith, G.E.; Summers, M.D. Acquisition of host cell DNA sequences by baculoviruses: Relationship between host DNA insertions and FP mutants of Autographa californica and Galleria mellonella nuclear polyhedrosis viruses. J. Virol. 1983, 47, 287–300. [Google Scholar] [PubMed]
- Fraser, M.J.; Brusca, J.S.; Smith, G.E.; Summers, M.D. Transposon-mediated mutagenesis of a baculovirus. Virology 1985, 145, 356–361. [Google Scholar] [CrossRef]
- Wang, H.H.; Fraser, M.J.; Cary, L.C. Transposon mutagenesis of baculoviruses: Analysis of TFP3 lepidopteran transposon insertions at the fp locus of nuclear polyhedrosis viruses. Gene 1989, 81, 97–108. [Google Scholar] [CrossRef]
- O’Reilly, D.R.; Passarelli, A.L.; Goldman, I.F.; Miller, L.K. Characterization of the DA26 gene in a hypervariable region of the Autographa californica nuclear polyhedrosis virus genome. J. Gen. Virol. 1990, 71 Pt 5, 1029–1037. [Google Scholar] [CrossRef] [PubMed]
- Hughes, A.L.; Friedman, R. Genome-wide survey for genes horizontally transferred from cellular organisms to baculoviruses. Mol. Biol. Evol. 2003, 20, 979–987. [Google Scholar] [CrossRef] [PubMed]
- Thézé, J.; Takatsuka, J.; Nakai, M.; Arif, B.; Herniou, E.A. Gene acquisition convergence between entomopoxviruses and baculoviruses. Viruses 2015, 7, 1960–1974. [Google Scholar] [CrossRef] [PubMed]
- Biernat, M.A.; Ros, V.I.; Vlak, J.M.; van Oers, M.M. Baculovirus cyclobutane pyrimidine dimer photolyases show a close relationship with lepidopteran host homologues. Insect Mol. Biol. 2011, 20, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Van Houte, S.; Ros, V.I.; Mastenbroek, T.G.; Vendrig, N.J.; Hoover, K.; Spitzen, J.; van Oers, M.M. Protein tyrosine phosphatase-induced hyperactivity is a conserved strategy of a subset of baculoviruses to manipulate lepidopteran host behavior. PLoS ONE 2012, 7, e46933. [Google Scholar] [CrossRef] [PubMed]
- Aragao-Silva, C.W.; Andrade, M.S.; Ardisson-Araujo, D.M.; Fernandes, J.E.; Morgado, F.S.; Bao, S.N.; Moraes, R.H.; Wolff, J.L.; Melo, F.L.; Ribeiro, B.M. The complete genome of a baculovirus isolated from an insect of medical interest: Lonomia obliqua (Lepidoptera: Saturniidae). Sci. Rep. 2016, 6, 23127. [Google Scholar] [CrossRef] [PubMed]
- Gundersen-Rindal, D.; Dupuy, C.; Huguet, E.; Drezen, J.-M. Parasitoid polydnaviruses: Evolution, pathology and applications. Biocontrol Sci. Technol. 2013, 23, 1–61. [Google Scholar] [CrossRef]
- Thézé, J.; Bézier, A.; Periquet, G.; Drezen, J.M.; Herniou, E.A. Paleozoic origin of insect large dsDNA viruses. Proc. Natl. Acad. Sci. USA 2011, 108, 15931–15935. [Google Scholar] [CrossRef] [PubMed]
- Bézier, A.; Herbinière, J.; Lanzrein, B.; Drezen, J.M. Polydnavirus hidden face: The genes producing virus particles of parasitic wasps. J. Invertebr. Pathol. 2009, 101, 194–203. [Google Scholar] [CrossRef] [PubMed]
- Burke, G.R.; Thomas, S.A.; Eum, J.H.; Strand, M.R. Mutualistic polydnaviruses share essential replication gene functions with pathogenic ancestors. PLoS Pathog. 2013, 9, e1003348. [Google Scholar] [CrossRef] [PubMed]
- Wetterwald, C.; Roth, T.; Kaeslin, M.; Annaheim, M.; Wespi, G.; Heller, M.; Maser, P.; Roditi, I.; Pfister-Wilhelm, R.; Bézier, A.; et al. Identification of bracovirus particle proteins and analysis of their transcript levels at the stage of virion formation. J. Gen. Virol. 2010, 91, 2610–2619. [Google Scholar] [CrossRef] [PubMed]
- Louis, F.; Bézier, A.; Periquet, G.; Ferras, C.; Drezen, J.M.; Dupuy, C. The bracovirus genome of the parasitoid wasp Cotesia congregata is amplified within 13 replication units, including sequences not packaged in the particles. J. Virol. 2013, 87, 9649–9660. [Google Scholar] [CrossRef] [PubMed]
