Sequence Composition and Evolution of Mammalian B Chromosomes

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Mammalian Genomes: Organization and Diversity
3. Mammalian B Chromosomes: Prevalence and DNA Content
4. Transcriptional Activity of DNA in Mammalian B Chromosomes
5. Where Do Mammalian B Chromosomes Reside in the Interphase Nucleus?
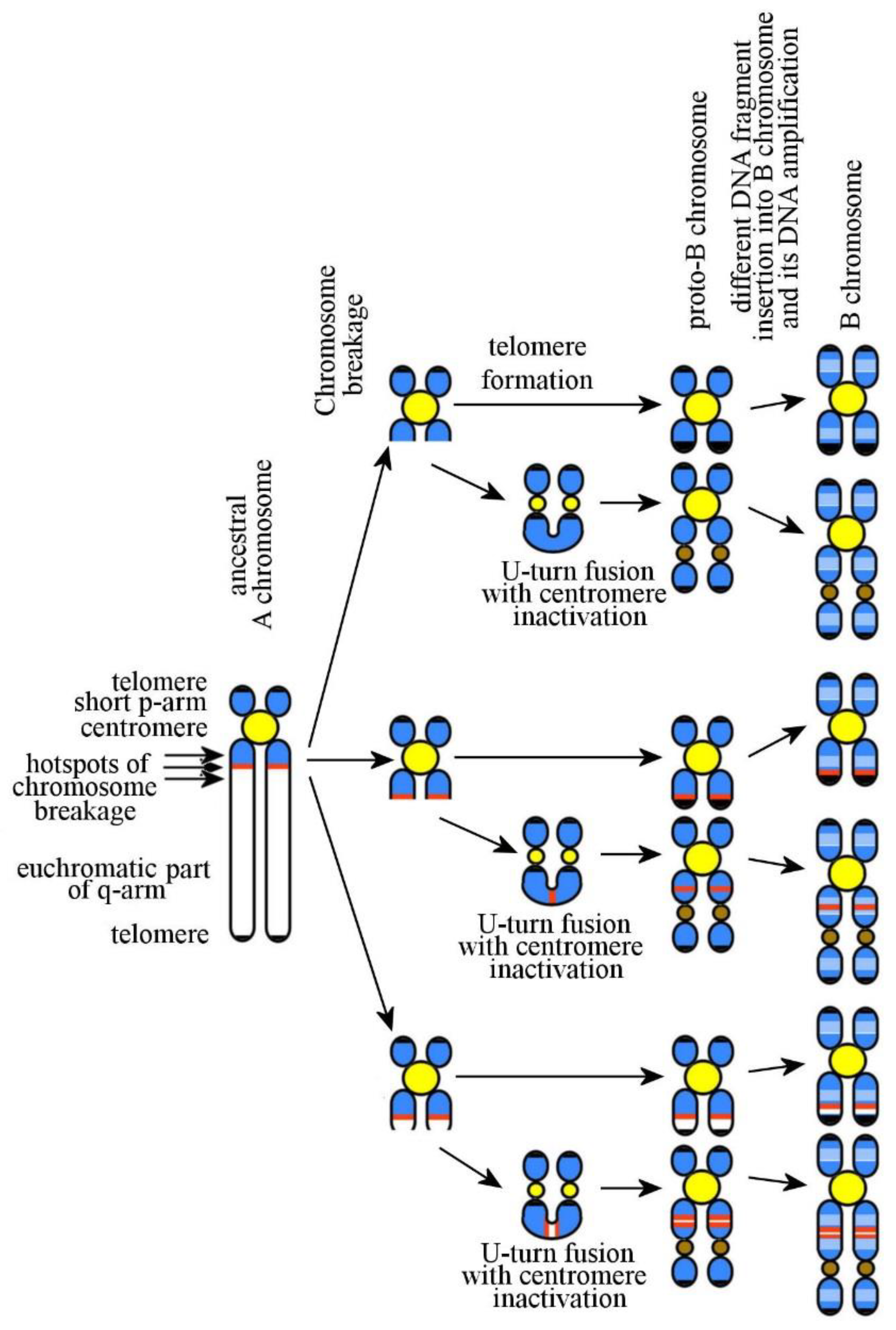
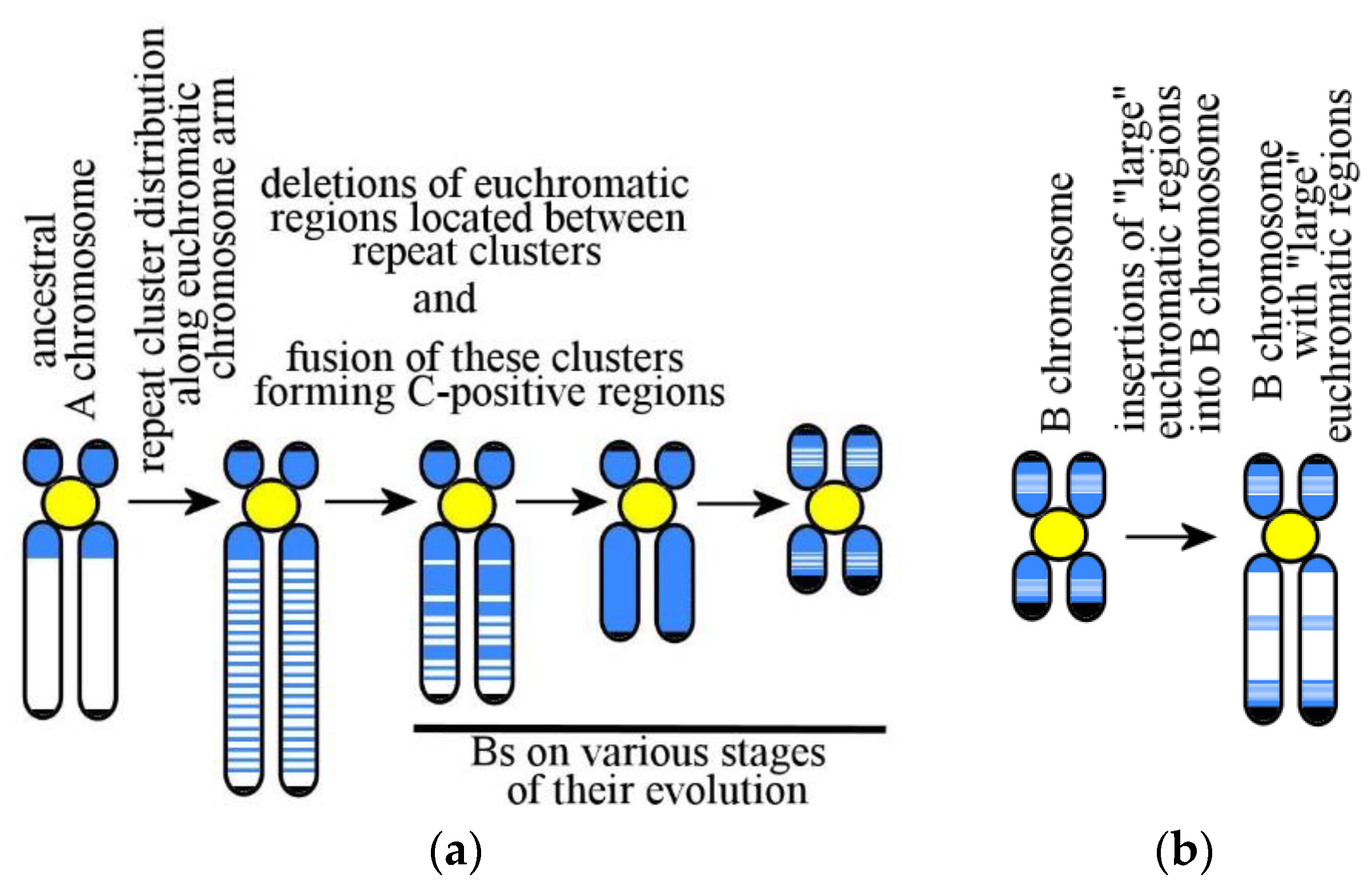
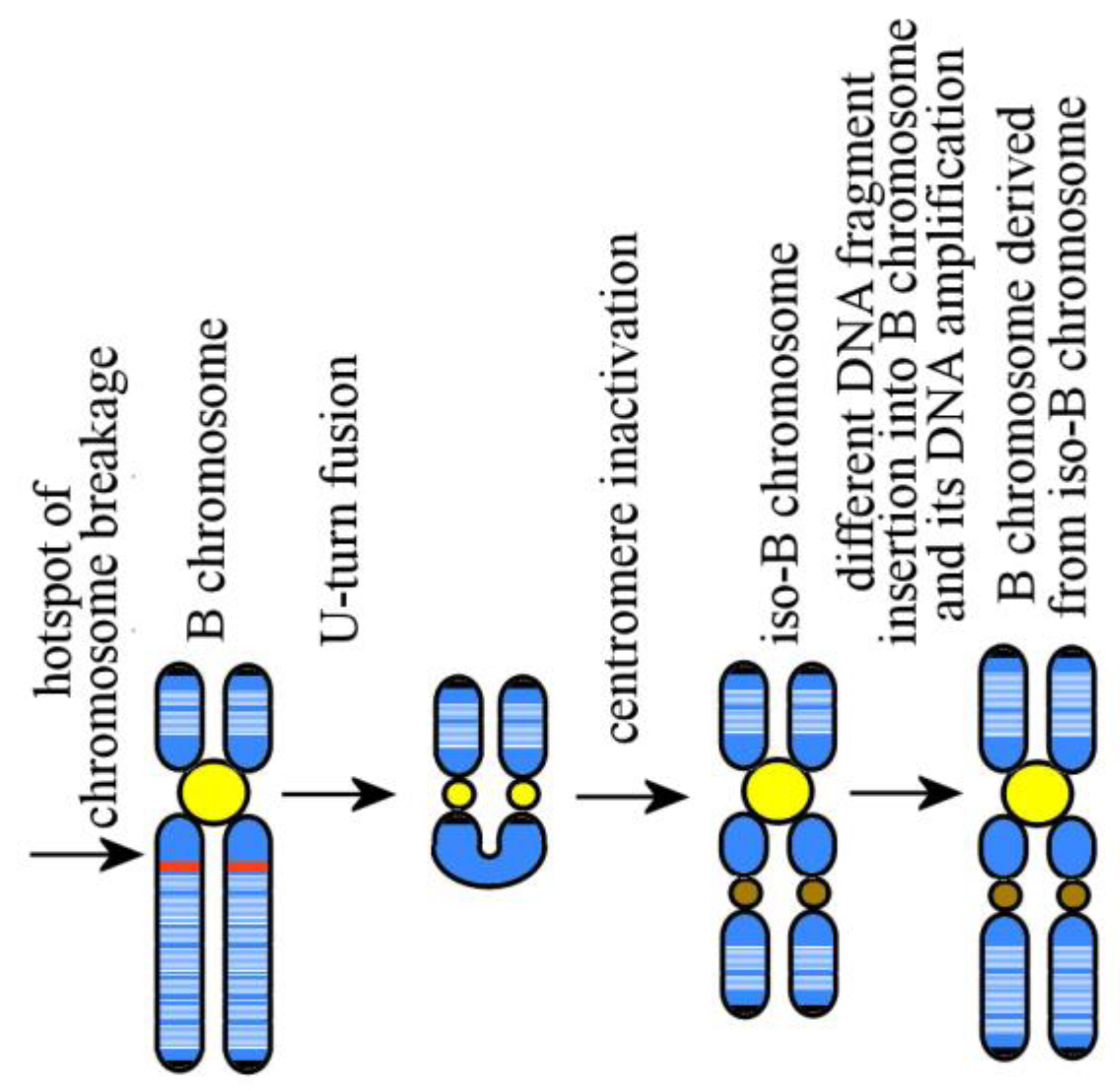
6. Mammalian B Chromosomes: Origin and Evolution
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wilson, E. The supernumerary chromosomes of Hemiptera. Science 1907, 26, 870–873. [Google Scholar] [CrossRef]
- Randolph, L.F. Types of supernumerary chromosomes in maize. Anat. Rec. 1928, 41, 102. [Google Scholar]
- Pigozzi, M.I.; Solari, A.J. Germ cell restriction and regular transmission of an accessory chromosome that mimics a sex body in the zebra finch, Taeniopygia guttata. Chromosom. Res. 1998, 6, 105–113. [Google Scholar] [CrossRef]
- Biederman, M.K.; Nelson, M.M.; Asalone, K.C.; Pedersen, A.L.; Saldanha, C.J.; Bracht, J.R. Discovery of the first germline-restricted gene by subtractive transcriptomic analysis in the zebra finch, Taeniopygia guttata. Curr. Biol. 2018, 28, 1620–1627. [Google Scholar] [CrossRef] [PubMed]
- Del Priore, L.; Pigozzi, M.I. Histone modifications related to chromosome silencing and elimination during male meiosis in Bengalese finch. Chromosoma 2014, 123, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Kapusta, A.; Suh, A.; Feschotte, C. Dynamics of genome size evolution in birds and mammals. Proc. Natl. Acad. Sci. USA 2017, 114, E1460–E1469. [Google Scholar] [CrossRef] [PubMed]
- D’Ambrosio, U.; Alonso-Lifante, M.P.; Barros, K.; Kovařík, A.; Mas de Xaxars, G.; Garcia, S. B-chrom: A database on B-chromosomes of plants, animals and fungi. New Phytol. 2017, 635–642. [Google Scholar] [CrossRef] [PubMed]
- Rubtsov, N.B.; Kartavtseva, I.V.; Roslik, G.V.; Karamysheva, T.V.; Pavlenko, M.V.; Iwasa, M.A.; Koh, H.S. Features of the B chromosome in Korean field mouse Apodemus peninsulae (Thomas, 1906) from Transbaikalia and the Far East identified by the FISH method. Russ. J. Genet. 2015, 51, 278–288. [Google Scholar] [CrossRef]
- Rubtsov, N.B.; Karamysheva, T.V.; Andreenkova, O.V.; Bochkaerev, M.N.; Kartavtseva, I.V.; Roslik, G.V.; Borissov, Y.M. Comparative analysis of micro and macro B chromosomes in the Korean field mouse Apodemus peninsulae (Rodentia, Murinae) performed by chromosome microdissection and FISH. Cytogenet. Genome Res. 2004, 106, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Camacho, J.P.M.; Sharbel, T.F.; Beukeboom, L.W. B-chromosome evolution. Philos. Trans. R. Soc. B Biol. Sci. 2000, 355, 163–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stitou, S.; Díaz De La Guardia, R.; Jiménez, R.; Burgos, M. Inactive ribosomal cistrons are spread throughout the B chromosomes of Rattus rattus (Rodentia, Muridae). Implications for their origin and evolution. Chromosom. Res. 2000, 8, 305–311. [Google Scholar] [CrossRef]
- Duke Becker, S.E.; Thomas, R.; Trifonov, V.A.; Wayne, R.K.; Graphodatsky, A.S.; Breen, M. Anchoring the dog to its relatives reveals new evolutionary breakpoints across 11 species of the Canidae and provides new clues for the role of B chromosomes. Chromosom. Res. 2011, 19, 685–708. [Google Scholar] [CrossRef] [PubMed]
