The Obscure World of Integrative and Mobilizable Elements, Highly Widespread Elements that Pirate Bacterial Conjugative Systems



Abstract
:1. Introduction
2. Maintenance of IMEs
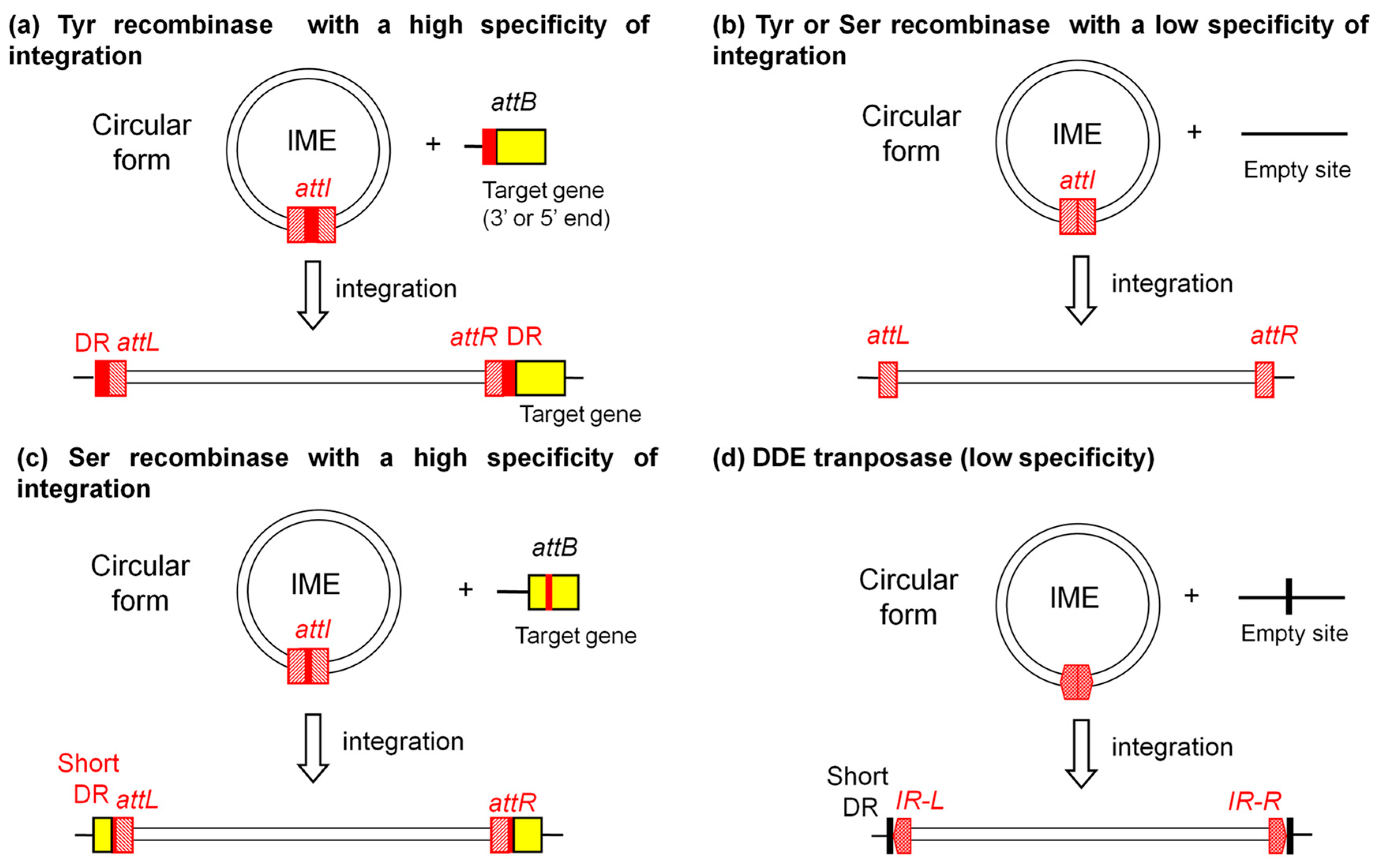
2.1. Integration and Excision
2.2. Maintenance of Excised IMEs: An Unexplored World
2.3. Impact of Other Mobile Genetic Elements on IME Maintenance
3. IMEs: Mobile Elements That Hijack the Conjugative Apparatus of Self-Transmissible Elements
3.1. Canonical IMEs Encoding Their Own Relaxases but no T4SS Protein
3.2. Non-Canonical IMEs Devoid of Relaxases
3.3. Non-Canonical IMEs Encoding Their Own Canonical Relaxase and Some Proteins of the T4SS
3.4. Non-Canonical IMEs Encoding a Non-Canonical Relaxase and/or CP
3.5. IMEs: Harmless Hitchhikers or Harmful Pirates of Conjugative Elements?
4. Moving with IMEs: Their Cargo Genes
4.1. IMEs: A Reservoir of Antibiotic Resistance Genes
4.2. Other Putative Functions Encoded by IMEs
5. Evolution of IMEs
6. IMEs: An Obscure World to Explore
7. Concluding Remarks
Conflicts of Interest
References
- Guglielmini, J.; Quintais, L.; Garcillan-Barcia, M.P.; de la Cruz, F.; Rocha, E.P. The repertoire of ICE in prokaryotes underscores the unity, diversity and ubiquity of conjugation. PLoS Genet. 2011, 7, e1002222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, C.M.; Nielsen, K.M. Mechanisms of and barriers to, horizontal gene transfer between bacteria. Nat. Rev. Microbiol. 2005, 3, 711–721. [Google Scholar] [CrossRef] [PubMed]
- Cabezon, E.; Ripoll-Rozada, J.; Pena, A.; de la Cruz, F.; Arechaga, I. Towards an integrated model of bacterial conjugation. FEMS Microbiol. Rev. 2015, 39, 81–95. [Google Scholar] [CrossRef] [PubMed]
- Frost, L.S.; Leplae, R.; Summers, A.O.; Toussaint, A. Mobile genetic elements: The agents of open source evolution. Nat. Rev. Microbiol. 2005, 3, 722–732. [Google Scholar] [CrossRef] [PubMed]
- Smillie, C.; Garcillan-Barcia, M.P.; Francia, M.V.; Rocha, E.P.; de la Cruz, F. Mobility of plasmids. Microbiol. Mol. Biol. Rev. 2010, 74, 434–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thoma, L.; Muth, G. The conjugative DNA-transfer apparatus of Streptomyces. Int. J. Med. Microbiol. 2015, 305, 224–229. [Google Scholar] [CrossRef] [PubMed]
- Garcillan-Barcia, M.P.; Francia, M.V.; de la Cruz, F. The diversity of conjugative relaxases and its application in plasmid classification. FEMS Microbiol. Rev. 2009, 33, 657–687. [Google Scholar] [CrossRef] [PubMed]
- Ilangovan, A.; Connery, S.; Waksman, G. Structural biology of the gram-negative bacterial conjugation systems. Trends Microbiol. 2015, 23, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Goessweiner-Mohr, N.; Arends, K.; Keller, W.; Grohmann, E. Conjugative type IV secretion systems in gram-positive bacteria. Plasmid 2013, 70, 289–302. [Google Scholar] [CrossRef] [PubMed]
- Guglielmini, J.; Neron, B.; Abby, S.S.; Garcillan-Barcia, M.P.; de la Cruz, F.; Rocha, E.P. Key components of the eight classes of type IV secretion systems involved in bacterial conjugation or protein secretion. Nucleic Acids Res. 2014, 42, 5715–5727. [Google Scholar] [CrossRef] [PubMed]
- Ramsay, J.P.; Kwong, S.M.; Murphy, R.J.; Yui Eto, K.; Price, K.J.; Nguyen, Q.T.; O’Brien, F.G.; Grubb, W.B.; Coombs, G.W.; Firth, N. An updated view of plasmid conjugation and mobilization in Staphylococcus. Mob. Genet. Elements 2016, 6, e1208317. [Google Scholar] [CrossRef] [PubMed]
- Bellanger, X.; Payot, S.; Leblond-Bourget, N.; Guedon, G. Conjugative and mobilizable genomic islands in bacteria: Evolution and diversity. FEMS Microbiol. Rev. 2014, 38, 720–760. [Google Scholar] [CrossRef] [PubMed]
- Burrus, V.; Pavlovic, G.; Decaris, B.; Guedon, G. Conjugative transposons: The tip of the iceberg. Mol. Microbiol. 2002, 46, 601–610. [Google Scholar] [CrossRef] [PubMed]
- Ambroset, C.; Coluzzi, C.; Guedon, G.; Devignes, M.D.; Loux, V.; Lacroix, T.; Payot, S.; Leblond-Bourget, N. New insights into the classification and integration specificity of Streptococcus integrative conjugative elements through extensive genome exploration. Front. Microbiol. 2016, 6, 1483. [Google Scholar] [CrossRef] [PubMed]
- Ghinet, M.G.; Bordeleau, E.; Beaudin, J.; Brzezinski, R.; Roy, S.; Burrus, V. Uncovering the prevalence and diversity of integrating conjugative elements in actinobacteria. PLoS ONE 2011, 6, e27846. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.M.; Grossman, A.D. Integrative and conjugative elements (ICEs): What they do and how they work. Annu. Rev. Genet. 2015, 49, 577–601. [Google Scholar] [CrossRef] [PubMed]
- Guglielmini, J.; de la Cruz, F.; Rocha, E.P. Evolution of conjugation and type IV secretion systems. Mol. Biol. Evol. 2013, 30, 315–331. [Google Scholar] [CrossRef] [PubMed]
- Parsons, J.A.; Bannam, T.L.; Devenish, R.J.; Rood, J.I. TcpA, an FtsK/SpoIIIE homolog, is essential for transfer of the conjugative plasmid pCW3 in Clostridium perfringens. J. Bacteriol. 2007, 189, 7782–7790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steen, J.A.; Bannam, T.L.; Teng, W.L.; Devenish, R.J.; Rood, J.I. The putative coupling protein TcpA interacts with other pCW3-encoded proteins to form an essential part of the conjugation complex. J. Bacteriol. 2009, 191, 2926–2933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burrus, V. Mechanisms of stabilization of integrative and conjugative elements. Curr. Opin. Microbiol. 2017, 38, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Carraro, N.; Burrus, V. The dualistic nature of integrative and conjugative elements. Mob. Genet. Elements 2015, 5, 98–102. [Google Scholar] [CrossRef] [PubMed]
