Chromosome Painting in Neotropical Long- and Short-Tailed Parrots (Aves, Psittaciformes): Phylogeny and Proposal for a Putative Ancestral Karyotype for Tribe Arini


 , ,
, , 

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Material and Methods
2.1. Samples, Cell Culture, and Chromosome Preparation
2.2. Fluorescent In Situ Hybridization
2.3. Phylogenetic Analysis
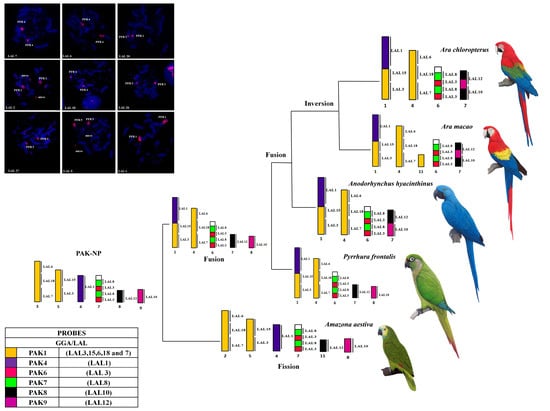
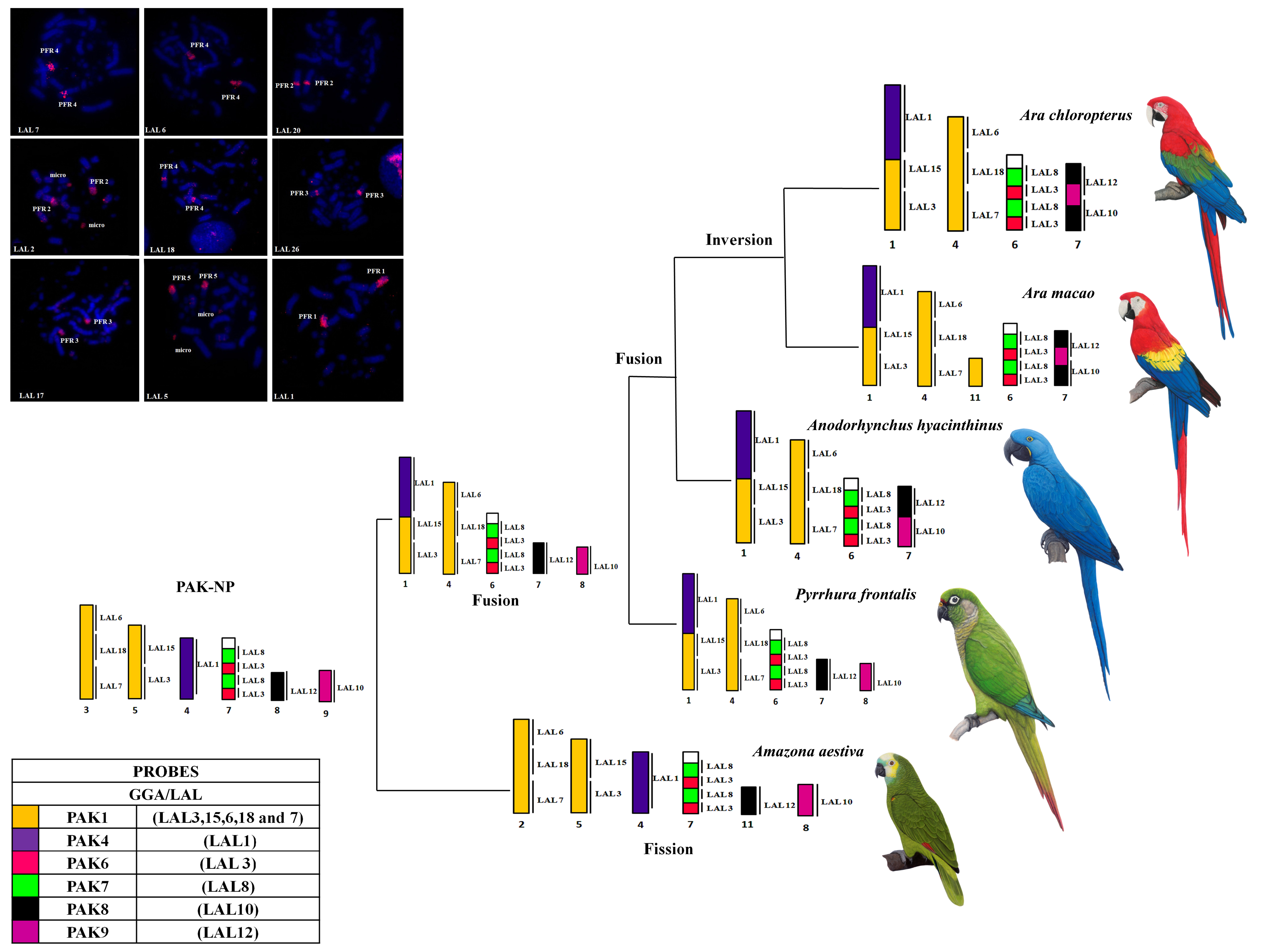
