
DNA Methylation Dynamics and Cocaine in the Brain: Progress and Prospects

Abstract
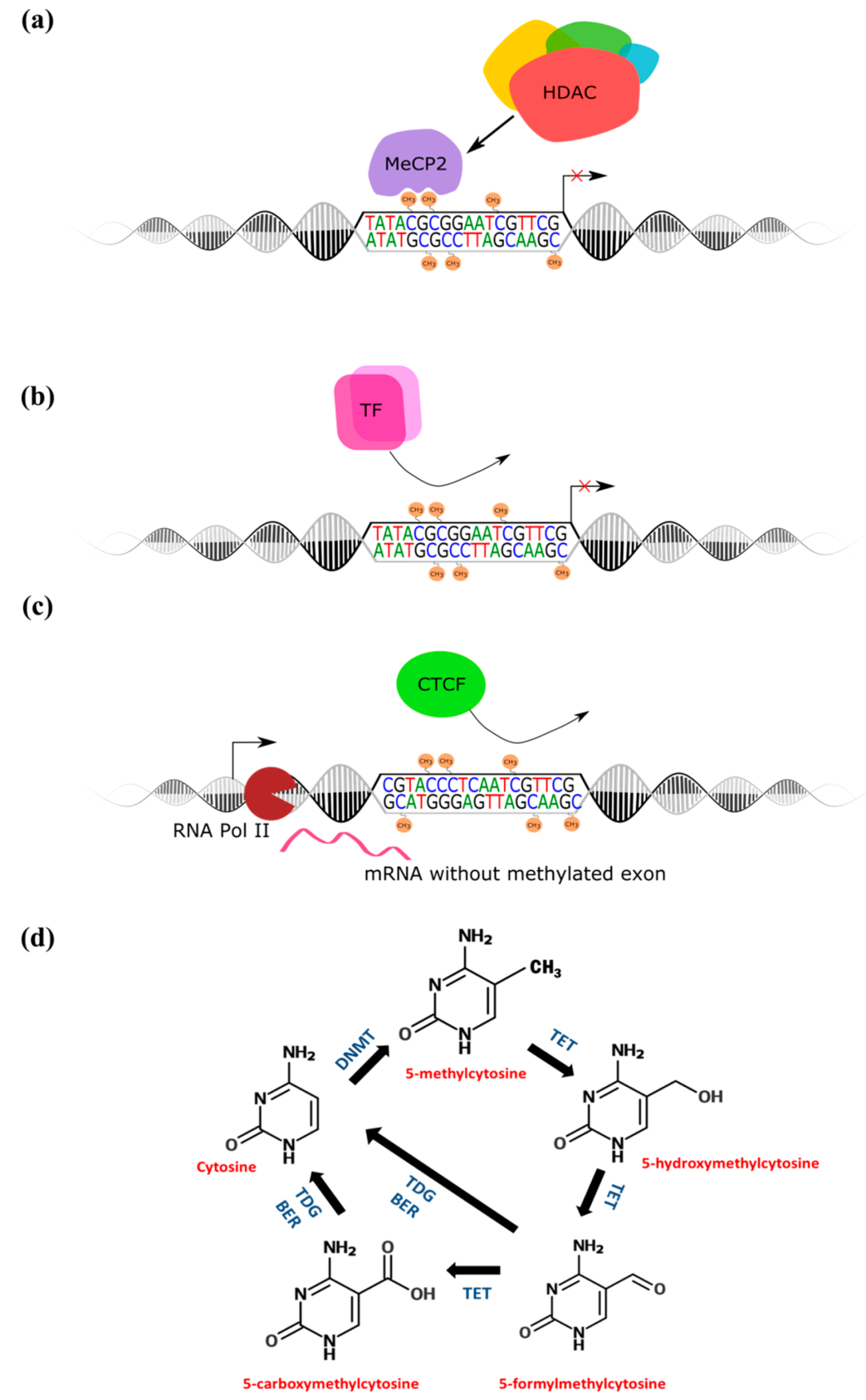
:1. Introduction
2. Cocaine-Associated Alteration of DNA Methylation Machinery
2.1. Pharmacological Manipulations: Impact on Drug-Related Behaviors
2.2. DNA Methyltransferases: The Writers of DNA Methylation
2.3. Methyl-Binding Proteins: The Readers of DNA Methylation
2.4. Methylcytosine Dioxygenases—The Modifiers of DNA Methylation
3. Cocaine-Associated Dysregulation of Methylation Dynamics
3.1. Global and Methylome-Wide Observations
3.2. Differentially Methylated Loci
3.3. Developmental Findings
4. Future Directions
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders; American Psychiatric Association: Arlington, VA, USA, 2013. [Google Scholar]
- Heal, D.J.; Gosden, J.; Smith, S.L. Dopamine reuptake transporter (DAT) “inverse agonism”—A novel hypothesis to explain the enigmatic pharmacology of cocaine. Neuropharmacology 2014, 87, 19–40. [Google Scholar] [CrossRef] [PubMed]
- Koob, G.F.; Volkow, N.D. Neurocircuitry of Addiction. Neuropsychopharmacology 2010, 35, 217–238. [Google Scholar] [CrossRef] [PubMed]
- Dumitriu, D.; LaPlant, Q.; Grossman, Y.S.; Dias, C.; Janssen, W.G.; Russo, S.J.; Morrison, J.H.; Nestler, E.J. Subregional, Dendritic Compartment, and Spine Subtype Specificity in Cocaine Regulation of Dendritic Spines in the Nucleus Accumbens. J. Neurosci. 2012, 32, 6957–6966. [Google Scholar] [CrossRef] [PubMed]
- Robinson, T.E.; Kolb, B. Alterations in the morphology of dendrites and dendritic spines in the nucleus accumbens and prefrontal cortex following repeated treatment with amphetamine or cocaine. Eur. J. Neurosci. 1999, 11, 1598–1604. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Wilkinson, M.; Liu, X.; Purushothaman, I.; Ferguson, D.; Vialou, V.; Maze, I.; Shao, N.; Kennedy, P.; Koo, J.; et al. Chronic cocaine-regulated epigenomic changes in mouse nucleus accumbens. Genome Biol. 2014, 15, R65. [Google Scholar] [CrossRef] [PubMed]
- Cowan, C.W.; Taniguchi, M.; Hale, C.F. Regulation of synaptic connectivity with chronic cocaine. Am. J. Psychiatry 2008, 165, 1393. [Google Scholar] [CrossRef] [PubMed]
- Otaka, M.; Ishikawa, M.; Lee, B.R.; Liu, L.; Neumann, P.A.; Cui, R.; Huang, Y.H.; Schluter, O.M.; Dong, Y. Exposure to Cocaine Regulates Inhibitory Synaptic Transmission in the Nucleus Accumbens. J. Neurosci. 2013, 33, 6753–6758. [Google Scholar] [CrossRef] [PubMed]
- Freeman, W.M.; Lull, M.E.; Patel, K.M.; Brucklacher, R.M.; Morgan, D.; Roberts, D.C.S.; Vrana, K.E. Gene expression changes in the medial prefrontal cortex and nucleus accumbens following abstinence from cocaine self-administration. BMC Neurosci. 2010, 11, 29. [Google Scholar] [CrossRef] [PubMed]
- Albertson, D.N.; Pruetz, B.; Schmidt, C.J.; Kuhn, D.M.; Kapatos, G.; Bannon, M.J. Gene expression profile of the nucleus accumbens of human cocaine abusers: Evidence for dysregulation of myelin. J. Neurochem. 2004, 88, 1211–1219. [Google Scholar] [CrossRef] [PubMed]
- Bannon, M.J.; Savonen, C.L.; Jia, H.; Dachet, F.; Halter, S.D.; Schmidt, C.J.; Lipovich, L.; Kapatos, G. Identification of long noncoding RNAs dysregulated in the midbrain of human cocaine abusers. J. Neurochem. 2015, 135, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Mash, D.C.; Ffrench-Mullen, J.; Adi, N.; Qin, Y.; Buck, A.; Pablo, J. Gene Expression in Human Hippocampus from Cocaine Abusers Identifies Genes which Regulate Extracellular Matrix Remodeling. PLoS ONE 2007, 2, e1187. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Yuan, Q.; Mash, D.C.; Goldman, D. Substance-specific and shared transcription and epigenetic changes in the human hippocampus chronically exposed to cocaine and alcohol. Proc. Natl. Acad. Sci. USA 2011, 108, 6626–6631. [Google Scholar] [CrossRef] [PubMed]