- Toussaint, A.; Merlin, C. Mobile elements as a combination of functional modules. Plasmid 2002, 47, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Burrus, V.; Pavlovic, G.; Decaris, B.; Guédon, G. The ICESt1 element of Streptococcus thermophilus belongs to a large family of integrative and conjugative elements that exchange modules and change their specificity of integration. Plasmid 2002, 48, 77–97. [Google Scholar] [CrossRef]
- Ramsay, J.P.; Firth, N. Diverse mobilization strategies facilitate transfer of non-conjugative mobile genetic elements. Curr. Opin. Microbiol. 2017, 38, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Carraro, N.; Rivard, N.; Burrus, V.; Ceccarelli, D. Mobilizable genomic islands, different strategies for the dissemination of multidrug resistance and other adaptive traits. Mob. Genet. Elements 2017, 7, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Coluzzi, C.; Guedon, G.; Devignes, M.D.; Ambroset, C.; Loux, V.; Lacroix, T.; Payot, S.; Leblond-Bourget, N. A glimpse into the world of integrative and mobilizable elements in streptococci reveals an unexpected diversity and novel families of mobilization proteins. Front. Microbiol. 2017, 8, 443. [Google Scholar] [CrossRef] [PubMed]
- Daccord, A.; Ceccarelli, D.; Burrus, V. Integrating conjugative elements of the SXT/R391 family trigger the excision and drive the mobilization of a new class of Vibrio genomic islands. Mol. Microbiol. 2010, 78, 576–588. [Google Scholar] [CrossRef] [PubMed]
- Achard, A.; Leclercq, R. Characterization of a small mobilizable transposon, MTnSag1, in Streptococcus agalactiae. J. Bacteriol. 2007, 189, 4328–4331. [Google Scholar] [CrossRef] [PubMed]
- Lyras, D.; Adams, V.; Ballard, S.A.; Teng, W.L.; Howarth, P.M.; Crellin, P.K.; Bannam, T.L.; Songer, J.G.; Rood, J.I. tISCpe8, an IS1595-family lincomycin resistance element located on a conjugative plasmid in Clostridium perfringens. J. Bacteriol. 2009, 191, 6345–6351. [Google Scholar] [CrossRef] [PubMed]
- Giovanetti, E.; Brenciani, A.; Tiberi, E.; Bacciaglia, A.; Varaldo, P.E. ICESp2905, the erm(TR)-tet(O) element of Streptococcus pyogenes, is formed by two independent integrative and conjugative elements. Antimicrob. Agents Chemother. 2012, 56, 591–594. [Google Scholar] [CrossRef] [PubMed]
- Brenciani, A.; Tiberi, E.; Bacciaglia, A.; Petrelli, D.; Varaldo, P.E.; Giovanetti, E. Two distinct genetic elements are responsible for erm(TR)-mediated erythromycin resistance in tetracycline-susceptible and tetracycline-resistant strains of Streptococcus pyogenes. Antimicrob. Agents Chemother. 2011, 55, 2106–2112. [Google Scholar] [CrossRef] [PubMed]
- Brouwer, M.S.; Warburton, P.J.; Roberts, A.P.; Mullany, P.; Allan, E. Genetic organisation, mobility and predicted functions of genes on integrated, mobile genetic elements in sequenced strains of Clostridium difficile. PLoS ONE 2011, 6, e23014. [Google Scholar] [CrossRef] [PubMed]
- Adams, V.; Lyras, D.; Farrow, K.A.; Rood, J.I. The clostridial mobilisable transposons. Cell. Mol. Life Sci. 2002, 59, 2033–2043. [Google Scholar] [CrossRef] [PubMed]
- Billington, S.J.; Songer, J.G.; Jost, B.H. Widespread distribution of a Tet W determinant among tetracycline-resistant isolates of the animal pathogen Arcanobacterium pyogenes. Antimicrob. Agents Chemother. 2002, 46, 1281–1287. [Google Scholar] [CrossRef] [PubMed]
- Lavigne, J.P.; Vergunst, A.C.; Bourg, G.; O’Callaghan, D. The IncP island in the genome of Brucella suis 1330 was acquired by site-specific integration. Infect. Immun. 2005, 73, 7779–7783. [Google Scholar] [CrossRef] [PubMed]
- Harmer, C.J.; Hamidian, M.; Hall, R.M. pIP40a, a type 1 IncC plasmid from 1969 carries the integrative element GIsul2 and a novel class II mercury resistance transposon. Plasmid 2017, 92, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo-Diaz, F.; Fernandez-Lopez, C.; Douarre, P.E.; Baez-Ortega, A.; Flores, C.; Glaser, P.; Espinosa, M. Streptococcal group B integrative and mobilizable element IMESag-rpsI encodes a functional relaxase involved in its transfer. Open Biol. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Doublet, B.; Boyd, D.; Mulvey, M.R.; Cloeckaert, A. The Salmonella genomic island 1 is an integrative mobilizable element. Mol. Microbiol. 2005, 55, 1911–1924. [Google Scholar] [CrossRef] [PubMed]
- Carraro, N.; Rivard, N.; Ceccarelli, D.; Colwell, R.R.; Burrus, V. IncA/C conjugative plasmids mobilize a new family of multidrug resistance islands in clinical Vibrio cholerae non-O1/non-O139 isolates from Haiti. MBio 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Daccord, A.; Ceccarelli, D.; Rodrigue, S.; Burrus, V. Comparative analysis of mobilizable genomic islands. J. Bacteriol. 2013, 195, 606–614. [Google Scholar] [CrossRef] [PubMed]
- Carraro, N.; Matteau, D.; Luo, P.; Rodrigue, S.; Burrus, V. The master activator of IncA/C conjugative plasmids stimulates genomic islands and multidrug resistance dissemination. PLoS Genet. 2014, 10, e1004714. [Google Scholar] [CrossRef] [PubMed]
- Holden, M.T.; Seth-Smith, H.M.; Crossman, L.C.; Sebaihia, M.; Bentley, S.D.; Cerdeno-Tarraga, A.M.; Thomson, N.R.; Bason, N.; Quail, M.A.; Sharp, S.; et al. The genome of Burkholderia cenocepacia J2315, an epidemic pathogen of cystic fibrosis patients. J. Bacteriol. 2009, 191, 261–277. [Google Scholar] [CrossRef] [PubMed]
- Shoemaker, N.B.; Wang, G.R.; Stevens, A.M.; Salyers, A.A. Excision, transfer and integration of NBU1, a mobilizable site-selective insertion element. J. Bacteriol. 1993, 175, 6578–6587. [Google Scholar] [CrossRef] [PubMed]