- Makunin, A.I.; Rajičić, M.; Karamysheva, T.V.; Romanenko, S.A.; Druzhkova, A.S.; Blagojević, J.; Vujošević, M.; Rubtsov, N.B.; Graphodatsky, A.S.; Trifonov, V.A. Low-pass single-chromosome sequencing of human small supernumerary marker chromosomes (sSMCs) and Apodemus B chromosomes. Chromosoma 2018, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Rubtzov, N.B.; Borissov, Y.M.; Karamysheva, T.V.; Bochkarev, M.N. The mechanisms of formation and evolution of B chromosomes in Korean field mice Apodemus peninsulae (Mammalia, Rodentia). Russ. J. Genet. 2009, 45, 389–396. [Google Scholar] [CrossRef]
- Teruel, M.; Cabrero, J.; Perfectti, F.; Camacho, J.P.M. B chromosome ancestry revealed by histone genes in the migratory locust. Chromosoma 2010, 119, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Martis, M.M.; Klemme, S.; Banaei-Moghaddam, A.M.; Blattner, F.R.; Macas, J.; Schmutzer, T.; Scholz, U.; Gundlach, H.; Wicker, T.; Simkova, H.; et al. Selfish supernumerary chromosome reveals its origin as a mosaic of host genome and organellar sequences. Proc. Natl. Acad. Sci. USA 2012, 109, 13343–13346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolomiets, O.L.; Borbiev, T.E.; Safronova, L.D.; Borisov, Y.M.; Bogdanov, Y.F. Synaptonemal complex analysis of B-chromosome behavior in meiotic prophase I in the east-asiatic mouse Apodemus peninsulae (Muridae, Rodentia). Cytogenet. Genome Res. 1988, 48, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Karamysheva, T.V.; Torgasheva, A.A.; Yefremov, Y.R.; Bogomolov, A.G.; Liehr, T.; Borodin, P.M.; Rubtsov, N.B. Spatial organization of fibroblast and spermatocyte nuclei with different B-chromosome content in Korean field mouse, Apodemus peninsulae (Rodentia, Muridae). Genome 2017, 60, 815–824. [Google Scholar] [CrossRef] [PubMed]
- Plowman, A.B.; Bougourd, S.M. Selectively advantageous effects of B chromosomes on germination behaviour in Allium schoenoprasum L. Heredity 1994, 72, 587–593. [Google Scholar] [CrossRef]
- Blagojević, J.; Vujošević, M. B chromosomes and developmental homeostasis in the yellow-necked mouse, Apodemus flavicollis (Rodentia, Mammalia): Effects on nonmetric traits. Heredity 2004, 93, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Jojić, V.; Blagojević, J.; Vujošević, M. B chromosomes and cranial variability in yellow-necked field mice (Apodemus flavicollis). J. Mammal. 2011, 92, 396–406. [Google Scholar] [CrossRef]
- Yoshida, K.; Terai, Y.; Mizoiri, S.; Aibara, M.; Nishihara, H.; Watanabe, M.; Kuroiwa, A.; Hirai, H.; Hirai, Y.; Matsuda, Y.; et al. B chromosomes have a functional effect on female sex determination in Lake Victoria cichlid fishes. PLoS Genet. 2011, 7, e1002203. [Google Scholar] [CrossRef]
- Graphodatsky, A.S.; Trifonov, V.A.; Stanyon, R. The genome diversity and karyotype evolution of mammals. Mol. Cytogenet. 2011, 4, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Moriyama, Y.; Koshiba-Takeuchi, K. Significance of whole-genome duplications on the emergence of evolutionary novelties. Brief. Funct. Genom. 2018, 17, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Gallardo, M.H.; González, C.A.; Cebrián, I. Molecular cytogenetics and allotetraploidy in the red vizcacha rat, Tympanoctomys barrerae (Rodentia, Octodontidae). Genomics 2006, 88, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Evans, B.J.; Upham, N.S.; Golding, G.B.; Ojeda, R.A.; Ojeda, A.A. Evolution of the Largest Mammalian Genome. Genome Biol. Evol. 2017, 9, 1711–1724. [Google Scholar] [CrossRef] [PubMed]