- Nestler, E.J. Epigenetic mechanisms of drug addiction. Neuropharmacology 2014, 76, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Bird, A.P.; Wolffe, A.P. Methylation-Induced Repression—Belts, Braces, and Chromatin. Cell 1999, 99, 451–454. [Google Scholar] [CrossRef]
- Kundu, K.; Rao, M.R. CpG Islands in Chromatin Organization and Gene Expression. J. Biochem. 1999, 125, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Domcke, S.; Bardet, A.F.; Adrian Ginno, P.; Hartl, D.; Burger, L.; Schübeler, D. Competition between DNA methylation and transcription factors determines binding of NRF1. Nature 2015, 528, 575–579. [Google Scholar] [CrossRef] [PubMed]
- Hope, B.T. Cocaine and the AP-1 transcription factor complex. Ann. N. Y. Acad. Sci. 1998, 844, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Lister, R.; Mukamel, E.A.; Nery, J.R.; Urich, M.; Puddifoot, C.A.; Johnson, N.D.; Lucero, J.; Huang, Y.; Dwork, A.J.; Schultz, M.D.; et al. Global epigenomic reconfiguration during mammalian brain development. Science 2013, 341, 1237905. [Google Scholar] [CrossRef] [PubMed]
- Maunakea, A.K.; Nagarajan, R.P.; Bilenky, M.; Ballinger, T.J.; D’Souza, C.; Fouse, S.D.; Johnson, B.E.; Hong, C.; Nielsen, C.; Zhao, Y.; et al. Conserved role of intragenic DNA methylation in regulating alternative promoters. Nature 2010, 466, 253–257. [Google Scholar] [CrossRef] [PubMed]
- Marina, R.J.; Sturgill, D.; Bailly, M.A.; Thenoz, M.; Varma, G.; Prigge, M.F.; Nanan, K.K.; Shukla, S.; Haque, N.; Oberdoerffer, S. TET-catalyzed oxidation of intragenic alternative splicing. EMBO J. 2016, 35, 335–355. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; Kavak, E.; Gregory, M.; Imashimizu, M.; Shutinoski, B.; Kashlev, M. CTCF-promoted RNA polymerase II pausing links DNA methylation to splicing. Nature 2011, 479, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Bachman, M.; Uribe-Lewis, S.; Yang, X.; Williams, M.; Murrell, A.; Balasubramanian, S. 5-Hydroxymethylcytosine is a predominantly stable DNA modification. Nat. Chem. 2014, 6, 1049–1055. [Google Scholar] [CrossRef] [PubMed]
- Alaghband, Y.; Bredy, T.W.; Wood, M.A. The role of active DNA demethylation and Tet enzyme function in memory formation and cocaine action. Neurosci. Lett. 2016, 625, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.U.; Su, Y.; Zhong, C.; Ming, G.; Song, H. Emerging roles of TET proteins and 5-hydroxymethylcytosines in active DNA demethylation and beyond. Cell Cycle 2011, 10, 2662–2668. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Shin, J.; Zhong, C.; Wang, S.; Roychowdhury, P.; Lim, J.; Kim, D.; Ming, G.; Song, H. Neuronal activity modifies the chromatin accessibility landscape in the adult brain. Nat. Neurosci. 2017, 20, 476–483. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.U.; Ma, D.K.; Mo, H.; Ball, M.P.; Jang, M.-H.; Bonaguidi, M.A.; Balazer, J.A.; Eaves, H.L.; Xie, B.; Ford, E.; et al. Neuronal activity modifies the DNA methylation landscape in the adult brain. Nat. Neurosci. 2011, 14, 1345–1351. [Google Scholar] [CrossRef] [PubMed]
- Maag, J.L.V.; Kaczorowski, D.C.; Panja, D.; Peters, T.J.; Bramham, C.R.; Wibrand, K.; Dinger, M.E. Widespread promoter methylation of synaptic plasticity genes in long-term potentiation in the adult brain in vivo. BMC Genom. 2017, 18, 250. [Google Scholar] [CrossRef] [PubMed]
- Valjent, E.; Corvol, J.C.; Pages, C.; Besson, M.J.; Maldonado, R.; Caboche, J. Involvement of the extracellular signal-regulated kinase cascade for cocaine-rewarding properties. J. Neurosci. 2000, 20, 8701–8709. [Google Scholar] [PubMed]