- Naito, M.; Ogura, Y.; Itoh, T.; Shoji, M.; Okamoto, M.; Hayashi, T.; Nakayama, K. The complete genome sequencing of Prevotella intermedia strain OMA14 and a subsequent fine-scale, intra-species genomic comparison reveal an unusual amplification of conjugative and mobile transposons and identify a novel prevotella-lineage-specific repeat. DNA Res. 2016, 23, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Puymege, A.; Bertin, S.; Chuzeville, S.; Guedon, G.; Payot, S. Conjugative transfer and cis-mobilization of a genomic island by an integrative and conjugative element of Streptococcus agalactiae. J. Bacteriol. 2013. [CrossRef] [PubMed]
- Ghosh, S.; Sadowsky, M.J.; Roberts, M.C.; Gralnick, J.A.; LaPara, T.M. Sphingobacterium sp. Strain PM2-P1–29 harbours a functional tet(X) gene encoding for the degradation of tetracycline. J. Appl. Microbiol. 2009, 106, 1336–1342. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Shoemaker, N.B.; Wang, G.R.; Salyers, A.A. Characterization of a Bacteroides mobilizable transposon, NBU2, which carries a functional lincomycin resistance gene. J. Bacteriol. 2000, 182, 3559–3571. [Google Scholar] [CrossRef] [PubMed]
- Puymege, A.; Bertin, S.; Guedon, G.; Payot, S. Analysis of Streptococcus agalactiae pan-genome for prevalence, diversity and functionality of integrative and conjugative or mobilizable elements integrated in the tRNA(Lys CTT) gene. Mol. Genet. Genom. 2015, 290, 1727–1740. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.J.; Parker, A.C. Identification of a circular intermediate in the transfer and transposition of Tn4555, a mobilizable transposon from Bacteroides spp. J. Bacteriol. 1993, 175, 2682–2691. [Google Scholar] [CrossRef] [PubMed]
- Vedantam, G.; Novicki, T.J.; Hecht, D.W. Bacteroides fragilis transfer factor Tn5520: The smallest bacterial mobilizable transposon containing single integrase and mobilization genes that function in Escherichia coli. J. Bacteriol. 1999, 181, 2564–2571. [Google Scholar] [PubMed]
- Dingle, K.E.; Elliott, B.; Robinson, E.; Griffiths, D.; Eyre, D.W.; Stoesser, N.; Vaughan, A.; Golubchik, T.; Fawley, W.N.; Wilcox, M.H.; et al. Evolutionary history of the Clostridium difficile pathogenicity locus. Genome Biol. Evol. 2014, 6, 36–52. [Google Scholar] [CrossRef] [PubMed]
- Bass, K.A.; Hecht, D.W. Isolation and characterization of cLV25, a Bacteroides fragilis chromosomal transfer factor resembling multiple Bacteroides sp. mobilizable transposons. J. Bacteriol. 2002, 184, 1895–1904. [Google Scholar] [CrossRef] [PubMed]
- Hecht, D.W.; Malamy, M.H. Tn4399, a conjugal mobilizing transposon of Bacteroides fragilis. J. Bacteriol. 1989, 171, 3603–3608. [Google Scholar] [CrossRef] [PubMed]
- Bannam, T.L.; Crellin, P.K.; Rood, J.I. Molecular genetics of the chloramphenicol-resistance transposon Tn4451 from Clostridium perfringens: The TnpX site-specific recombinase excises a circular transposon molecule. Mol. Microbiol. 1995, 16, 535–551. [Google Scholar] [CrossRef] [PubMed]
- Lyras, D.; Adams, V.; Lucet, I.; Rood, J.I. The large resolvase TnpX is the only transposon-encoded protein required for transposition of the Tn4451/3 family of integrative mobilizable elements. Mol. Microbiol. 2004, 51, 1787–1800. [Google Scholar] [CrossRef] [PubMed]
- Siguier, P.; Gourbeyre, E.; Varani, A.; Ton-Hoang, B.; Chandler, M. Everyman’s guide to bacterial insertion sequences. Microbiol. Spectr. 2015, 3. [Google Scholar] [CrossRef] [PubMed]
- Groth, A.C.; Calos, M.P. Phage integrases: Biology and applications. J. Mol. Biol. 2004, 335, 667–678. [Google Scholar] [CrossRef] [PubMed]
- Daccord, A.; Mursell, M.; Poulin-Laprade, D.; Burrus, V. Dynamics of the SetCD-regulated integration and excision of genomic islands mobilized by integrating conjugative elements of the SXT/R391 family. J. Bacteriol. 2012, 194, 5794–5802. [Google Scholar] [CrossRef] [PubMed]
- Parker, A.C.; Smith, C.J. A multicomponent system is required for tetracycline-induced excision of Tn4555. J. Bacteriol. 2004, 186, 438–444. [Google Scholar] [CrossRef] [PubMed]
- Wood, M.M.; Rajeev, L.; Gardner, J.F. Interactions of NBU1 IntN1 and Orf2x proteins with attachment site DNA. J. Bacteriol. 2013, 195, 5516–5525. [Google Scholar] [CrossRef] [PubMed]
- Shoemaker, N.B.; Wang, G.R.; Salyers, A.A. Multiple gene products and sequences required for excision of the mobilizable integrated bacteroides element NBU1. J. Bacteriol. 2000, 182, 928–936. [Google Scholar] [CrossRef] [PubMed]
- Rutherford, K.; Van Duyne, G.D. The ins and outs of serine integrase site-specific recombination. Curr. Opin. Struct. Biol. 2014, 24, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Douarre, P.E.; Sauvage, E.; Poyart, C.; Glaser, P. Host specificity in the diversity and transfer of lsa resistance genes in group B Streptococcus. J. Antimicrob. Chemother. 2015, 70, 3205–3213. [Google Scholar] [CrossRef] [PubMed]
- Persaud, C.; Lu, Y.; Vila-Sanjurjo, A.; Campbell, J.L.; Finley, J.; O’Connor, M. Mutagenesis of the modified bases, m(5)U1939 and psi2504, in Escherichia coli 23S rRNA. Biochem. Biophys. Res. Commun. 2010, 392, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Doublet, B.; Golding, G.R.; Mulvey, M.R.; Cloeckaert, A. Secondary chromosomal attachment site and tandem integration of the mobilizable Salmonella genomic island 1. PLoS ONE 2008, 3, e2060. [Google Scholar] [CrossRef] [PubMed]
- Shoemaker, N.B.; Wang, G.R.; Salyers, A.A. NBU1, a mobilizable site-specific integrated element from Bacteroides spp., can integrate nonspecifically in Escherichia coli. J. Bacteriol. 1996, 178, 3601–3607. [Google Scholar] [CrossRef] [PubMed]
- Tribble, G.D.; Parker, A.C.; Smith, C.J. The Bacteroides mobilizable transposon Tn4555 integrates by a site-specific recombination mechanism similar to that of the gram-positive bacterial element Tn916. J. Bacteriol. 1997, 179, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Tribble, G.D.; Parker, A.C.; Smith, C.J. Transposition genes of the Bacteroides mobilizable transposon Tn4555: Role of a novel targeting gene. Mol. Microbiol. 1999, 34, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Lyras, D.; Storie, C.; Huggins, A.S.; Crellin, P.K.; Bannam, T.L.; Rood, J.I. Chloramphenicol resistance in Clostridium difficile is encoded on Tn4453 transposons that are closely related to Tn4451 from Clostridium perfringens. Antimicrob. Agents Chemother. 1998, 42, 1563–1567. [Google Scholar] [PubMed]