- Burke, G.R.; Strand, M.R. Deep sequencing identifies viral and wasp genes with potential roles in replication of Microplitis demolitor bracovirus. J. Virol. 2012, 86, 3293–3306. [Google Scholar] [CrossRef] [PubMed]
- Burke, G.R.; Simmonds, T.J.; Thomas, S.A.; Strand, M.R. Microplitis demolitor bracovirus proviral loci and clustered replication genes exhibit distinct DNA amplification patterns during replication. J. Virol. 2015, 89, 9511–9523. [Google Scholar] [CrossRef] [PubMed]
- Drezen, J.M.; Chevignon, G.; Louis, F.; Huguet, E. Origin and evolution of symbiotic viruses associated with parasitoid wasps. Curr. Opin. Insect Sci. 2014, 6, 35–43. [Google Scholar] [CrossRef]
- Pasquier-Barre, F.; Dupuy, C.; Huguet, E.; Monteiro, F.; Moreau, A.; Poirie, M.; Drezen, J.M. Polydnavirus replication: The EP1 segment of the parasitoid wasp Cotesia congregata is amplified within a larger precursor molecule. J. Gen. Virol. 2002, 83, 2035–2045. [Google Scholar] [CrossRef] [PubMed]
- Strand, M.R. Polydnavirus gene products that interact with the host immune system. In Parasitoid Viruses Symbionts and Pathogens; Beckage, N.E., Drezen, J.-M., Eds.; Elsevier: San Diego, CA, USA, 2012; pp. 149–161. [Google Scholar]
- Beck, M.H.; Inman, R.B.; Strand, M.R. Microplitis demolitor bracovirus genome segments vary in abundance and are individually packaged in virions. Virology 2007, 359, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Schneider, S.E.; Thomas, J.H. Accidental genetic engineers: Horizontal sequence transfer from parasitoid wasps to their lepidopteran hosts. PLoS ONE 2014, 9, e109446. [Google Scholar] [CrossRef] [PubMed]
- Serbielle, C.; Dupas, S.; Perdereau, E.; Hericourt, F.; Dupuy, C.; Huguet, E.; Drezen, J.M. Evolutionary mechanisms driving the evolution of a large polydnavirus gene family coding for protein tyrosine phosphatases. BMC Evol. Biol. 2012, 12, 253. [Google Scholar] [CrossRef] [PubMed]
- Lushai, G.; Smith, D.A.S.; Goulson, D.; Allen, J.A. Mitochondrial DNA clocks and the phylogeny of Danaus butterflies. Insect Sci. Appl. 2003, 23, 309–315. [Google Scholar] [CrossRef]
- Dushay, M.S.; Beckage, N.E. Dose-dependent separation of Cotesia congregata associated polydnavirus effects on Manduca sexta larval development and immunity. J. Insect Physiol. 1993, 39, 1029–1040. [Google Scholar] [CrossRef]
- Gitau, C.W.; Gundersen-Rindal, D.; Pedroni, M.; Mbugi, P.J.; Dupas, S. Differential expression of the Crv1 haemocyte inactivation-associated polydnavirus gene in the african maize stem borer Busseola fusca (fuller) parasitized by two biotypes of the endoparasitoid Cotesia sesamiae (cameron). J. Insect. Physiol. 2007, 53, 676–684. [Google Scholar] [CrossRef] [PubMed]
- Beckage, N.E.; Tan, F.F. Development of the braconid wasp Cotesia congregata in a semi-permissive noctuid host, Trichoplusia ni. J. Invertebr. Pathol. 2002, 81, 49–52. [Google Scholar] [CrossRef]
- Harwood, S.H.; McElfresh, J.S.; Nguyen, A.; Conlan, C.A.; Beckage, N.E. Production of early expressed parasitism-specific proteins in alternate sphingid hosts of the braconid wasp Cotesia congregata. J. Invertebr. Pathol. 1998, 71, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Wyder, S.; Blank, F.; Lanzrein, B. Fate of polydnavirus DNA of the egg-larval parasitoid Chelonus inanitus in the host Spodoptera littoralis. J. Insect Physiol. 2003, 49, 491–500. [Google Scholar] [CrossRef]
- Chevignon, G.; Thézé, J.; Cambier, S.; Poulain, J.; Da Silva, C.; Bézier, A.; Musset, K.; Moreau, S.J.; Drezen, J.M.; Huguet, E. Functional annotation of Cotesia congregata bracovirus: Identification of the viral genes expressed in parasitized host immune tissues. J. Virol. 2014, 88, 8795–8812. [Google Scholar] [CrossRef] [PubMed]