- Savell, K.E.; Gallus, N.V.N.; Simon, R.C.; Brown, J.A.; Revanna, J.S.; Osborn, M.K.; Song, E.Y.; O’Malley, J.J.; Stackhouse, C.T.; Norvil, A.; et al. Extra-coding RNAs regulate neuronal DNA methylation dynamics. Nat. Commun. 2016, 7, 12091. [Google Scholar] [CrossRef] [PubMed]
- Wiegert, J.S.; Bading, H. Activity-dependent calcium signaling and ERK-MAP kinases in neurons: A link to structural plasticity of the nucleus and gene transcription regulation. Cell Calcium 2011, 49, 296–305. [Google Scholar] [CrossRef] [PubMed]
- Anier, K.; Malinovskaja, K.; Aonurm-Helm, A.; Zharkovsky, A.; Kalda, A. DNA methylation regulates cocaine-induced behavioral sensitization in mice. Neuropsychopharmacology 2010, 35, 2450–2461. [Google Scholar] [CrossRef] [PubMed]
- Anier, K.; Zharkovsky, A.; Kalda, A. S-adenosylmethionine modifies cocaine-induced DNA methylation and increases locomotor sensitization in mice. Int. J. Neuropsychopharmacol. 2013, 16, 2053–2066. [Google Scholar] [CrossRef] [PubMed]
- Laplant, Q.; Vialou, V.; Covington, H.E.; Dumitriu, D.; Feng, J.; Warren, B.L.; Maze, I.; Dietz, D.M.; Watts, E.L.; Iñiguez, S.D.; et al. DNMT3A regulates emotional behavior and spine plasticity in the nucleus accumbens. Nat. Neurosci. 2010, 13, 1137–1143. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Li, Y.; Wang, D.; Wei, C.; Yang, X.; Sui, N. Effect of 5-aza-2-deoxycytidine microinjecting into hippocampus and prelimbic cortex on acquisition and retrieval of cocaine-induced place preference in C57BL/6 mice. Eur. J. Pharmacol. 2010, 642, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Tian, W.; Zhao, M.; Li, M.; Song, T.; Zhang, M.; Quan, L.; Li, S.; Sun, Z.S. Reversal of cocaine-conditioned place preference through methyl supplementation in mice: Altering global DNA methylation in the prefrontal cortex. PLoS ONE 2012, 7, e33435. [Google Scholar] [CrossRef] [PubMed]
- Wright, K.N.; Hollis, F.; Duclot, F.; Dossat, A.M.; Strong, C.E.; Francis, T.C.; Mercer, R.; Feng, J.; Dietz, D.M.; Lobo, M.K.; et al. Methyl Supplementation Attenuates Cocaine-Seeking Behaviors and Cocaine-Induced c-Fos Activation in a DNA Methylation-Dependent Manner. J. Neurosci. 2015, 35, 8948–8958. [Google Scholar] [CrossRef] [PubMed]
- Massart, R.; Barnea, R.; Dikshtein, Y.; Suderman, M.; Meir, O.; Hallett, M.; Kennedy, P.; Nestler, E.J.; Szyf, M.; Yadid, G. Role of DNA methylation in the nucleus accumbens in incubation of cocaine craving. J. Neurosci. 2015, 35, 8042–8058. [Google Scholar] [CrossRef] [PubMed]
- Okano, M.; Bell, D.W.; Haber, D.A.; Li, E. DNA methyltransferases DNMT3A and DNMT3B are essential for de novo methylation and mammalian development. Cell 1999, 99, 247–257. [Google Scholar] [CrossRef]
- Hsieh, C.L. In vivo activity of murine de novo methyltransferases, DNMT3A and DNMT3B. Mol. Cell. Biol. 1999, 19, 8211–8218. [Google Scholar] [CrossRef] [PubMed]
- Li, E. Chromatin modification and epigenetic reprogramming in mammalian development. Nat. Rev. Genet. 2002, 3, 662–673. [Google Scholar] [CrossRef] [PubMed]
- Romeo, K.; Louault, Y.; Cantaloube, S.; Loiodice, I.; Almouzni, G.; Quivy, J.P. The SENP7 SUMO-Protease presents a module of two HP1 interaction motifs that locks HP1 protein at pericentric heterochromatin. Cell Rep. 2015, 10, 771–782. [Google Scholar] [CrossRef] [PubMed]
- Turek-Plewa, J.; Jagodzinski, P.P. The role of mammalian DNA methyltransferases in the regulation of gene expression. Cell. Mol. Biol. Lett. 2005, 10, 631–647. [Google Scholar] [PubMed]