- Salyers, A.A.; Shoemaker, N.B.; Li, L.Y. In the driver’s seat: The bacteroides conjugative transposons and the elements they mobilize. J. Bacteriol. 1995, 177, 5727–5731. [Google Scholar] [CrossRef] [PubMed]
- Guerillot, R.; Da Cunha, V.; Sauvage, E.; Bouchier, C.; Glaser, P. Modular evolution of TnGBSs, a new family of integrative and conjugative elements associating insertion sequence transposition, plasmid replication and conjugation for their spreading. J. Bacteriol. 2013, 195, 1979–1990. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.A.; Babic, A.; Grossman, A.D. Autonomous plasmid-like replication of a conjugative transposon. Mol. Microbiol. 2010, 75, 268–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, L.D.; Grossman, A.D. Autonomous replication of the conjugative transposon Tn916. J. Bacteriol. 2016, 198, 3355–3366. [Google Scholar] [CrossRef] [PubMed]
- Carraro, N.; Poulin, D.; Burrus, V. Replication and active partition of integrative and conjugative elements (ICEs) of the SXT/R391 family: The line between ICEs and conjugative plasmids is getting thinner. PLoS Genet. 2015, 11, e1005298. [Google Scholar] [CrossRef] [PubMed]
- Burrus, V.; Bontemps, C.; Decaris, B.; Guedon, G. Characterization of a novel type II restriction-modification system, Sth368I, encoded by the integrative element ICESt1 of Streptococcus thermophilus CNRZ368. Appl. Environ. Microbiol. 2001, 67, 1522–1528. [Google Scholar] [CrossRef] [PubMed]
- Mruk, I.; Kobayashi, I. To be or not to be: Regulation of restriction-modification systems and other toxin-antitoxin systems. Nucleic Acids Res. 2014, 42, 70–86. [Google Scholar] [CrossRef] [PubMed]
- Kiss, J.; Nagy, B.; Olasz, F. Stability, entrapment and variant formation of Salmonella genomic island 1. PLoS ONE 2012, 7, e32497. [Google Scholar] [CrossRef] [PubMed]
- Huguet, K.T.; Gonnet, M.; Doublet, B.; Cloeckaert, A. A toxin antitoxin system promotes the maintenance of the IncA/C-mobilizable Salmonella genomic island 1. Sci. Rep. 2016, 6, 32285. [Google Scholar] [CrossRef] [PubMed]
- Pavlovic, G.; Burrus, V.; Gintz, B.; Decaris, B.; Guedon, G. Evolution of genomic islands by deletion and tandem accretion by site-specific recombination: ICESt1-related elements from Streptococcus thermophilus. Microbiology 2004, 150, 759–774. [Google Scholar] [CrossRef] [PubMed]
- Bellanger, X.; Morel, C.; Gonot, F.; Puymege, A.; Decaris, B.; Guedon, G. Site-specific accretion of an integrative conjugative element together with a related genomic island leads to cis mobilization and gene capture. Mol. Microbiol. 2011, 81, 912–925. [Google Scholar] [CrossRef] [PubMed]
- Carraro, N.; Libante, V.; Morel, C.; Charron-Bourgoin, F.; Leblond, P.; Guedon, G. Plasmid-like replication of a minimal streptococcal integrative and conjugative element. Microbiology 2016. [Google Scholar] [CrossRef] [PubMed]
- Lautner, M.; Schunder, E.; Herrmann, V.; Heuner, K. Regulation, integrase-dependent excision and horizontal transfer of genomic islands in Legionella pneumophila. J. Bacteriol. 2013, 195, 1583–1597. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Loria, R. Emergence of novel pathogenic Streptomyces species by site-specific accretion and cis-mobilization of pathogenicity islands. Mol. Plant Microbe Interact. 2017, 30, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Brochet, M.; Couve, E.; Glaser, P.; Guedon, G.; Payot, S. Integrative conjugative elements and related elements are major contributors to the genome diversity of Streptococcus agalactiae. J. Bacteriol. 2008, 190, 6913–6917. [Google Scholar] [CrossRef] [PubMed]
- Murphy, C.G.; Malamy, M.H. Requirements for strand- and site-specific cleavage within the oriT region of Tn4399, a mobilizing transposon from Bacteroides fragilis. J. Bacteriol. 1995, 177, 3158–3165. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.J.; Parker, A.C. The transfer origin for Bacteroides mobilizable transposon Tn4555 is related to a plasmid family from gram-positive bacteria. J. Bacteriol. 1998, 180, 435–439. [Google Scholar] [PubMed]
- Marcoleta, A.E.; Berrios-Pasten, C.; Nunez, G.; Monasterio, O.; Lagos, R. Klebsiella pneumoniae asparagine tDNAs are integration hotspots for different genomic islands encoding microcin e492 production determinants and other putative virulence factors present in hypervirulent strains. Front. Microbiol. 2016, 7, 849. [Google Scholar] [CrossRef] [PubMed]
- Murphy, C.G.; Malamy, M.H. Characterization of a “mobilization cassette” in transposon Tn4399 from Bacteroides fragilis. J. Bacteriol. 1993, 175, 5814–5823. [Google Scholar] [CrossRef] [PubMed]
- Yoshii, A.; Moriyama, H.; Fukuhara, T. The novel kasugamycin 2′-N-acetyltransferase gene aac(2′)-IIa, carried by the IncP island, confers kasugamycin resistance to rice-pathogenic bacteria. Appl. Environ. Microbiol. 2012, 78, 5555–5564. [Google Scholar] [CrossRef] [PubMed]
- Goh, S.; Hussain, H.; Chang, B.J.; Emmett, W.; Riley, T.V.; Mullany, P. Phage phiC2 mediates transduction of Tn6215, encoding erythromycin resistance, between Clostridium difficile strains. MBio 2013, 4, e00840-13. [Google Scholar] [CrossRef] [PubMed]
- Crellin, P.K.; Rood, J.I. Tn4451 from Clostridium perfringens is a mobilizable transposon that encodes the functional Mob protein, TnpZ. Mol. Microbiol. 1998, 27, 631–642. [Google Scholar] [CrossRef] [PubMed]