- Perry, G.H.; Dominy, N.J.; Claw, K.G.; Lee, A.S.; Fiegler, H.; Redon, R.; Werner, J.; Villanea, F.A.; Mountain, J.L.; Misra, R.; et al. Diet and the evolution of human amylase gene copy number variation. Nat. Genet. 2007, 39, 1256–1260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibraimov, A.I.; Mirrakhimov, M.M.; Nazarenko, S.A.; Axenrod, E.I.; Akbanova, G.A. Human chromosomal polymorphism. I. Chromosomal Q polymorphism in Mongoloid populations of central Asia. Hum. Genet. 1982, 60, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ibraimov, A.I.; Mirrakhimov, M.M.; Axenrod, E.I.; Kurmanova, G.U. Human chromosomal polymorphism. IX. Further data on the possible selective value of chromosomal Q-heterochromatin material. Hum. Genet. 1986, 73, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Graphodatsky, A.S.; Kukekova, A.V.; Yudkin, D.V.; Trifonov, V.A.; Vorobieva, N.V.; Beklemisheva, V.R.; Perelman, P.L.; Graphodatskaya, D.A.; Trut, L.N.; Yang, F.; et al. The proto-oncogene C-KIT maps to canid B-chromosomes. Chromosom. Res. 2005, 13, 113–122. [Google Scholar] [CrossRef]
- Makunin, A.; Romanenko, S.; Beklemisheva, V.; Perelman, P.; Druzhkova, A.; Petrova, K.; Prokopov, D.; Chernyaeva, E.; Johnson, J.; Kukekova, A.; et al. Sequencing of supernumerary chromosomes of red fox and raccoon dog confirms a non-random gene acquisition by B chromosomes. Genes 2018, 9, 405. [Google Scholar] [CrossRef] [PubMed]
- Trifonov, V.A.; Dementyeva, P.V.; Larkin, D.M.; O’Brien, P.C.M.; Perelman, P.L.; Yang, F.; Ferguson-Smith, M.A.; Graphodatsky, A.S. Transcription of a protein-coding gene on B chromosomes of the Siberian roe deer (Capreolus pygargus). BMC Biol. 2013, 11, 1. [Google Scholar] [CrossRef] [PubMed]
- Vujosevic, M.; Zivkovic, S. Numerical chromosome polymorphism in Apodemus flaviocollis and A. sylvaticus (Mammalia: Rodentia) caused by supernumerary chromosomes. Acta Vet. 1987, 37, 115–122. [Google Scholar]
- Vujošević, M.; Blagojević, J. B chromosomes in populations of mammals. Cytogenet. Genome Res. 2004, 106, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Yonenaga-Yassuda, Y.; Maia, V.; L’Abbate, M. Two tandem fusions and supernumerary chromosomes in Nectomys squamipes (Cricetidae, Rodentia). Caryologia 1988, 41, 25–39. [Google Scholar] [CrossRef]
- Liehr, T. Benign and Pathological Chromosomal Imbalances: Microscopic and Submicroscopic Copy Number Variations (CNVs) in Genetics and Counseling; Academic Press: Cambridge, MS, USA, 2014; ISBN 9780124046313. [Google Scholar]
- McCarroll, S.A.; Altshuler, D.M. Copy-number variation and association studies of human disease. Nat. Genet. 2007, 39, S37–S42. [Google Scholar] [CrossRef] [PubMed]
- Nowakowska, B. Clinical interpretation of copy number variants in the human genome. J. Appl. Genet. 2017, 58, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Sati, S.; Cavalli, G. Chromosome conformation capture technologies and their impact in understanding genome function. Chromosoma 2017, 126, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Yudkin, D.V.; Trifonov, V.A.; Kukekova, A.V.; Vorobieva, N.V.; Rubtsova, N.V.; Yang, F.; Acland, G.M.; Ferguson-Smith, M.A.; Graphodatsky, A.S. Mapping of KIT adjacent sequences on canid autosomes and B chromosomes. Cytogenet. Genome Res. 2007, 116, 100–103. [Google Scholar] [CrossRef] [PubMed]