- Jeltsch, A.; Jurkowska, R.Z. New concepts in DNA methylation. Trends Biochem. Sci. 2014, 39, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Fatemi, M.; Hermann, A.; Gowher, H.; Jeltsch, A. DNMT3A and DNMT1 functionally cooperate during de novo methylation of DNA. Eur. J. Biochem. 2002, 269, 4981–4984. [Google Scholar] [CrossRef] [PubMed]
- Heiman, M.; Schaefer, A.; Gong, S.; Peterson, J.D.; Day, M.; Ramsey, K.E.; Suárez-Fariñas, M.; Schwarz, C.; Stephan, D.A.; Surmeier, D.J.; et al. Translational Profiling Approach for the Molecular Characterization of CNS Cell Types. Cell 2008, 135, 738–748. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Shao, N.; Szulwach, K.E.; Vialou, V.; Huynh, J.; Zhong, C.; Le, T.; Ferguson, D.; Cahill, M.E.; Li, Y.; et al. Role of Tet1 and 5-hydroxymethylcytosine in cocaine action. Nat. Neurosci. 2015, 18, 536–544. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.L.; Periyasamy, P.; Liao, K.; Kook, Y.H.; Niu, F.; Callen, S.E.; Buch, S. Cocaine-mediated downregulation of microglial miR-124 expression involves promoter DNA methylation. Epigenetics 2016, 11, 819–830. [Google Scholar] [CrossRef] [PubMed]
- Cassel, S.; Carouge, D.; Gensburger, C.; Anglard, P.; Burgun, C.; Dietrich, J.-B.; Aunis, D.; Zwiller, J. Fluoxetine and cocaine induce the epigenetic factors MeCP2 and MBD1 in adult rat brain. Mol. Pharmacol. 2006, 70, 487–492. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.V.; Rodriguiz, R.M.; Hutchinson, A.N.; Kim, I.-H.; Wetsel, W.C.; West, A.E. MeCP2 in the nucleus accumbens contributes to neural and behavioral responses to psychostimulants. Nat. Neurosci. 2010, 13, 1128–1136. [Google Scholar] [CrossRef] [PubMed]
- Im, H.-I.; Hollander, J.A.; Bali, P.; Kenny, P.J. MeCP2 controls BDNF expression and cocaine intake through homeostatic interactions with microRNA-212. Nat. Neurosci. 2010, 13, 1120–1127. [Google Scholar] [CrossRef] [PubMed]
- Mao, L.; Horton, E.; Guo, M.; Xue, B.; Jin, D.; Fibuch, E.E.; Wang, J.Q. Cocaine increases phosphorylation of MeCP2 in the rat striatum in vivo : A differential role of NMDA receptors. Neurochem. Int. 2012, 59, 610–617. [Google Scholar] [CrossRef] [PubMed]
- Baker-Andresen, D.; Zhao, Q.; Li, X.; Jupp, B.; Chesworth, R.; Lawrence, A.J.; Bredy, T.W. Persistent variations in neuronal DNA methylation following cocaine self-administration and protracted abstinence in mice. Neuroepigenetics 2015, 4, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kozlenkov, A.; Roussos, P.; Timashpolsky, A.; Barbu, M.; Rudchenko, S.; Bibikova, M.; Klotzle, B.; Byne, W.; Lyddon, R.; Di Narzo, A.F.; et al. Differences in DNA methylation between human neuronal and glial cells are concentrated in enhancers and non-CpG sites. Nucleic Acids Res. 2014, 42, 109–127. [Google Scholar] [CrossRef] [PubMed]
- Kristiansen, L.V.; Bannon, M.J.; Meador-Woodruff, J.H. Expression of Transcripts for Myelin Related Genes in Postmortem Brain from Cocaine Abusers. Neurochem. Res. 2009, 34, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Gigek, C.O.; Chen, E.S.; Smith, M.A.C. Methyl-CpG-Binding Protein (MBD) Family: Epigenomic Read-Outs Functions and Roles in Tumorigenesis and Psychiatric Diseases. J. Cell. Biochem. 2016, 117, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Skene, P.J.; Illingworth, R.S.; Webb, S.; Kerr, A.R.W.; James, K.D.; Turner, D.J.; Andrews, R.; Bird, A.P. Neuronal MeCP2 Is Expressed at Near Histone-Octamer Levels and Globally Alters the Chromatin State. Mol. Cell 2010, 37, 457–468. [Google Scholar] [CrossRef] [PubMed]
- Zoghbi, H.Y.; Amir, R.E.; Van den Veyver, I.B.; Wan, M.; Tran, C.Q.; Francke, U. Rett syndrome is caused by mutations in X-linked MECP2, encoding methyl-CpG-binding protein 2. Nat. Genet. 1999, 23, 185–188. [Google Scholar] [CrossRef] [PubMed]