- Douard, G.; Praud, K.; Cloeckaert, A.; Doublet, B. The Salmonella genomic island 1 is specifically mobilized in trans by the IncA/C multidrug resistance plasmid family. PLoS ONE 2010, 5, e15302. [Google Scholar] [CrossRef] [PubMed]
- Boyd, D.; Peters, G.A.; Cloeckaert, A.; Boumedine, K.S.; Chaslus-Dancla, E.; Imberechts, H.; Mulvey, M.R. Complete nucleotide sequence of a 43-kilobase genomic island associated with the multidrug resistance region of Salmonella enterica serovar Typhimurium DT104 and its identification in phage type DT120 and serovar Agona. J. Bacteriol. 2001, 183, 5725–5732. [Google Scholar] [CrossRef] [PubMed]
- Siebor, E.; de Curraize, C.; Amoureux, L.; Neuwirth, C. Mobilization of the Salmonella genomic island SGI1 and the Proteus genomic island PGI1 by the A/C2 plasmid carrying blaTEM-24 harboured by various clinical species of Enterobacteriaceae. J. Antimicrob. Chemother. 2016, 71, 2167–2170. [Google Scholar] [CrossRef] [PubMed]
- Haase, J.; Kalkum, M.; Lanka, E. TrbK, a small cytoplasmic membrane lipoprotein, functions in entry exclusion of the IncP alpha plasmid RP4. J. Bacteriol. 1996, 178, 6720–6729. [Google Scholar] [CrossRef] [PubMed]
- Haase, J.; Lurz, R.; Grahn, A.M.; Bamford, D.H.; Lanka, E. Bacterial conjugation mediated by plasmid RP4: RSF1010 mobilization, donor-specific phage propagation and pilus production require the same Tra2 core components of a proposed DNA transport complex. J. Bacteriol. 1995, 177, 4779–4791. [Google Scholar] [CrossRef] [PubMed]
- Wisniewski, J.A.; Traore, D.A.; Bannam, T.L.; Lyras, D.; Whisstock, J.C.; Rood, J.I. TcpM: A novel relaxase that mediates transfer of large conjugative plasmids from Clostridium perfringens. Mol. Microbiol. 2016, 99, 884–896. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.A.; Thomas, J.; Grossman, A.D. The Bacillus subtilis conjugative transposon ICEBs1 mobilizes plasmids lacking dedicated mobilization functions. J. Bacteriol. 2012, 194, 3165–3172. [Google Scholar] [CrossRef] [PubMed]
- Naglich, J.G.; Andrews, R.E., Jr. Tn916-dependent conjugal transfer of pC194 and pUB110 from Bacillus subtilis into Bacillus thuringiensis subsp. israelensis. Plasmid 1988, 20, 113–126. [Google Scholar] [CrossRef]
- Showsh, S.A.; Andrews, R.E., Jr. Analysis of the requirement for a pUB110 mob region during Tn916-dependent mobilization. Plasmid 1999, 41, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Mingoia, M.; Morici, E.; Marini, E.; Brenciani, A.; Giovanetti, E.; Varaldo, P.E. Macrolide resistance gene erm(TR) and erm(TR)-carrying genetic elements in Streptococcus agalactiae: Characterization of ICESagTR7, a new composite element containing IMESp2907. J. Antimicrob. Chemother. 2016, 71, 593–600. [Google Scholar] [CrossRef] [PubMed]
- Morici, E.; Simoni, S.; Brenciani, A.; Giovanetti, E.; Varaldo, P.E.; Mingoia, M. A new mosaic integrative and conjugative element from Streptococcus agalactiae carrying resistance genes for chloramphenicol (catQ) and macrolides [mef(I) and erm(TR)]. J. Antimicrob. Chemother. 2016, 72, 64–67. [Google Scholar] [CrossRef] [PubMed]
- Harmer, C.J.; Hamidian, M.; Ambrose, S.J.; Hall, R.M. Destabilization of IncA and IncC plasmids by SGI1 and SGI2 type Salmonella genomic islands. Plasmid 2016, 87–88, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Carraro, N.; Durand, R.; Rivard, N.; Anquetil, C.; Barrette, C.; Humbert, M.; Burrus, V. Salmonella genomic island 1 (SGI1) reshapes the mating apparatus of IncC conjugative plasmids to promote self-propagation. PLoS Genet. 2017, 13, e1006705. [Google Scholar] [CrossRef] [PubMed]
- Billington, S.J.; Jost, B.H. Multiple genetic elements carry the tetracycline resistance gene tet(W) in the animal pathogen Arcanobacterium pyogenes. Antimicrob. Agents Chemother. 2006, 50, 3580–3587. [Google Scholar] [CrossRef] [PubMed]
- Whittle, G.; Hund, B.D.; Shoemaker, N.B.; Salyers, A.A. Characterization of the 13-kilobase ermF region of the Bacteroides conjugative transposon cTnDOT. Appl. Environ. Microbiol. 2001, 67, 3488–3495. [Google Scholar] [CrossRef] [PubMed]
- Siebor, E.; Neuwirth, C. The new variant of Salmonella genomic island 1 (SGI1-V) from a Proteus mirabilis French clinical isolate harbours blaVEB-6 and qnrA1 in the multiple antibiotic resistance region. J. Antimicrob. Chemother. 2011, 66, 2513–2520. [Google Scholar] [CrossRef] [PubMed]
- Schultz, E.; Barraud, O.; Madec, J.Y.; Haenni, M.; Cloeckaert, A.; Ploy, M.C.; Doublet, B. Multidrug resistance Salmonella genomic island 1 in a Morganella morganii subsp. Morganii human clinical isolate from france. mSphere 2017, 2. [Google Scholar] [CrossRef]
- Ferreira, L.Q.; Avelar, K.E.; Vieira, J.M.; de Paula, G.R.; Colombo, A.P.; Domingues, R.M.; Ferreira, M.C. Association between the cfxA gene and transposon Tn4555 in Bacteroides distasonis strains and other bacteroides species. Curr. Microbiol. 2007, 54, 348–353. [Google Scholar] [CrossRef] [PubMed]
- Soliman, A.M.; Ahmed, A.M.; Shimamoto, T.; El-Domany, R.A.; Nariya, H. First report in Africa of two clinical isolates of Proteus mirabilis carrying Salmonella genomic island (SGI1) variants, SGI1-PmABB and SGI1-W. Infect. Genet. Evol. 2017, 51, 132–137. [Google Scholar] [CrossRef] [PubMed]
- Schultz, E.; Haenni, M.; Mereghetti, L.; Siebor, E.; Neuwirth, C.; Madec, J.Y.; Cloeckaert, A.; Doublet, B. Survey of multidrug resistance integrative mobilizable elements SGI1 and PGI1 in Proteus mirabilis in humans and dogs in france, 2010–2013. J. Antimicrob. Chemother. 2015, 70, 2543–2546. [Google Scholar] [CrossRef] [PubMed]
- Yoshii, A.; Omatsu, T.; Katayama, Y.; Koyama, S.; Mizutani, T.; Moriyama, H.; Fukuhara, T. Two types of genetic carrier, the IncP genomic island and the novel IncP-1beta plasmid, for the aac(2′)-IIa gene that confers kasugamycin resistance in Acidovorax avenae ssp. avenae. Mol. Plant Pathol. 2015, 16, 288–300. [Google Scholar] [CrossRef] [PubMed]