- Abhiman, S.; Iyer, L.M.; Aravind, L. Ben: A novel domain in chromatin factors and DNA viral proteins. Bioinformatics 2008, 24, 458–461. [Google Scholar] [CrossRef] [PubMed]
- Dai, Q.; Ren, A.; Westholm, J.O.; Serganov, A.A.; Patel, D.J.; Lai, E.C. The BEN domain is a novel sequence-specific DNA-binding domain conserved in neural transcriptional repressors. Genes Dev. 2013, 27, 602–614. [Google Scholar] [CrossRef] [PubMed]
- Bézier, A.; Louis, F.; Jancek, S.; Periquet, G.; Thézé, J.; Gyapay, G.; Musset, K.; Lesobre, J.; Lenoble, P.; Dupuy, C.; et al. Functional endogenous viral elements in the genome of the parasitoid wasp Cotesia congregata: Insights into the evolutionary dynamics of bracoviruses. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2013, 368, 20130047. [Google Scholar] [CrossRef] [PubMed]
- Gasmi, L.; Jakubowska, A.K.; Herrero, S. Gasmin (bv2-5), a polydnaviral-acquired gene in Spodoptera exigua. Trade-off in the defense against bacterial and viral infections. Dev. Comp. Immunol. 2016, 56, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Gasmi, L.; Ferre, J.; Herrero, S. High bacterial agglutination activity in a single-crd c-type lectin from Spodoptera exigua (lepidoptera: Noctuidae). Biosensors 2017, 7. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Gene Name | Potential Function/Phenotype | Potential Donor | Lepidopteran Recipient(s)—Genomic or EST Sequences | References |
|---|---|---|---|---|
| Glycosyl hydrolase GH31 | Assimilation of plant carbohydrates | Enterococcus bacteria | B. mori, D. plexippus, P. xylostella, H. melpomene, C. catalpae, A. edwardsi, S. pyri, G. geneura, H. fraternal, L. griseipennis, C. bicoloraria, N. fimetaria, T. erectarius, E. lisa, E. claudia, G. rufobrunnea, P. populi, A. velutinana, A. assama, S. littoralis, M. sexta | [24,26,27,42,43] |
| β-fructofuranosidase GH32 | Assimilation of plant carbohydrates/overcoming plant defense | Bacteria of several genera | B. mori, D. plexippus, H. melpomene, E. postvittana, C. fumiferana, P. interpunctella, O. nubilis, B. anynana, H. erato, T. ni, H. virescens, S. littoralis, M. sexta | [24,25,26,42,43,44] |
| β-cyanoaline synthase CAS/ cysteine synthase | Amino acid assimilation/overcoming plant defense | Methylobacterium bacteria | B. mori, D. plexippus, H. melpomene, P. rapae, S. cynthia ricini, M. sexta, S. frugiperda, S. littoralis, T. ni, H. virescens, B. anynana | [24,26,28,43,47] |
| Kynureninase | Body coloration/amino acid assimilation/overcoming plant defense | Listeria grayi bacteria | B. mori, D. plexippus, H. melpomene, C. fumiferana, O. nubilis, B. anynana, H. virescens, M. sexta | [24,25,26,42,48] |
| Glucose-1 phosphatase/inositol phosphatase | Glycolysis/Resistance to bacterial pathogens | Bacteria of several genera | B. mori, D. plexippus, H. melpomene, T. ni, H. virescens, S. frugiperda, M. sexta | [24,25,26,42] |
| N-methyltryptophane oxidase | Amino acid degradation/detoxification of malpighian tubules/resistance to bacterial pathogens | Serratia bacteria | B. mori, M. sexta, A. assama | [24,25,26,42] |
| Glycerophosphoryl diester phosphodiesterase | Glycerophospholipid metabolism | Pseudomonas bacteria | B. mori, D. plexippus, H. melpomene, C. fumiferana, O. nubilis, B. anynana, M. brassicae, T. ni, H. virescens, S. littoralis, S. frugiperda, M. sexta, A. assama | [24,25,26,42] |
| Chitinase | Carbohydrate transport and metabolism | Serratia bacteria | B. mori, D. plexippus, H. melpomene, C. fumiferana, P. xuthus, B. anynana, H. erato, H. virescens, S. littoralis, S. frugiperda, M. sexta, A. assama | [24,25,26,42] |