- Ausió, J. MeCP2 and the enigmatic organization of brain chromatin. Implications for depression and cocaine addiction. Clin. Epigenet. 2016, 8, 58. [Google Scholar]
- Mellén, M.; Ayata, P.; Dewell, S.; Kriaucionis, S.; Heintz, N. MeCP2 Binds to 5hmC Enriched within Active Genes and Accessible Chromatin in the Nervous System. Cell 2012, 151, 1417–1430. [Google Scholar]
- Nakao, M.; Matsui, S.; Yamamoto, S.; Okumura, K.; Shirakawa, M.; Fujita, N. Regulation of transcription and chromatin by methyl-CpG binding protein MBD1. Brain Dev. 2001, 23, S174–S176. [Google Scholar] [CrossRef]
- Pol Bodetto, S.; Carouge, D.; Fonteneau, M.; Dietrich, J.B.; Zwiller, J.; Anglard, P. Cocaine represses protein phosphatase-1CB through DNA methylation and Methyl-CpG Binding Protein-2 recruitment in adult rat brain. Neuropharmacology 2013, 73, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Hong, E.J.; Cohen, S.; Zhao, W.-N.; Ho, H.-Y.H.; Schmidt, L.; Chen, W.G.; Lin, Y.; Savner, E.; Griffith, E.C.; et al. Brain-specific phosphorylation of MeCP2 regulates activity-dependent BDNF transcription, dendritic growth, and spine maturation. Neuron 2006, 52, 255–269. [Google Scholar] [CrossRef] [PubMed]
- Ito, S.; Shen, L.; Dai, Q.; Wu, S.C.; Collins, L.B.; Swenberg, J.A.; He, C.; Zhang, Y. Tet proteins can convert 5-methylcytosine to 5-formylcytosine and 5-carboxylcytosine. Science 2011, 333, 1300–1303. [Google Scholar] [CrossRef] [PubMed]
- Tahiliani, M.; Koh, K.P.; Shen, Y.; Pastor, W.A.; Bandukwala, H.; Brudno, Y.; Agarwal, S.; Iyer, L.M.; Liu, D.R.; Aravind, L.; et al. Conversion of 5-methylcytosine to 5-hydroxymethylcytosine in mammalian DNA by MLL partner TET1. Science 2009, 324, 930–935. [Google Scholar] [CrossRef] [PubMed]
- Gross, J.A.; Pacis, A.; Chen, G.G.; Barreiro, L.B.; Ernst, C.; Turecki, G. Characterizing 5-hydroxymethylcytosine in human prefrontal cortex at single base resolution. BMC Genom. 2015, 16, 672. [Google Scholar] [CrossRef] [PubMed]
- Fragou, D.; Zanos, P.; Kouidou, S.; Njau, S.; Kitchen, I.; Bailey, A.; Kovatsi, L. Effect of chronic heroin and cocaine administration on global DNA methylation in brain and liver. Toxicol. Lett. 2013, 218, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Fonteneau, M.; Filliol, D.; Anglard, P.; Befort, K.; Romieu, P.; Zwiller, J. Inhibition of DNA methyltransferases regulates cocaine self-administration by rats: A genome-wide DNA methylation study. Genes Brain Behav. 2016. [Google Scholar] [CrossRef] [PubMed]
- Carouge, D.; Host, L.; Aunis, D.; Zwiller, J.; Anglard, P. CDKL5 is a brain MeCP2 target gene regulated by DNA methylation. Neurobiol. Dis. 2010, 38, 414–424. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, D.A.; Huang, W.; Hamon, S.C.; Maili, L.; Witkin, B.M.; Fox, R.G.; Cunningham, K.; Moeller, F.G. Forced abstinence from cocaine self-administration is associated with DNA methylation changes in myelin genes in the corpus callosum: A preliminary study. Front. Psychiatry 2012, 3, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Pol Bodetto, S.; Romieu, P.; Sartori, M.; Tesone-Coelho, C.; Majchrzak, M.; Barbelivien, A.; Zwiller, J.; Anglard, P. Differential regulation of MeCP2 and PP1 in passive or voluntary administration of cocaine or food. Int. J. Neuropsychopharmacol. 2014, 1, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Tian, W.; Wang, J.; Zhang, K.; Teng, H.; Li, C.; Szyf, M.; Sun, Z.S.; Zhao, M. Demethylation of c-MYB binding site mediates upregulation of BDNF IV in cocaine-conditioned place preference. Sci. Rep. 2016, 6, 22087. [Google Scholar] [CrossRef] [PubMed]
- Khatri, P.; Drăghici, S. Ontological analysis of gene expression data: Current tools, limitations, and open problems. Bioinformatics 2005, 21, 3587–3595. [Google Scholar] [CrossRef] [PubMed]