- Shoemaker, N.B.; Barber, R.D.; Salyers, A.A. Cloning and characterization of a Bacteroides conjugal tetracycline-erythromycin resistance element by using a shuttle cosmid vector. J. Bacteriol. 1989, 171, 1294–1302. [Google Scholar] [CrossRef] [PubMed]
- Magot, M. Physical characterization of the Clostridium perfringens tetracycline-chloramphenicol resistance plasmid pIP401. Ann. Microbiol. (Paris) 1984, 135, 269–282. [Google Scholar] [CrossRef]
- Balado, M.; Lemos, M.L.; Osorio, C.R. Integrating conjugative elements of the SXT/R391 family from fish-isolated Vibrios encode restriction-modification systems that confer resistance to bacteriophages. FEMS Microbiol. Ecol. 2013, 83, 457–467. [Google Scholar] [CrossRef] [PubMed]
- Ershova, A.S.; Rusinov, I.S.; Spirin, S.A.; Karyagina, A.S.; Alexeevski, A.V. Role of restriction-modification systems in prokaryotic evolution and ecology. Biochemistry (Mosc.) 2015, 80, 1373–1386. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Dietrich, C.; Hongoh, Y.; Brune, A. Restriction-modification systems as mobile genetic elements in the evolution of an intracellular symbiont. Mol. Biol. Evol. 2016, 33, 721–725. [Google Scholar] [CrossRef] [PubMed]
- Gerdes, K.; Christensen, S.K.; Lobner-Olesen, A. Prokaryotic toxin-antitoxin stress response loci. Nat. Rev. Microbiol. 2005, 3, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.Y.; Awano, N.; Masuda, H.; Park, J.H.; Inouye, M. Transcriptional repressor HipB regulates the multiple promoters in Escherichia coli. J. Mol. Microbiol. Biotechnol. 2013, 23, 440–447. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, M.A.; Balani, P.; Min, J.; Chinnam, N.B.; Hansen, S.; Vulic, M.; Lewis, K.; Brennan, R.G. HipBA-promoter structures reveal the basis of heritable multidrug tolerance. Nature 2015, 524, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, M.A.; Piro, K.M.; Xu, W.; Hansen, S.; Lewis, K.; Brennan, R.G. Molecular mechanisms of HipA-mediated multidrug tolerance and its neutralization by HipB. Science 2009, 323, 396–401. [Google Scholar] [CrossRef] [PubMed]
- Cury, J.; Touchon, M.; Rocha, E.P.C. Integrative and conjugative elements and their hosts: Composition, distribution and organization. Nucleic Acids Res. 2017, 45, 8943–8956. [Google Scholar] [CrossRef] [PubMed]
- Trudel, M.V.; Vincent, A.T.; Attere, S.A.; Labbe, M.; Derome, N.; Culley, A.I.; Charette, S.J. Diversity of antibiotic-resistance genes in Canadian isolates of Aeromonas salmonicida subsp. salmonicida: Dominance of pSN254b and discovery of pAsa8. Sci. Rep. 2016, 6, 35617. [Google Scholar] [CrossRef] [PubMed]
- Adams, V.; Johanesen, P.A.; Rood, J.I.; Lyras, D. Mobilisable genetic elements from the Clostridia. In Bacterial Integrative Mobile Genetic Elements; Roberts, A., Mullany, P., Eds.; Landes Bioscience: Austin, TX, USA, 2013; pp. 120–134. [Google Scholar]
- Mulvey, M.R.; Boyd, D.A.; Olson, A.B.; Doublet, B.; Cloeckaert, A. The genetics of Salmonella genomic island 1. Microbes Infect. 2006, 8, 1915–1922. [Google Scholar] [CrossRef] [PubMed]
- Bardaji, L.; Echeverria, M.; Rodriguez-Palenzuela, P.; Martinez-Garcia, P.M.; Murillo, J. Four genes essential for recombination define GInts, a new type of mobile genomic island widespread in bacteria. Sci. Rep. 2017, 7, 46254. [Google Scholar] [CrossRef] [PubMed]
- Boyd, E.F.; Almagro-Moreno, S.; Parent, M.A. Genomic islands are dynamic, ancient integrative elements in bacterial evolution. Trends Microbiol. 2009, 17, 47–53. [Google Scholar] [CrossRef] [PubMed]
- McMurdie, P.J.; Hug, L.A.; Edwards, E.A.; Holmes, S.; Spormann, A.M. Site-specific mobilization of vinyl chloride respiration islands by a mechanism common in Dehalococcoides. BMC Genom. 2011, 12, 287. [Google Scholar] [CrossRef] [PubMed]
- Christie, G.E.; Dokland, T. Pirates of the Caudovirales. Virology 2012, 434, 210–221. [Google Scholar] [CrossRef] [PubMed]
- Siguier, P.; Gagnevin, L.; Chandler, M. The new IS1595 family, its relation to IS1 and the frontier between insertion sequences and transposons. Res. Microbiol. 2009, 160, 232–241. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IME a | Species (Division) b | Int c | Integration Site d | Putative Maintenance Genes e | Reference |
|---|---|---|---|---|---|
| MTnSag1 | Streptococcus agalactiae (fir.) | DDE | AT-rich regions | None | [28] |
| tISCpe8 | Clostridium perfringens (fir.) | DDE | AT-rich regions | None | [29] |
| IME_SpnAP200_rumA | Streptococcus pneumoniae (fir.) | Ser | Internal site of rumA (23S rRNA methyltransferase) | Replisome organizer | [26] |
| IME_ScoC232_maff2_site1 f | Streptococcus constellatus (fir.) | Ser | Internal site S1 of a gene (Maff2-related) from Tn5252-related ICEs | Replisome organizer, DnaC | [26] |
| IMESp2907 f | Streptococcus pyogenes (fir.) | Ser | Internal site S2 of a gene (Maff2-related) from Tn5252-related ICEs | Replisome organizer | [30] |
| tet(O) fragment | Streptococcus pyogenes (fir.) | Ser | Internal site of a gene (SNF2 helicase) from Tn5252-related ICEs | RepA, ParB | [31] |
| Tn6104 | Clostridioides difficile (fir.) | Ser | Internal site of traG (VirD4 CP) from Tn5252 related ICEs | Replisome organizer, DnaC, TA | [32] |
| IME_Sco1050_traG_site1 f | Streptococcus constellatus C1050 (fir.) | Ser | Internal site S1 of traG (VirD4 CP) from Tn5252-related ICEs | Replisome organizer | [26] |
| IME_Sco1050_traG_site2 f | Streptococcus constellatus C1050 (fir.) | Ser | Internal site S2 of traG (VirD4 CP) from Tn5252-related ICEs | Replisome organizer | [26] |
| Tn4451 f | Clostridium perfringens (fir.) | Ser | Numerous sites (GA) | [33] | |