- Borisov, I.M.; Afanas’ev, A.G.; Lebedev, T.T.; Bochkarev, M.N. Multiplicity of B microchromosomes in a Siberian population of mice Apodemus peninsulae (2n = 48 + 4–30 B chromosomes). Russ. J. Genet. 2010, 46, 505–711. [Google Scholar] [CrossRef]
- Kartavtseva, I.V.; Roslik, G.V. A complex B chromosome system in the Korean field mouse, Apodemus peninsulae. Cytogenet. Genome Res. 2004, 106, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Liehr, T.; Claussen, U.; Starke, H. Small supernumerary marker chromosomes (sSMC) in humans. Cytogenet. Genome Res. 2004, 107, 55–67. [Google Scholar] [CrossRef] [PubMed]
- Liehr, T.; Weise, A. Frequency of small supernumerary marker chromosomes in prenatal, newborn, developmentally retarded and infertility diagnostics. Int. J. Mol. Med. 2007, 9, 719–731. [Google Scholar] [CrossRef]
- Liehr, T.; Mrasek, K.; Kosyakova, N.; Ogilvie, C.; Vermeesch, J.; Trifonov, V.; Rubtsov, N. Small supernumerary marker chromosomes (sSMC) in humans; are there B chromosomes hidden among them. Mol. Cytogenet. 2008, 1, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armanet, N.; Tosca, L.; Brisset, S.; Liehr, T.; Tachdjian, G. Small supernumerary marker chromosomes in human infertility. Cytogenet. Genome Res. 2015, 146, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Hashemzadeh-Chaleshtori, M.; Teimori, H.; Ghasemi-Dehkordi, P.; Jafari-Ghahfarokhi, H.; Moradi-Chaleshtori, M.; Liehr, T. Small supernumerary marker chromosomes and their correlation with specific syndromes. Adv. Biomed. Res. 2015, 4, 140. [Google Scholar] [CrossRef] [PubMed]
- Reddy, K.S.; Aradhya, S.; Meck, J.; Tiller, G.; Abboy, S.; Bass, H. A systematic analysis of small supernumerary marker chromosomes using array CGH exposes unexpected complexity. Genet. Med. 2013, 15, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Hamid Al-Rikabi, A.B.; Pekova, S.; Fan, X.; Jancuskova, T.; Liehr, T. Small supernumerary marker chromosome may provide information on dosage-insensitive pericentric regions in human. Curr. Genom. 2018, 19, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Horvath, J.E. Lessons from the human genome: transitions between euchromatin and heterochromatin. Hum. Mol. Genet. 2001, 10, 2215–2223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makunin, A.I.; Dementyeva, P.V.; Graphodatsky, A.S.; Volobouev, V.T.; Kukekova, A.V.; Trifonov, V.A. Genes on B chromosomes of vertebrates. Mol. Cytogenet. 2014, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Makunin, A.I.; Kichigin, I.G.; Larkin, D.M.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; Yang, F.; Proskuryakova, A.A.; Vorobieva, N.V.; Chernyaeva, E.N.; O’Brien, S.J.; et al. Contrasting origin of B chromosomes in two cervids (Siberian roe deer and grey brocket deer) unravelled by chromosome-specific DNA sequencing. BMC Genom. 2016, 17, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Wurster-Hill, D.H.; Ward, O.G.; Davis, B.H.; Park, J.P.; Moyzis, R.K.; Meyne, J. Fragile sites, telomeric DNA sequences, B chromosomes, and DNA content in raccoon dogs, Nyctereutes procyonoides, with comparative notes on foxes, coyote, wolf, and raccoon. Cytogenet. Genome Res. 1988, 49, 278–281. [Google Scholar] [CrossRef] [PubMed]
- Trifonov, V.A.; Perelman, P.L.; Kawada, S.I.; Iwasa, M.A.; Oda, S.I.; Graphodatsky, A.S. Complex structure of B-chromosomes in two mammalian species: Apodemus peninsulae (Rodentia) and Nyctereutes procyonoides (Carnivora). Chromosom. Res. 2002, 10, 109–116. [Google Scholar] [CrossRef]