| NAD-dependent epimerase/dehydratase | Nutrient and energy metabolism | Bacteria of several genera | B. mori, D. plexippus, H. melpomene, H. virescens, S. frugiperda, S. cynthia ricini, A. assama | [24,25,26] |
| Aromatic ring-opening dioxygenase LigB subunit | unknown | Fungi, or Naegleria gruberi amebia | B. mori, D. plexippus, H. melpomene, E. postvittana, O. nubilis, T. ni, H. virescens, S. littoralis, S. frugiperda, S. cynthia ricini, A. assama, A. mylitta | [25,26] |
| Alginate lyse/alcohol dehydrogenase | Energy production and conversion | Bacteria of several genera | B. mori, D. plexippus, H. melpomene, E. postvittana, C. fumiferana, O. nubilis, B. anynana, T. ni, H. virescens, S. littoralis, S. frugiperda, S. cynthia ricini, A. assama, A. mylitta, M. sexta | [25,26,42] |
| Gamma-glutamyltranspeptidase | Amino acid transport and metabolism/resistance to bacterial pathogens | Bacteria of several genera | B. mori, D. plexippus, H. melpomene, B. anynana, T. ni, H. virescens, S. frugiperda | [24,25,26] |
| Hypothetical protein | unknown | Lactococcus bacteria | B. mori, H. melpomene, C. fumiferana, S. frugiperda, A. mylitta, M. sexta | [24,25,26,42] |
| Methylated DNA-protein-cysteine methyltransferase | unknown | Bacteria | D. plexippus, H. virescens, S. frugiperda | [26] |
| TE Type or Name * | Class | Order | Baculovirus | Origin | Preferred Insertion Site | TE Extremities | References |
|---|---|---|---|---|---|---|---|
| 5S | I | SINE | AcMNPV | T. ni | - | - | [75] |
| BEL | I | LTR | AcMNPV | T. ni S. exigua | - | - | [14] |
| Copia | I | LTR | AcMNPV | T. ni S. exigua | - | - | [14] |
| Gypsy (TED *) | I | LTR | AcMNPV | T. ni S. exigua | AATG TATAT | - | [14,86,92,93] |
| E | II | TIR | AcMNPV | S. frugiperda | TTAA | CCT/AGG | [94] |
| Harbinger | II | TIR | AcMNPV GmMNPV | T. ni S. exigua | T[A/T]A | GGG/CCC | [14,95] |
| hAT | II | TIR | AcMNPV | T. ni S. exigua | - | - | [14] |
| Helitron | II | Helitron | AcMNPV | S. exigua | - | TC/CTAG | [14,69] |
| IFP1.6 * | II | TIR | AcMNPV | S. frugiperda | TTAA | CCT/AGG | [96,97] |
| IFP2.2 * | II | - | AcMNPV | S. frugiperda | GTTTTTAC | TAC/GTT | [96,97] |
| M5 | II | - | AcMNPV | S. frugiperda | TTAA | CCG/CGG | [98] |
| Tc1-Mariner | II | TIR | AcMNPV | T. ni S. exigua | TA | - | [14,75] |
| CpGV | C. leucotreta C. pomonella | TA | CAG/CTG | [99,100,101] | |||
| Mutator | II | TIR | AcMNPV | T. ni | - | - | [14] |
| MULE | II | TIR | AcMNPV | S. exigua | - | - | [14] |
| P element | II | TIR | AcMNPV | T. ni | - | - | [14] |
| PiggyBac (formerly IFP2) | II | TIR | AcMNPV GmMNPV SeMNPV | T. ni S. exigua | TTAA | CCC/GGG | [14,75,88,102] |
| Sola | II | TIR | AcMNPV | T. ni S. exigua | AT | - | [14] |
| Tagalong (formerly TFP3) | II | TIR | AcMNPV GmMNPV | T. ni | TTAA | CCC/GGG | [103,104,105,106] |
| Transib | II | TIR | AcMNPV | T. ni | CGNCG | - | [14] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drezen, J.-M.; Josse, T.; Bézier, A.; Gauthier, J.; Huguet, E.; Herniou, E.A. Impact of Lateral Transfers on the Genomes of Lepidoptera. Genes 2017, 8, 315. https://doi.org/10.3390/genes8110315
Drezen J-M, Josse T, Bézier A, Gauthier J, Huguet E, Herniou EA. Impact of Lateral Transfers on the Genomes of Lepidoptera. Genes. 2017; 8(11):315. https://doi.org/10.3390/genes8110315
Chicago/Turabian StyleDrezen, Jean-Michel, Thibaut Josse, Annie Bézier, Jérémy Gauthier, Elisabeth Huguet, and Elisabeth Anne Herniou. 2017. "Impact of Lateral Transfers on the Genomes of Lepidoptera" Genes 8, no. 11: 315. https://doi.org/10.3390/genes8110315
APA StyleDrezen, J. -M., Josse, T., Bézier, A., Gauthier, J., Huguet, E., & Herniou, E. A. (2017). Impact of Lateral Transfers on the Genomes of Lepidoptera. Genes, 8(11), 315. https://doi.org/10.3390/genes8110315