| ATE-1 | Trueperella pyogenes (act.) | Tyr | 3′ end of guaA (GMP synthase) | TA | [34] |
| IncP island g | Brucella suis (α) | Tyr | 3′ end of guaA (GMP synthase) | RepA, antitoxin | [35] |
| Gisul2 | Pseudomonas aeruginosa (γ) h | Tyr | 3′ end of guaA (GMP synthase) | RepA, RepC | [36] |
| IME_SagNEM316_rplL f | Streptococcus agalactiae (fir.) | Tyr | 3′ end of rplL (L7/L12 ribosomal protein) | [26] | |
| IME_Sga2069_rpmE f | Streptococcus gallolyticus (fir.) | Tyr | 3′ end of rpmE (L31 ribosomal protein) | Rep_Trans | [26] |
| IME_SSalJIM777_rpmG f | Streptococcus salivarius (fir.) | Tyr | 3′ end of rpmG (L33 ribosomal protein) | [26] | |
| IMESag-rpsI g | Streptococcus agalactiae HRC (fir.) | Tyr | 3′ end of rpsI (S9 ribosomal protein) | Rep_Trans, TA | [37] |
| SGI1 g | Salmonella enterica DT104 (γ) | Tyr | 3′ end of trmE (tRNA modification GTPase) | Rep_3, TA | [38] |
| MGIVchHai6 | Vibrio cholerae (γ) | Tyr | 3′ end of trmE (tRNA modification GTPase) | [39] | |
| MGIVchMoz6 | Vibrio cholerae (γ) | Tyr | 3′ end of yicC (unknown) | RM II | [40] |
| MGIVchUSA1 f | Vibrio cholerae (γ) | Tyr | 3′ end of yicC (unknown) | 2 TAs | [27] |
| MGIVflInd1 f | Vibrio fluvialis (γ) | Tyr | 3′ end of yicC (unknown) | [27] | |
| MGIVmi1 | Vibrio mimicus (γ) | Tyr | 3′ end of yicC (unknown) | [41] | |
| BcenGI2 f | Burkholderia cenocepacia (β) | Tyr | 3′ end of tRNAala gene | Rep_3, TA | [42] |
| IME_SsuTL13-tRNAasn | Streptococcus suis (fir.) | Tyr | 3′ end of tRNAarg gene | [26] | |
| IME_SanC1051_tRNAarg | Streptococcus anginosus (fir.) | Tyr | 3′ end of tRNAasn gene | [26] | |
| NBU1 | Bacteroides uniformis (bac.) | Tyr | 3′ end of tRNAleu gene | TA | [43] |
| MTnPi4 | Prevotella intermedia (bac.) | Tyr | 3′ end of tRNAleu gene | [44] | |
| IME_Sag2603_tRNAlys f | Streptococcus agalactiae (fir.) | Tyr | 3′ end of tRNAlys gene | [45] | |
| Tn6031 | Sphingobacterium sp. (bac.) | Tyr | 3′ end of tRNApro gene | [46] | |
| NBU2 | Bacteroides fragilis (bac.) | Tyr | 3′ end of tRNAser gene | [47] | |
| IME_SdyRE378_ebfC f | Streptococcus dysgalactiae (fir.) | Tyr | 5′ end of ebfC (nucleoid associated protein) | [26] | |
| IME_SanC238_tatD | Streptococcus anginosus (fir.) | Tyr | 5′ end of tatD (DNAse) | [26] | |
| IME_oriT g | Streptococcus agalactiae (fir.) | Tyr | oriT from Tn916 and ICESt3-related ICEs | [48] | |
| Tn4555 | Bacteroides vulgatus (bac.) | Tyr | Two preferred sites | [49] | |
| Tn5520 | Bacteroides fragilis (bac.) | Tyr | AT-rich regions | None | [50] |
| Tn6218 g | Clostridioides difficile (fir.) | Tyr | AT-rich regions | [51] | |
| cLV25 | Bacteroides fragilis (bac.) | Tyr | ND | [52] | |
| MTnPi1 | Prevotella intermedia (bac.) | Tyr duo | TTAC NNNNN AA | [44] | |
| MTnPi2 | Prevotella intermedia (bac.) | Tyr duo | TTGC NNNNN AA | [44] | |
| MTnPi3 | Prevotella intermedia (bac.) | Tyr duo | TTAC NNNNN A/G A/G | [44] | |
| Tn4399 | Bacteroides fragilis (bac.) | ND | Numerous sites | [53] |
| IME a | Species (Division) b | Mobilization Proteins Encoded by the IME c | Mobilizing Element (CP, MPF) d | Reference | |
|---|---|---|---|---|---|
| Relaxase | Others | ||||
| MTnSag1 | Streptococcus agalactiae (fir.) | None | None | Tn916 (TcpA, FA) | [28] |
| tISCpe8 | Clostridium perfringens (fir.) | None | None | Tn916 (TcpA, FA) | [29] |
| GIE492 e | Klebsiella pneumoniae (γ) | None | Proposed: ICEKp1 (VirD4, T) | [87] | |
| MGIVflInd1 e | Vibrio fluvialis (γ) | None | ICEVflInd1 and SXT (VirD4, F) | [27] | |
| MGIVchHai6 e | Vibrio cholerae (γ) | None | 1 RAF | IncA/C plasmids (VirD4, F) | [39] |
| MGIVmi1 e | Vibrio mimicus (γ) | None | 1 RAF | IncA/C plasmids (VirD4, F) | [41] |
| SGI1 e,f | Salmonella enterica (γ) | None | TraG, TraH, TraN | IncA/C plasmids (VirD4, F) | [38] |
| Gisul2 g | Pseudomonas aeruginosa (γ) | None | TrbJ, TrbK, TrbL | Proposed: IncP plasmids (VirD4, T) | [36] |
| IME_SsalCCHSS3_ND | Streptococcus salivarius (fir.) | MobC | VirD4 | [26] | |
| NBU1 | Bacteroides uniformis (bac.) | MobP | CTnERL and CTnDOT (VirD4, B); IncP plasmids (VirD4, T) | [43] | |
| NBU2 | Bacteroides fragilis (bac.) | MobP | CTnERL (VirD4, B); IncP plasmids (VirD4, T) | [47] | |
| Tn4555 | Bacteroides vulgatus (bac.) | MobP | CTn341 (VirD4, B); IncP plasmids (VirD4, T) | [49] | |
| cLV25 | Bacteroides fragilis (bac.) | MobP | 1 RAF | IncP plasmids (VirD4, T) | [52] |
| Tn4399 | Bacteroides fragilis (bac.) | MobP | 1 RAF | CTnDOT (VirD4, B); IncP plasmids (VirD4, T) | [88] |
| IncP island f | Burkholderia glumae (β) | MobP | 2 RAFs, TrbJ, TrbK, TrbL | Proposed: IncP plasmids (VirD4, T) | [89] |
| IME_ScoC232_maff2_site 1 | Streptococcus constellatus (fir.) | MobP | [26] | ||
| IMESp2907 e | Streptococcus pyogenes (fir.) | MobQ | Proposed: Tn5252 superfamily (VirD4, FATA) | [30] | |
| ATE-1 | Trueperella pyogenes (act.) | MobV | IncP plasmids (VirD4, T) | [34] | |
| Tn5520 | Bacteroides fragilis (bac.) | MobV | None | IncP plasmids (VirD4, T) | [50] |
| Tn6215 | Clostridioides difficile (fir.) | MobV | ND | [90] | |
| Tn4451 e | Clostridium perfringens (fir.) | MobV | IncP plasmids (VirD4, T) | [91] | |
| IMESag-rpsI f,g | Streptococcus agalactiae HRC (fir.) | MobV | pAMβ1 plasmid (VirD4, FATA) | [37] | |
| tet(O) fragment | Streptococcus pyogenes (fir.) | MobV | Proposed: Tn5252 superfamily (VirD4, FATA) | [31] | |
| IME-oriT e,f,h | Streptococcus agalactiae (fir.) | MobT | Proposed: Tn916 and ICESt3 (TcpA, FA) | [63] | |
| IME_Sag2603_tRNAlys e | Streptococcus agalactiae (fir.) | MobT | Proposed: helpers with TcpA and FA | [26] | |
| IME_SsuBM407_tRNAleu e | Streptococcus suis (fir.) | MobT | TcpA | Proposed: helpers with TcpA and FA | [26] |
| IME_SdyRE378_ebfC e | Streptococcus dysgalactiae (fir.) | PF01719 | Proposed: helpers with TcpA and FA | [26] | |