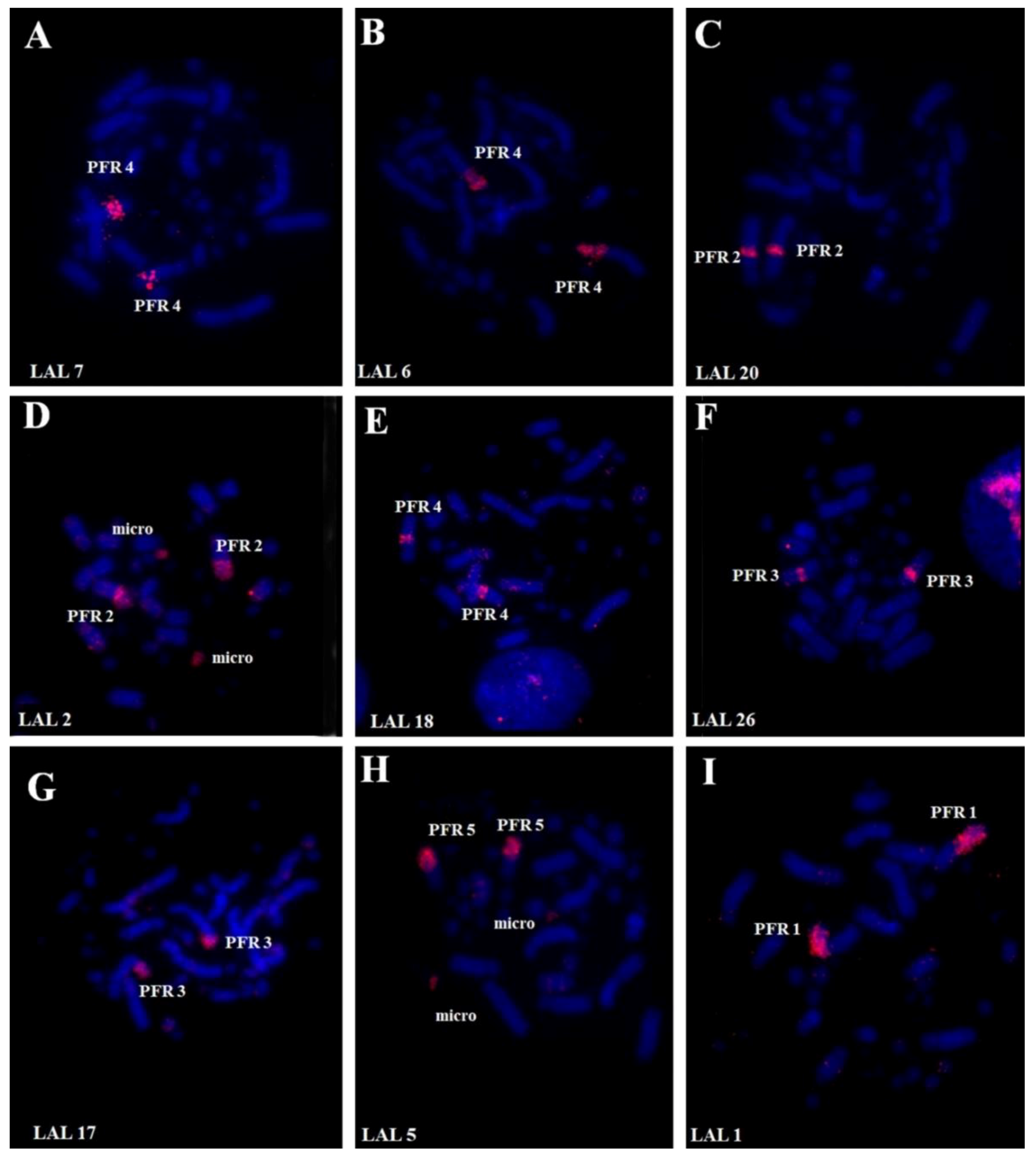
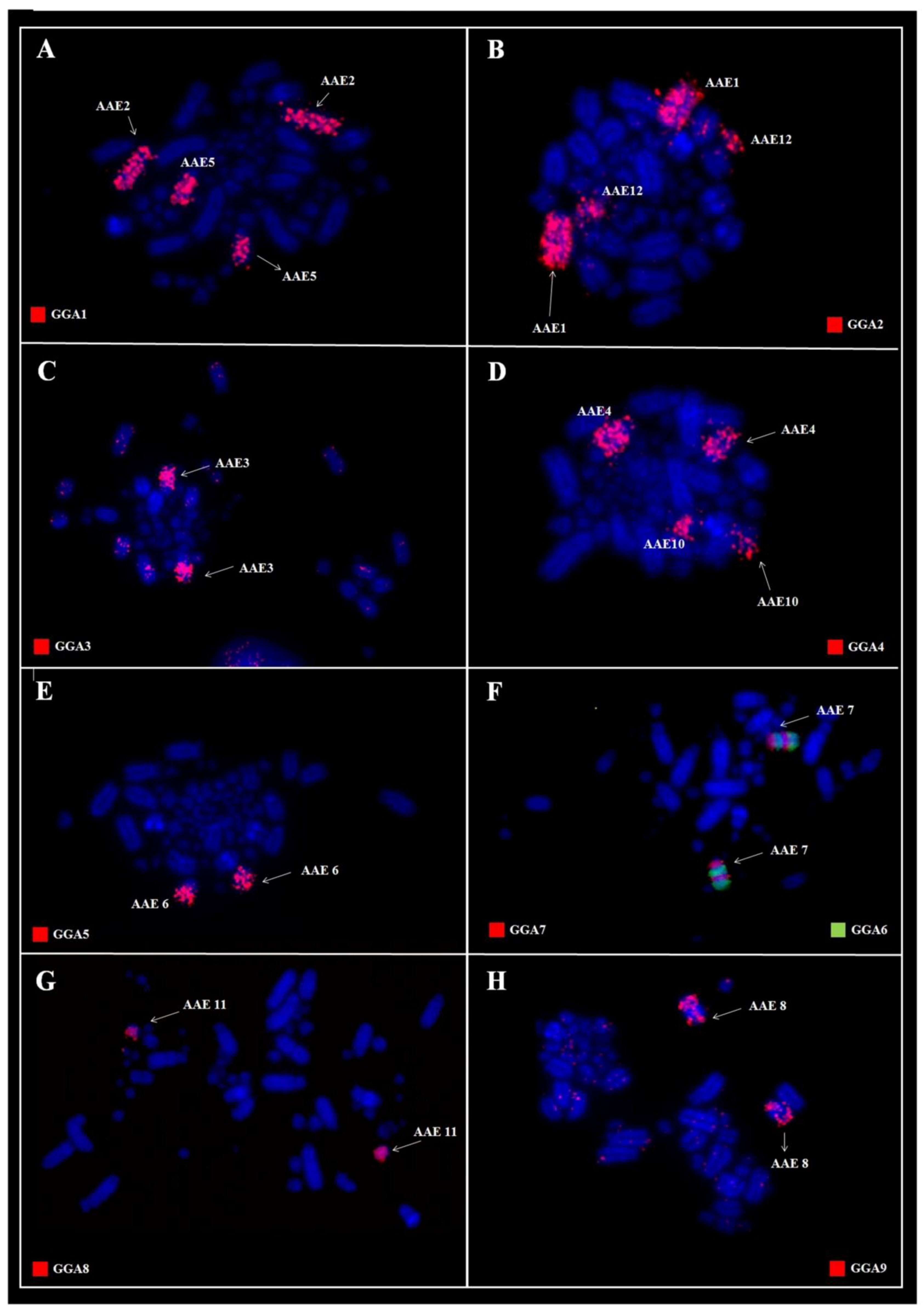
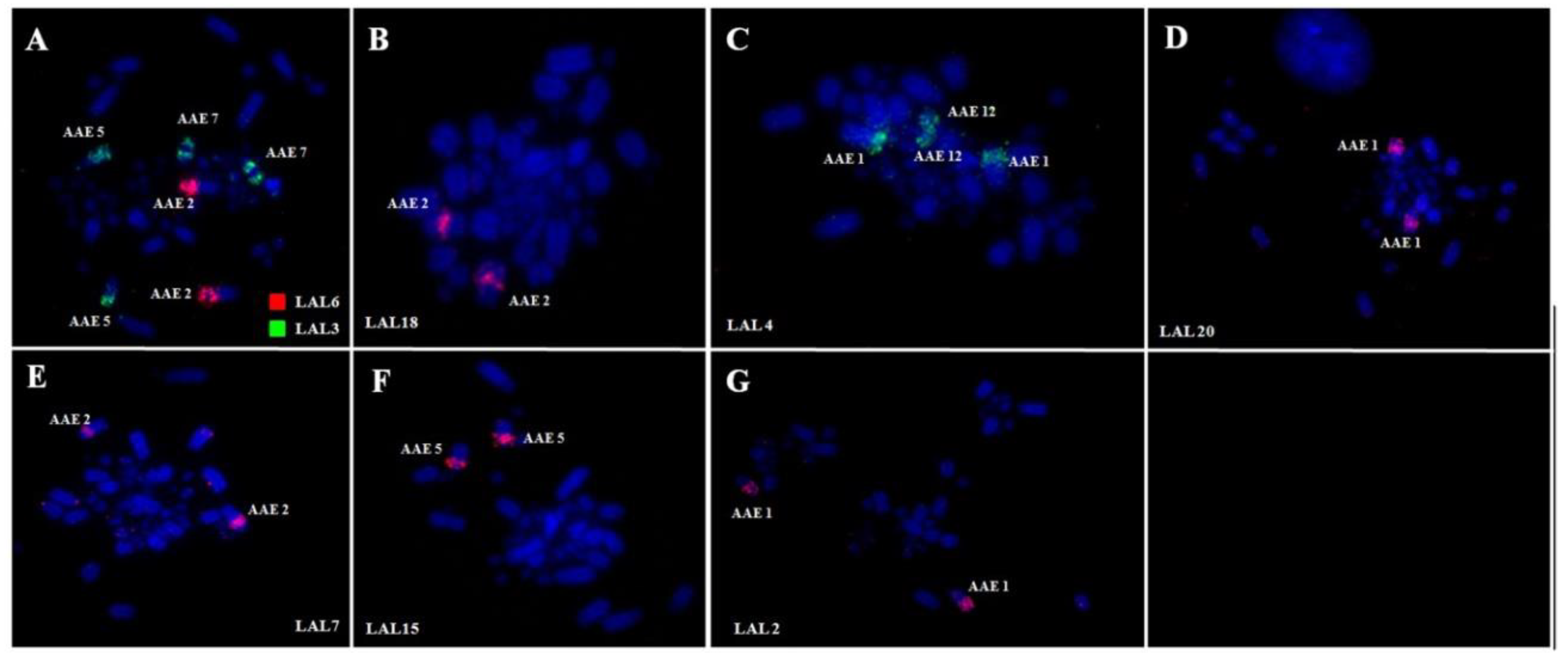