- Szczerbal, I.; Switonski, M. B chromosomes of the Chinese raccoon dog (Nyctereutes procyonoides procyonoides Gray) contain inactive NOR-like sequences. Caryologia 2003, 56, 213–216. [Google Scholar] [CrossRef]
- Bugarski-Stanojevic, V.; Stamenković, G.; Blagojević, J.; Liehr, T.; Kosyakova, N.; Rajičić, M.; Vujošević, M. Exploring supernumeraries—A new marker for screening of B-chromosomes presence in the yellow necked mouse Apodemus flavicollis. PLoS ONE 2016, 11, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Groenen, M.A.M.; Archibald, A.L.; Uenishi, H.; Tuggle, C.K.; Takeuchi, Y.; Rothschild, M.F.; Rogel-Gaillard, C.; Park, C.; Milan, D.; Megens, H.J.; et al. Analyses of pig genomes provide insight into porcine demography and evolution. Nature 2012, 491, 393–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valente, G.T.; Conte, M.A.; Fantinatti, B.E.A.; Cabral-De-Mello, D.C.; Carvalho, R.F.; Vicari, M.R.; Kocher, T.D.; Martins, C. Origin and evolution of B chromosomes in the cichlid fish Astatotilapia latifasciata based on integrated genomic analyses. Mol. Biol. Evol. 2014, 31, 2061–2072. [Google Scholar] [CrossRef] [PubMed]
- Banaei-Moghaddam, A.M.; Meier, K.; Karimi-Ashtiyani, R.; Houben, A. Formation and expression of pseudogenes on the B chromosome of rye. Plant Cell 2013, 25, 2536–2544. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Domínguez, B.; Ruiz-Ruano, F.J.; Cabrero, J.; Corral, J.M.; López-León, M.D.; Sharbel, T.F.; Camacho, J.P.M. Protein-coding genes in B chromosomes of the grasshopper Eyprepocnemis plorans. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef]
- Sochorová, J.; Garcia, S.; Gálvez, F.; Symonová, R.; Kovařík, A. Evolutionary trends in animal ribosomal DNA loci: Introduction to a new online database. Chromosoma 2018, 127, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Banaei-Moghaddam, A.M.; Martis, M.M.; Macas, J.; Gundlach, H.; Himmelbach, A.; Altschmied, L.; Mayer, K.F.X.; Houben, A. Genes on B chromosomes: Old questions revisited with new tools. Biochim. Biophys. Acta 2015, 1849, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Karamysheva, T.V.; Andreenkova, O.V.; Bochkaerev, M.N.; Borissov, Y.M.; Bogdanchikova, N.; Borodin, P.M.; Rubtsov, N.B. B chromosomes of Korean field mouse Apodemus peninsulae (Rodentia, Murinae) analysed by microdissection and FISH. Cytogenet. Genome Res. 2002, 96, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Tanić, N.; Vujošević, M.; Dedović-Tanić, N.; Dimitrijević, B. Differential gene expression in yellow-necked mice Apodemus flavicollis (Rodentia, Mammalia) with and without B chromosomes. Chromosoma 2005, 113, 418–427. [Google Scholar] [CrossRef] [PubMed]
- Hall, L.E.; Mitchell, S.E.; O’Neill, R.J. Pericentric and centromeric transcription: A perfect balance required. Chromosom. Res. 2012, 20, 535–546. [Google Scholar] [CrossRef] [PubMed]
- Saksouk, N.; Simboeck, E.; Déjardin, J. Constitutive heterochromatin formation and transcription in mammals. Epigenet. Chromatin 2015, 8, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Jia, S.T.; Jia, S. New Insights into the regulation of heterochromatin. Trends Genet. 2016, 32, 284–294. [Google Scholar] [CrossRef] [PubMed]
- Enukashvily, N.I.; Ponomartsev, N.V. Mammalian satellite DNA: A speaking dumb. Adv. Protein Chem. Struct. Biol. 2013, 90, 31–65. [Google Scholar] [CrossRef] [PubMed]