- Zachariou, V.; Sgambato-Faure, V.; Sasaki, T.; Svenningsson, P.; Berton, O.; Fienberg, A.A.; Nairn, A.C.; Greengard, P.; Nestler, E.J. Phosphorylation of DARPP-32 at Threonine-34 is Required for Cocaine Action. Neuropsychopharmacology 2006, 31, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Hiroi, N.; Fienberg, A.A.; Haile, C.N.; Alburges, M.; Hanson, G.R.; Greengard, P.; Nestler, E.J. Neuronal and behavioural abnormalities in striatal function in DARPP-32-mutant mice. Eur. J. Neurosci. 1999, 11, 1114–1118. [Google Scholar] [CrossRef] [PubMed]
- Veremeyko, T.; Siddiqui, S.; Sotnikov, I.; Yung, A.; Ponomarev, E.D. IL-4/IL-13-Dependent and Independent Expression of miR-124 and Its Contribution to M2 Phenotype of Monocytic Cells in Normal Conditions and during Allergic Inflammation. PLoS ONE 2013, 8, e81774. [Google Scholar] [CrossRef] [PubMed]
- Lambert, B.L.; Bauer, C.R. Developmental and behavioral consequences of prenatal cocaine exposure: A review. J. Perinatol. 2012, 32, 819–828. [Google Scholar] [CrossRef] [PubMed]
- Novikova, S.I.; He, F.; Bai, J.; Cutrufello, N.J.; Lidow, M.S.; Undieh, A.S. Maternal cocaine administration in mice alters DNA methylation and gene expression in hippocampal neurons of neonatal and prepubertal offspring. PLoS ONE 2008, 3, e1919. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Hou, J.; Chen, B.; Shao, X.; Zhu, R.; Bu, Q.; Gu, H.; Li, Y.; Zhang, B.; Du, C.; et al. Prenatal cocaine exposure impairs cognitive function of progeny via insulin growth factor II epigenetic regulation. Neurobiol. Dis. 2015, 82, 54–65. [Google Scholar] [CrossRef] [PubMed]
- Barros, M.; Dempster, E.L.; Illott, N.; Chabrawi, S.; Maior, R.S.; Tomaz, C.; De Souza Silva, M.A.; Huston, J.P.; Mill, J.; Müller, C.P. Decreased methylation of the NK3 receptor coding gene (TACR3) after cocaine-induced place preference in marmoset monkeys. Addict. Biol. 2013, 18, 452–454. [Google Scholar] [CrossRef] [PubMed]
- Walton, E.; Hass, J.; Liu, J.; Roffman, J.L.; Bernardoni, F.; Roessner, V.; Kirsch, M.; Schackert, G.; Calhoun, V.; Ehrlich, S. Correspondence of DNA Methylation Between Blood and Brain Tissue and Its Application to Schizophrenia Research. Schizophr. Bull. 2016, 42, 406–414. [Google Scholar] [CrossRef] [PubMed]
- Mo, A.; Mukamel, E.A.; Davis, F.P.; Luo, C.; Henry, G.L.; Picard, S.; Urich, M.A.; Nery, J.R.; Sejnowski, T.J.; Lister, R.; et al. Epigenomic Signatures of Neuronal Diversity in the Mammalian Brain. Neuron 2015, 86, 1369–1384. [Google Scholar] [CrossRef] [PubMed]
- Kozlenkov, A.; Wang, M.; Roussos, P.; Rudchenko, S.; Barbu, M.; Bibikova, M.; Klotzle, B.; Dwork, A.J.; Zhang, B.; Hurd, Y.L.; Koonin, E.V.; Wegner, M.; Dracheva, S. Substantial DNA methylation differences between two major neuronal subtypes in human brain. Nucleic Acids Res. 2016, 44, 2593–2612. [Google Scholar] [CrossRef] [PubMed]
- Schübeler, D. Function and information content of DNA methylation. Nature 2015, 517, 321–326. [Google Scholar]
- Lev Maor, G.; Yearim, A.; Ast, G. The alternative role of DNA methylation in splicing regulation. Trends Genet. 2015, 31, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Patil, V.; Ward, R.L.; Hesson, L.B. The evidence for functional non-CpG methylation in mammalian cells. Epigenetics 2014, 9, 823–828. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.U.; Su, Y.; Shin, J.H.; Shin, J.; Li, H.; Xie, B.; Zhong, C.; Hu, S.; Le, T.; Fan, G.; et al. Distribution, recognition and regulation of non-CpG methylation in the adult mammalian brain. Nat. Neurosci. 2013, 17, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, V.K.; McGarvey, K.M.; Licchesi, J.D.F.; Ohm, J.E.; Herman, J.G.; Schübeler, D.; Baylin, S.B. PcG proteins, DNA methylation, and gene repression by chromatin looping. PLoS Biol. 2008, 6, 2911–2927. [Google Scholar] [CrossRef] [PubMed]