| IME_SsalJIM8777_rpmG e | Streptococcus salivarius (fir.) | PF01719 | TcpA | Proposed: helpers with TcpA and FA | [26] |
| IME_SsuTL13_rpsI e | Streptococcus suis (fir.) | PF01719-helicase | Proposed: helpers with TcpA and FA | [26] | |
| IME_Seq35246_rpsI e | Streptococcus equi (fir.) | PF01719-helicase | TcpA | Proposed: helpers with TcpA and FA | [26] |
| IME_SanC238_tatD | Streptococcus anginosus (fir.) | PHA00330 | Proposed: helpers with TcpA and FA | [26] | |
| IME_SpnA45_tRNAleu e | Streptococcus pneumoniae (fir.) | PHA00330 | TcpA | Proposed: helpers with TcpA and FA | [26] |
| IME_SiniSF1_ebfC | Streptococcus iniae (fir.) | PF02407 | TcpA | Proposed: helpers with TcpA and FA | [26] |
| IME a | Species (Division) b | Size | Putative Cargo Genes | Reference | |
|---|---|---|---|---|---|
| Resistance Genes | Others | ||||
| ATE-1 | Trueperella pyogenes (act.) | 10.8 | tet(W) (tetracycline) | TA, 3 unknown | [105] |
| Tn6031 | Sphingobacterium sp. (bac.) | 13.0 | tet(X) (tetracycline), aadS (streptomycin) | 5 unknown | [46] |
| tet(O) fragment | Streptococcus pyogenes (fir.) | 13.4 | tet(O) (tetracycline) | RNA polymerase sigma factor sigma-70, 5 unknown | [31] |
| IMESp2907 | Streptococcus pyogenes (fir.) Streptococcus agalactiae (fir.) | 12.6 | erm(TR) (macrolide, lincosamide, streptogramin) | 11 unknown | [31] [101] |
| ermF region | Bacteroides thetaiotaomicron (bac.) | 13.0 | ermF (clindamycin, erythromycin) | 4 unknown | [106] |
| tISCpe8 | Clostridium perfringens (fir.) | 2.0 | lnu(P) (lincomycin) | [29] | |
| MTnSag1 (tISSag10) | Streptococcus agalactiae (fir.) | 1.7 | lnu(C) (lincomycin) | [28] | |
| IME_GB00957_oriT | Streptococcus agalactiae (fir.) | 5.2 | lsa(C) (lincosamide, streptogramin A and pleuromutilin) | 2 unknown | [48] |
| NBU2 | Bacteroides fragilis (bac.) | 11.1 | linAN2 (lincomycin, clindamycin), mefEN2 (erythromycin) | 2 unknown | [47] |
| SGI1 | Salmonella enterica (γ) | 42.4 | aadA2 (streptomycin, spectinomycin), floR (chloramphenicol, florfenicol), tet(G) (tetracycline), blaPSE-1 (ampicillin), sul1 (sulfonamides) | TA, 18 unknown | [38] |
| SGI1-V | Proteus mirabilis (γ) | 42.9 | aacA4 (kanamycin, tobramycin, netilmicin, amikacin), aadB (kanamycin, gentamicin, tobramycin); dhfrA1 (trimethoprim), blaVEB-6 (extended-spectrum cephalosporin), sul1 (sulfonamides), qnrA1 (quinolones) | 20 unknown | [107] |
| SGI1-L | Morganella morganii (γ) | 50.3 | tet(G) (tetracycline), floR (chloramphenicol, florfenicol), dhfrA15 (trimethoprim), blaPSE-1 (amoxicillin, clavulanate), sul1 (sulfonamides) | 11 unknown | [108] |
| MGIVchHai6 | Vibrio cholerae (γ) | 47.4 | aadA2 (streptomycin, spectinomycin), floR (chloramphenicol, florfenicol), tet(G) (tetracycline), blaPSE-1 (ampicillin), sul1 (sulfonamides), merEDAFPT (mercury) | RM I, 6 unknown | [39] |
| IncP island | Brucella suis (α) | 12.7 | antitoxin, 2 unknown | [35] | |
| IncP island c | Burkholderia glumae (β), Acidovorax avenae (β) | 14.1 | aac(2′)-IIa (kasugamycin) | antitoxin, 3 unknown | [89] |
| Tn4453 c | Clostridioides difficile | 6.3 | catD (chloramphenicol) | 2 unknown | [69] |
| Tn4555 | Bacteroides vulgatus | 12.2 | cfxA (cefoxitin) | 2 unknown | [67] |
| GIsul2 | Pseudomonas aeruginosa (γ) | 15.4 | sul2 (sulphonamide), arsBCHR (arsenate/arsenite) | TA, 4 unknown | [36] |
| IME_Sag2603_tRNAlys | Clostridioides difficile (fir.) | 10.5 | ABC transporter of the drug resistance transporter subfamily, 8 unknown | [48,84] | |
| IME_SagA909_tRNAlys | Clostridium perfringens (fir.) | 8.3 | Intracellular protease, 7 unknown | [48,84] | |
| IMESag-rpsI | Streptococcus agalactiae (fir.) | 9.1 | TA, 6 unknown | [37] | |
| IME_Sag2603_rpsI | Streptococcus agalactiae (fir.) | 9.0 | arsR (arsenate reductase) | TA, 5 unknown | [84] |
| IME_18RS21_oriT | Streptococcus agalactiae (fir.) | 6.4 | merA, merR (mercury) | 3 unknown | [48] |
| MTnPi2 | Prevotella intermedia (bac.) | 16.6 | LuxR family transcriptional regulator, 9 unknown | [44] | |
| MTnPi3 | Prevotella intermedia (bac.) | 18.4 | 2 ABC transporter components, 6 unknown | [44] | |
| MTnPi4 | Prevotella intermedia (bac.) | 12.4 | Subtilase-like protease, 7 unknown | [44] | |
| MGIVchUSA1 c | Vibrio cholerae (γ) | 22.0 | two TA, 10 unknown | [27] | |
| MGIVchMoz6 | Vibrio cholerae (γ) | 19.7 | RM II, 7 unknown | [40] | |
| MGIVmi1 | Vibrio mimicus (γ) | 16.5 | RM III, 10 unknown | [87] | |
| GIE492 | Klebsiella pneumoniae (γ) | 22.3 | MccE492 microcin (bacteriocin), 7 unknown | [32] | |
| Tn6104 | Clostridioides difficile (fir.) | 15.6 | Lantibiotic synthesis (bacteriocin), T/A, 7 unknown | [61] | |
| Tn5520 | Bacteroides fragilis (bac.) | 4.7 | [87] | ||
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guédon, G.; Libante, V.; Coluzzi, C.; Payot, S.; Leblond-Bourget, N. The Obscure World of Integrative and Mobilizable Elements, Highly Widespread Elements that Pirate Bacterial Conjugative Systems. Genes 2017, 8, 337. https://doi.org/10.3390/genes8110337
Guédon G, Libante V, Coluzzi C, Payot S, Leblond-Bourget N. The Obscure World of Integrative and Mobilizable Elements, Highly Widespread Elements that Pirate Bacterial Conjugative Systems. Genes. 2017; 8(11):337. https://doi.org/10.3390/genes8110337
Chicago/Turabian StyleGuédon, Gérard, Virginie Libante, Charles Coluzzi, Sophie Payot, and Nathalie Leblond-Bourget. 2017. "The Obscure World of Integrative and Mobilizable Elements, Highly Widespread Elements that Pirate Bacterial Conjugative Systems" Genes 8, no. 11: 337. https://doi.org/10.3390/genes8110337
APA StyleGuédon, G., Libante, V., Coluzzi, C., Payot, S., & Leblond-Bourget, N. (2017). The Obscure World of Integrative and Mobilizable Elements, Highly Widespread Elements that Pirate Bacterial Conjugative Systems. Genes, 8(11), 337. https://doi.org/10.3390/genes8110337