- Eymery, A.; Horard, B.; el Atifi-Borel, M.; Fourel, G.; Berger, F.; Vitte, A.L.; Van den Broeck, A.; Brambilla, E.; Fournier, A.; Callanan, M.; et al. A transcriptomic analysis of human centromeric and pericentric sequences in normal and tumor cells. Nucleic Acids Res. 2009, 7, 6340–6354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.; Gilbert, D.M. Proliferation-dependent and cell cycle-regulated transcription of mouse pericentric heterochromatin. J. Cell Biol. 2007, 179, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Chen, E.S.; Zhang, K.; Nicolas, E.; Cam, H.P.; Zofall, M.; Grewal, S.I.S. Cell cycle control of centromeric repeat transcription and heterochromatin assembly. Nature 2008, 451, 734–737. [Google Scholar] [CrossRef] [PubMed]
- Kloc, A.; Zaratiegui, M.; Nora, E.; Martienssen, R. RNA interference guides histone modification during the S phase of chromosomal replication. Curr. Biol. 2008, 8, 490–495. [Google Scholar] [CrossRef] [PubMed]
- Motamedi, M.R.; Verdel, A.; Colmenares, S.U.; Gerber, S.A.; Gygi, S.P.; Moazed, D. Two RNAi complexes, RITS and RDRC, physically interact and localize to noncoding centromeric RNAs. Cell 2004, 119, 789–802. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, T.; Cam, H.; Verdel, A.; Moazed, D.; Grewal, S.I.S. From the cover: RNA-dependent RNA polymerase is an essential component of a self-enforcing loop coupling heterochromatin assembly to siRNA production. Proc. Natl. Acad. Sci. USA 2005, 102, 152–157. [Google Scholar] [CrossRef] [PubMed]
- Verdel, A.; Jia, S.; Gerber, S.; Sugiyama, T.; Gygi, S.; Grewal, S.I.S.; Moazed, D. RNAi-mediated targeting of heterochromatin by the RITS complex. Science 2004, 303, 672–676. [Google Scholar] [CrossRef] [PubMed]
- Bayne, E.H.; Bijos, D.A.; White, S.A.; de Lima Alves, F.; Rappsilber, J.; Allshire, R.C. A systematic genetic screen identifies new factors influencing centromeric heterochromatin integrity in fission yeast. Genome Biol. 2014, 15, 481. [Google Scholar] [CrossRef] [PubMed]
- Hong, E.J.E.; Villén, J.; Gerace, E.L.; Gygi, S.P.; Moazed, D. A cullin E3 ubiquitin ligase complex associates with Rik1 and the Clr4 histone H3-K9 methyltransferase and is required for RNAi-mediated heterochromatin formation. RNA Biol. 2005, 2, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Jia, S.; Kobayashi, R.; Grewal, S.I.S. Ubiquitin ligase component Cul4 associates with Clr4 histone methyltransferase to assemble heterochromatin. Nat. Cell Biol. 2005. [Google Scholar] [CrossRef] [PubMed]
- Horn, P.J.; Bastie, J.N.; Peterson, C.L. A Rik1-associated, cullin-dependent E3 ubiquitin ligase is essential for heterochromatin formation. Genes Dev. 2005, 19, 1705–1714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noma, K.I.; Sugiyama, T.; Cam, H.; Verdel, A.; Zofall, M.; Jia, S.; Moazed, D.; Grewal, S.I.S. RITS acts in cis to promote RNA interference-mediated transcriptional and post-transcriptional silencing. Nat. Genet. 2004, 36, 1174–1180. [Google Scholar] [CrossRef] [PubMed]
- Cremer, T.; Cremer, C. Chromosome territories, nuclear architecture and gene regulation in mammalian cells. Nat. Rev. Genet. 2001, 2, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Cremer, T.; Cremer, M. Chromosome Territories. Cold Spring Harb. Perspect. Biol. 2010, 2, a003889. [Google Scholar] [CrossRef] [PubMed]