- Cedar, H.; Bergman, Y. Linking DNA methylation and histone modification: Patterns and paradigms. Nat. Rev. Genet. 2009, 10, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Gravina, S.; Dong, X.; Yu, B.; Vijg, J.; Gravina, S.; Ganapathi, S.; Vijg, J.; Feinberg, J.; Toner, C.; Greenland, P.; et al. Single-cell genome-wide bisulfite sequencing uncovers extensive heterogeneity in the mouse liver methylome. Genome Biol. 2016, 17, 150. [Google Scholar] [CrossRef] [PubMed]
- Clark, S.J.; Smallwood, S.A.; Lee, H.J.; Krueger, F.; Reik, W.; Kelsey, G. Genome-wide base-resolution mapping of DNA methylation in single cells using single-cell bisulfite sequencing (scBS-seq). Nat. Protoc. 2017, 12, 534–547. [Google Scholar] [CrossRef] [PubMed]
- Vojta, A.; Dobrinić, P.; Tadić, V.; Bočkor, L.; Korać, P.; Julg, B.; Klasić, M.; Zoldoš, V. Repurposing the CRISPR-Cas9 system for targeted DNA methylation. Nucleic Acids Res. 2016, 44, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Stepper, P.; Kungulovski, G.; Jurkowska, R.Z.; Chandra, T.; Krueger, F.; Reinhardt, R.; Reik, W.; Jeltsch, A.; Jurkowski, T.P. Efficient targeted DNA methylation with chimeric dCas9–DNMT3A–DNMT3L methyltransferase. Nucleic Acids Res. 2017, 45, 1703–1713. [Google Scholar] [CrossRef] [PubMed]
- McDonald, J.I.; Celik, H.; Rois, L.E.; Fishberger, G.; Fowler, T.; Rees, R.; Kramer, A.; Martens, A.; Edwards, J.R.; Challen, G.A. Reprogrammable CRISPR/Cas9-based system for inducing site-specific DNA methylation. Biol. Open 2016, 5, 866–874. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, S.R.; Cui, Y.; Lubecka, K.; Stefanska, B.; Irudayaraj, J. CRISPR-dCas9 mediated TET1 targeting for selective DNA demethylation at BRCA1 promoter. Oncotarget 2014, 7, 46545–46556. [Google Scholar] [CrossRef] [PubMed]
- Vassoler, F.M.; White, S.L.; Schmidt, H.D.; Sadri-vakili, G. Epigenetic Inheritance of a Cocaine Resistance Phenotype. Nat. Neurosci. 2013, 16, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Nagy, C.; Turecki, G. Transgenerational epigenetic inheritance: An open discussion. Epigenomics 2015, 7, 781–790. [Google Scholar] [CrossRef] [PubMed]
- Hope, B.T.; Skinner, M.K.; Kenny, P.J.; Akbarian, S. Exploring the epigenetics of cocaine resistance. Nat. Med. 2013, 19, 136–137. [Google Scholar] [PubMed]
- Heyward, F.D.; Sweatt, J.D. DNA Methylation in Memory Formation. Neuroscientist 2015, 21, 475–489. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Ref | Species * | Tissue ** | Paradigm | Withdrawal | Summary Finding |
|---|---|---|---|---|---|
| Methyltransferases and Dioxygenases | |||||
| [46] | M | Striatum | Injection | 4 h | ↑ Dnmt3a mRNA in D1-MSNs only after chronic injection |
| [33] | M | NAc; Hipp | Injection | 24 h | acute injection ↑ Dnmt3a/b mRNA in NAc and Dnmt3b in Hipp |
| repeat injection had no effect on Dnmt expression | |||||
| 1.5 h | acute injection ↑ Dnmt3a mRNA in NAc and Dnmt3b in Hipp | ||||
| chronic injection ↑ Dnmt3a mRNA in Hipp | |||||
| [34] | M | NAc | Self Admin | 24 h or 28 days | Biphasic expression of Dnmt3a mRNA (↓ 24 h but ↑ 28 days withdrawal) |
| Injection | 28 days | ↑ DNMT3a mRNA | |||
| [36] | M | PFC | Self Admin | 2 h | ↑ Dnmt3a mRNA and ↓ Dnmt3b mRNA and protein |
| [47] | M | NAc | Injection | 24 h | ↓ Tet1 mRNA and protein |
| H | NAc | Post mortem | N/A | ↓ TET1 mRNA | |
| [37] | R | NAc; PFC | Self Admin | 0 h | ↑ Dnmt3a/b mRNA expression in NAc only |
| [48] | M | BV-2 cells | 24 h Exposure | N/A | ↑ Dnmt1 and Dnmt3a protein |
| R | Microglia | 24 h Exposure | N/A | ↑ Dnmt1 protein and all Dnmt mRNA | |
| M | Microglia | Injection | 1 h | ↑ all Dnmt mRNA | |
| Methyl-Binding Proteins | |||||
| [49] | R | DCPu; FC; DG | Injection | 15 h | ↑ Mecp2 and Mbd1 positive cells in all areas |
| [50] | M | NAc | Injection | 2 h | ↑ phosphorylated Mecp2 |
| [51] | R | DCPu; PFC; Hipp | Self Admin | 24 h | ↑ Mecp2 protein and positive neurons in DCPu with extended access |
| ↓ Mecp2 protein a in PFC with extended access | |||||
| ↑ Mecp2 protein in Hipp with restricted or extended access | |||||
| [52] | R | CPu; Nac; PFC | Injection | 20 min | ↑ Mecp2 phosphorylation in NAc |
| 60 min | ↑ Mecp2 phosphorylation in CPu | ||||
| [53] | R | CPu; PFC | Injection/Self Admin | 5-15 h | ↑ Mecp2 expressing cells with no change in mRNA |
| Ref | Species * | Tissue ** | Paradigm # | Withdrawal | Technology ## | Summary Finding |
|---|---|---|---|---|---|---|
| Global Methylation Level | ||||||
| [36] | M | NAc; PFC | CPP | Not reported | HPLC | Slight decrease in 5mC in PFC |
| [67] | M | WB | Injection | 1 h | HPLC | No differences |
| [47] | M | NAc | Injection | 24 h | LC-ESI-MS/MS | No differences in 5mC nor 5hmC |
| Cocaine Methylome | ||||||
| [53] | M | PFC | Injection Self-Admin | 0–2 h | MBD-seq | distinct patterns of DNA methylation after active and passive cocaine |
| 29 persistent DMRs after self-administration (↑ 24 and ↓ 5) | ||||||
| [47] | M | NAc | Injection | 24 h | hMeDip-seq | 5hmC alterations at enhancer regions and alternative spliced sites |
| [38] | R | NAc | Self Admin | 1 or 30 days | MeDip-Array | Dynamic (hypo and hyper-) methylation after 1 day withdrawal |
| Locus-specific enhancement or reversal of early methylation changes after 30 days | ||||||
| Cue-reinstatement reversal of many withdrawal induced changes | ||||||
| [68] | R | PFC | Self Admin | 24 h | MBD-seq | More hyper- than hypomethyated DMRs at higher differential methylation ratios |
| Differentially Methylated Loci | ||||||
| [32] | M | NAc | Injection | 24 h | MeDip Bis-qPCR | ↑ PP1c promoter methylation after acute and chronic cocaine |
| 1.5 h | MeDip-qPCR | ↓ fosB promoter methylation after acute and chronic cocaine | ||||
| [69] | R | Striatum | Injection | 15 h | Bis-seq | ↑ Cdkl5 promoter methylation |
| [37] | R | NAc | Self Admin | 0 h | Bis-seq | ↓ c-Fos promoter methylation |
| [70] | R | CC | Self Admin | 30 days | Bis-seq | ↓ Sox10 promoter methylation |
| [33] | M | NAc | Injection | 24 h | MeDip-qPCR | ↑ Scl17a7 and Cck promoter methylation and ↓ Gal, DNMT3a and DMNT3b promoter methylation |
| [71] | R | CPu | Injection | 12 h | MeDip-qPCR | ↑ PP1Cβ promoter methylation after chronic cocaine |
| [48] | M | Microglia | Injection | 1 h | Bis-seq | ↑ Mir124 promoter methylation |
| [72] | M | NAc | CPP | 2 h | Bis-seq | ↓ BDNF IV promoter at single CpG |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vaillancourt, K.; Ernst, C.; Mash, D.; Turecki, G. DNA Methylation Dynamics and Cocaine in the Brain: Progress and Prospects. Genes 2017, 8, 138. https://doi.org/10.3390/genes8050138
Vaillancourt K, Ernst C, Mash D, Turecki G. DNA Methylation Dynamics and Cocaine in the Brain: Progress and Prospects. Genes. 2017; 8(5):138. https://doi.org/10.3390/genes8050138
Chicago/Turabian StyleVaillancourt, Kathryn, Carl Ernst, Deborah Mash, and Gustavo Turecki. 2017. "DNA Methylation Dynamics and Cocaine in the Brain: Progress and Prospects" Genes 8, no. 5: 138. https://doi.org/10.3390/genes8050138
APA StyleVaillancourt, K., Ernst, C., Mash, D., & Turecki, G. (2017). DNA Methylation Dynamics and Cocaine in the Brain: Progress and Prospects. Genes, 8(5), 138. https://doi.org/10.3390/genes8050138