- Lanctôt, C.; Cheutin, T.; Cremer, M.; Cavalli, G.; Cremer, T. Dynamic genome architecture in the nuclear space: Regulation of gene expression in three dimensions. Nat. Rev. Genet. 2007, 8, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Fritz, A.J.; Barutcu, A.R.; Martin-Buley, L.; Van Wijnen, A.J.; Zaidi, S.K.; Imbalzano, A.N.; Lian, J.B.; Stein, J.L.; Stein, G.S. Chromosomes at work: Organization of chromosome territories in the interphase nucleus. J. Cell. Biochem. 2016, 117, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Kociucka, B.; Sosnowski, J.; Kubiak, A.; Nowak, A.; Pawlak, P.; Szczerbal, I. Three-dimensional positioning of B chromosomes in fibroblast nuclei of the red fox and the Chinese raccoon dog. Cytogenet. Genome Res. 2013, 139, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Tanabe, H.; Küpper, K.; Ishida, T.; Neusser, M.; Mizusawa, H. Inter- and intra-specific gene-density-correlated radial chromosome territory arrangements are conserved in Old World monkeys. Cytogenet. Genome Res. 2005, 108, 255–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solovei, I.; Thanisch, K.; Feodorova, Y. How to rule the nucleus: divide et impera. Curr. Opin. Cell Biol. 2016, 40, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Tanabe, H.; Habermann, F.A.; Solovei, I.; Cremer, M.; Cremer, T. Non-random radial arrangements of interphase chromosome territories: Evolutionary considerations and functional implications. Mutat. Res. 2002, 504, 37–45. [Google Scholar] [CrossRef]
- Olmo, E. Nucleotype and cell size in vertebrates: A review. Basic Appl. Histochem. 1983, 27, 227–256. [Google Scholar] [PubMed]
- Gillooly, J.F.; Hein, A.; Damiani, R. Nuclear DNA content varies with cell size across human cell types. Cold Spring Harb. Perspect. Biol. 2015, 7, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Hetzer, M.W. The Nuclear Envelope. Cold Spring Harb. Perspect. Biol. 2010, 2, a000539. [Google Scholar] [CrossRef] [PubMed]
- Palestis, B.G.; Burt, A.; Jones, R.N.; Trivers, R. B chromosomes are more frequent in mammals with acrocentric karyotypes: Support for the theory of centromeric drive. Proc. R. Soc. B Biol. Sci. 2004, 271, 22–24. [Google Scholar] [CrossRef] [PubMed]
- Palestis, B.G.; Trivers, R.; Burt, A.; Jones, R.N. The distribution of B chromosomes across species. Cytogenet. Genome Res. 2004, 106, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Murmann, A.E.; Conrad, D.F.; Mashek, H.; Curtis, C.A.; Nicolae, R.I.; Ober, C.; Schwartz, S. Inverted duplications on acentric markers: Mechanism of formation. Hum. Mol. Genet. 2009, 15, 2241–2256. [Google Scholar] [CrossRef] [PubMed]
- Sheth, F.; Ewers, E.; Kosyakova, N.; Weise, A.; Sheth, J.; Patil, S.; Ziegler, M.; Liehr, T. A neocentric isochromosome Yp present as additional small supernumerary marker chromosome—Evidence against U-type exchange mechanism? Cytogenet. Genome Res. 2009, 125, 115–116. [Google Scholar] [CrossRef] [PubMed]
- Pennisi, E. Shaking up the tree of life. Science 2016, 354, 817–821. [Google Scholar] [CrossRef] [PubMed]
- Jetybayev, I.Y.; Bugrov, A.G.; Buleu, O.G.; Bogomolov, A.G.; Rubtsov, N.B. Origin and evolution of the neo-sex chromosomes in pamphagidae grasshoppers through chromosome fusion and following heteromorphization. Genes 2017, 8, 323. [Google Scholar] [CrossRef] [PubMed]
- Karamysheva, T.V.; Bogdanov, A.S.; Kartavtseva, I.V.; Likhoshvay, T.V.; Bochkarev, M.N.; Kolcheva, N.E.; Marochkina, V.V.; Rubtsov, N.B. Comparative FISH analysis of C-positive blocks of centromeric chromosomal regions of pygmy wood mice Sylvaemus uralensis (Rodentia, Muridae). Russ. J. Genet. 2010, 46, 712–724. [Google Scholar] [CrossRef]
- Rubtsov, N.B.; Karamysheva, T.V.; Bogdanov, A.S.; Likhoshvay, T.V.; Kartavtseva, I.V. Comparative FISH analysis of C-positive regions of chromosomes of wood mice (Rodentia, Muridae, Sylvaemus). Russ. J. Genet. 2011, 47, 1096–1110. [Google Scholar] [CrossRef]
- Rubtsov, N.B.; Karamysheva, T.V.; Bogdanov, A.S.; Kartavtseva, I.V.; Bochkarev, M.N.; Iwasa, M.A. Comparative analysis of DNA homology in pericentric regions of chromosomes of wood mice from genera Apodemus and Sylvaemus. Russ. J. Genet. 2015, 51, 1233–1242. [Google Scholar] [CrossRef]
- L’Abbate, A.; Macchia, G.; D’Addabbo, P.; Lonoce, A.; Tolomeo, D.; Trombetta, D.; Kok, K.; Bartenhagen, C.; Whelan, C.W.; Palumbo, O.; et al. Genomic organization and evolution of double minutes/homogeneously staining regions with MYC amplification in human cancer. Nucleic Acids Res. 2014, 42, 9131–9145. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.M.A.; Mahadevaiah, S.K.; Fernandez-Capetillo, O.; Nussenzweig, A.; Xu, X.; Deng, C.X.; Burgoyne, P.S. Silencing of unsynapsed meiotic chromosomes in the mouse. Nat. Genet. 2005, 37, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Baarends, W.M.; Wassenaar, E.; van der Laan, R.; Hoogerbrugge, J.; Sleddens-Linkels, E.; Hoeijmakers, J.H.J.; de Boer, P.; Grootegoed, J.A. Silencing of Unpaired Chromatin and Histone H2A Ubiquitination in Mammalian Meiosis. Mol. Cell. Biol. 2005, 25, 1041–1053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sciurano, R.; Rahn, M.; Rey-Valzacchi, G.; Solari, A.J. The asynaptic chromatin in spermatocytes of translocation carriers contains the histone variant γ-H2AX and associates with the XY body. Hum. Reprod. 2007, 22, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Sciurano, R.B.; Rahn, M.I.; Rey-Valzacchi, G.; Coco, R.; Solari, A.J. The role of asynapsis in human spermatocyte failure. Int. J. Androl. 2012, 35, 541–549. [Google Scholar] [CrossRef] [PubMed]
- Naumova, A.K.; Fayer, S.; Leung, J.; Boateng, K.A.; Camerini-Otero, R.D.; Taketo, T. Dynamics of Response to Asynapsis and Meiotic Silencing in Spermatocytes from Robertsonian Translocation Carriers. PLoS ONE 2013, 8, e75970. [Google Scholar] [CrossRef] [PubMed]
- Burgoyne, P.S.; Mahadevaiah, S.K.; Turner, J.M.A. The consequences of asynapsis for mammalian meiosis. Nat. Rev. Genet. 2009, 10, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Manterola, M.; Page, J.; Vasco, C.; Berríos, S.; Parra, M.T.; Viera, A.; Rufas, J.S.; Zuccotti, M.; Garagna, S.; Fernández-Donoso, R. A high incidence of meiotic silencing of unsynapsed chromatin is not associated with substantial pachytene loss in heterozygous male mice carrying multiple simple Robertsonian translocations. PLoS Genet. 2009, 5, e1000625. [Google Scholar] [CrossRef] [PubMed]



© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rubtsov, N.B.; Borisov, Y.M. Sequence Composition and Evolution of Mammalian B Chromosomes. Genes 2018, 9, 490. https://doi.org/10.3390/genes9100490
Rubtsov NB, Borisov YM. Sequence Composition and Evolution of Mammalian B Chromosomes. Genes. 2018; 9(10):490. https://doi.org/10.3390/genes9100490
Chicago/Turabian StyleRubtsov, Nikolay B., and Yury M. Borisov. 2018. "Sequence Composition and Evolution of Mammalian B Chromosomes" Genes 9, no. 10: 490. https://doi.org/10.3390/genes9100490
APA StyleRubtsov, N. B., & Borisov, Y. M. (2018). Sequence Composition and Evolution of Mammalian B Chromosomes. Genes, 9(10), 490. https://doi.org/10.3390/genes9100490