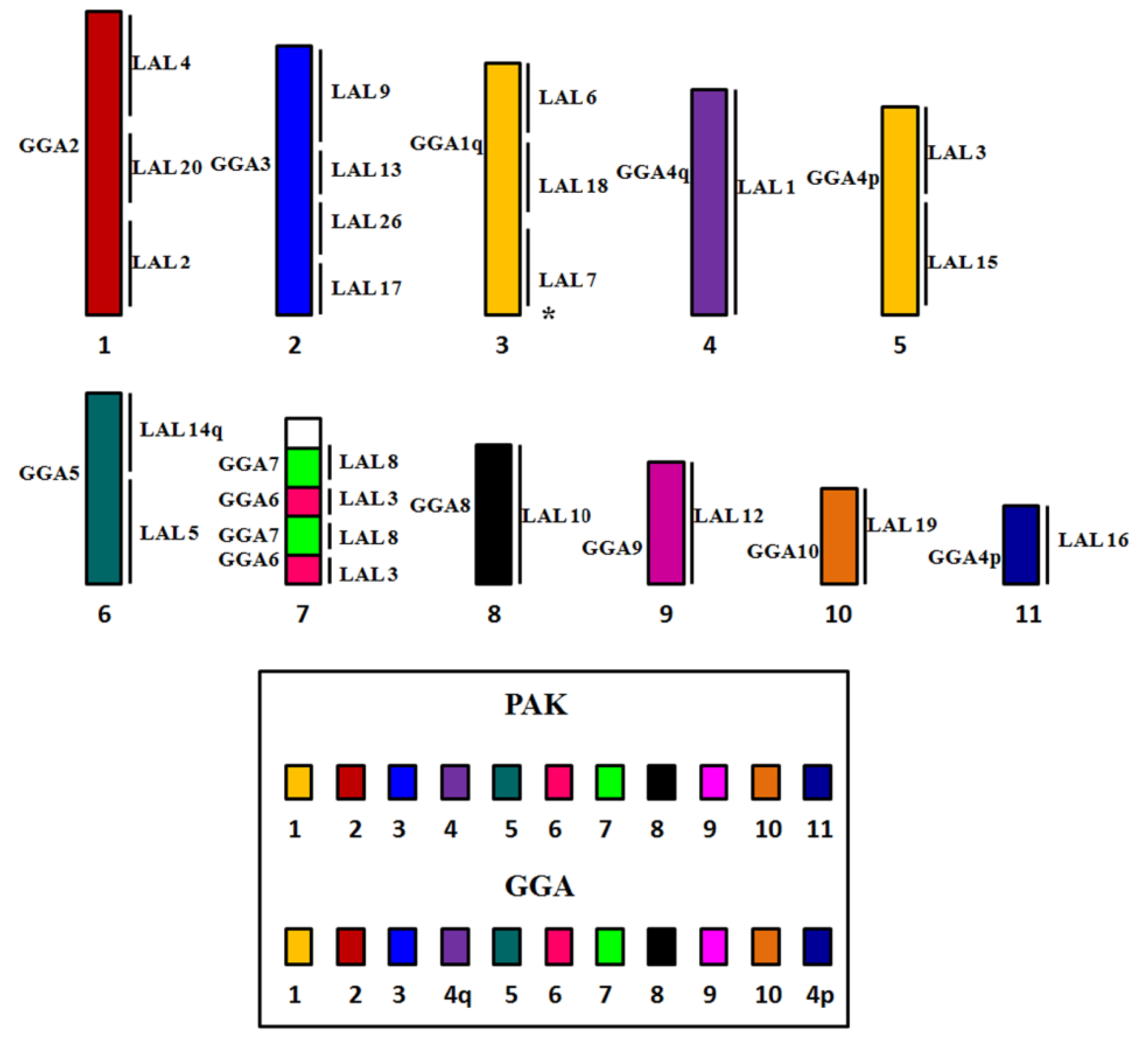
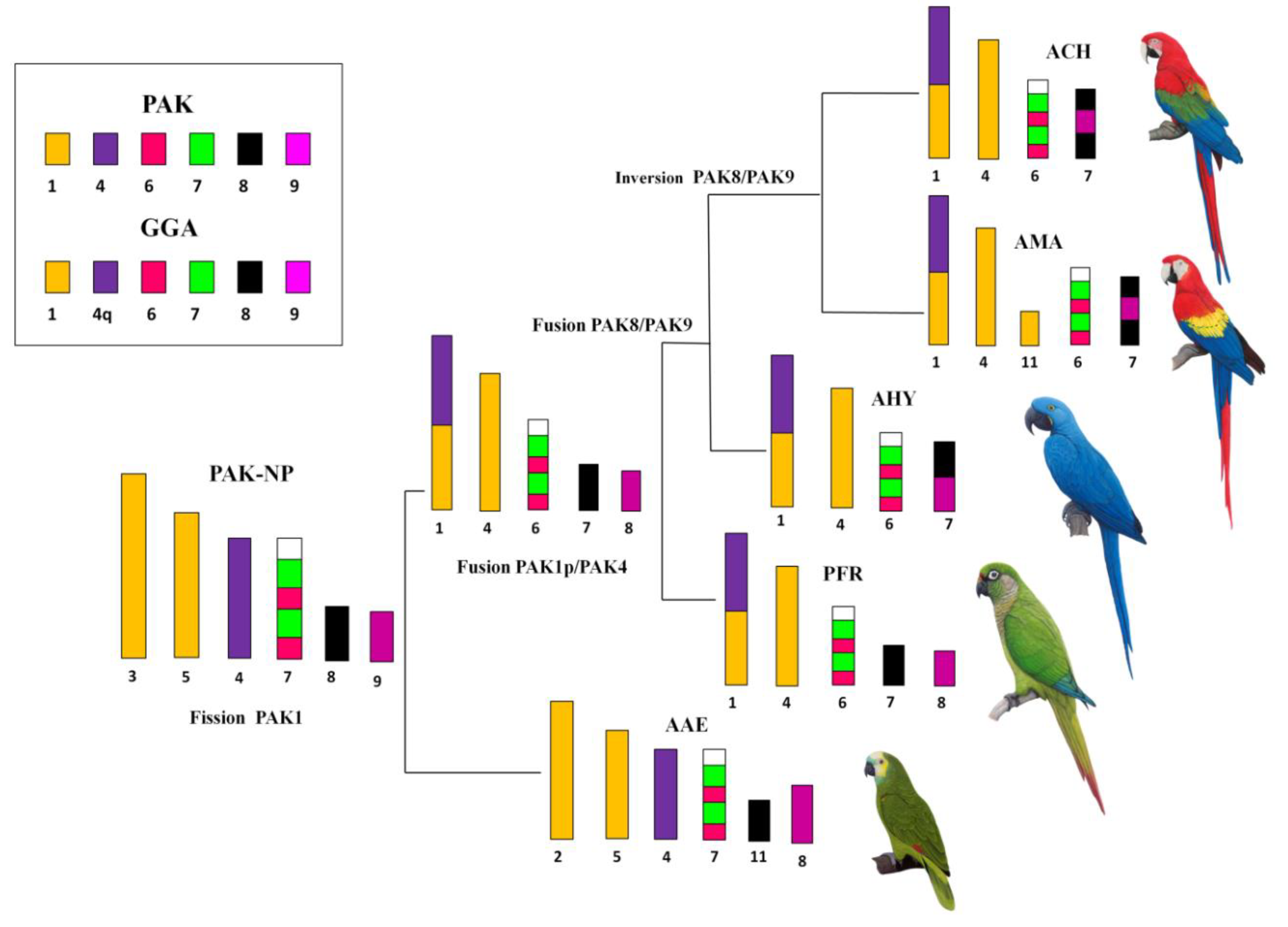
3. Results
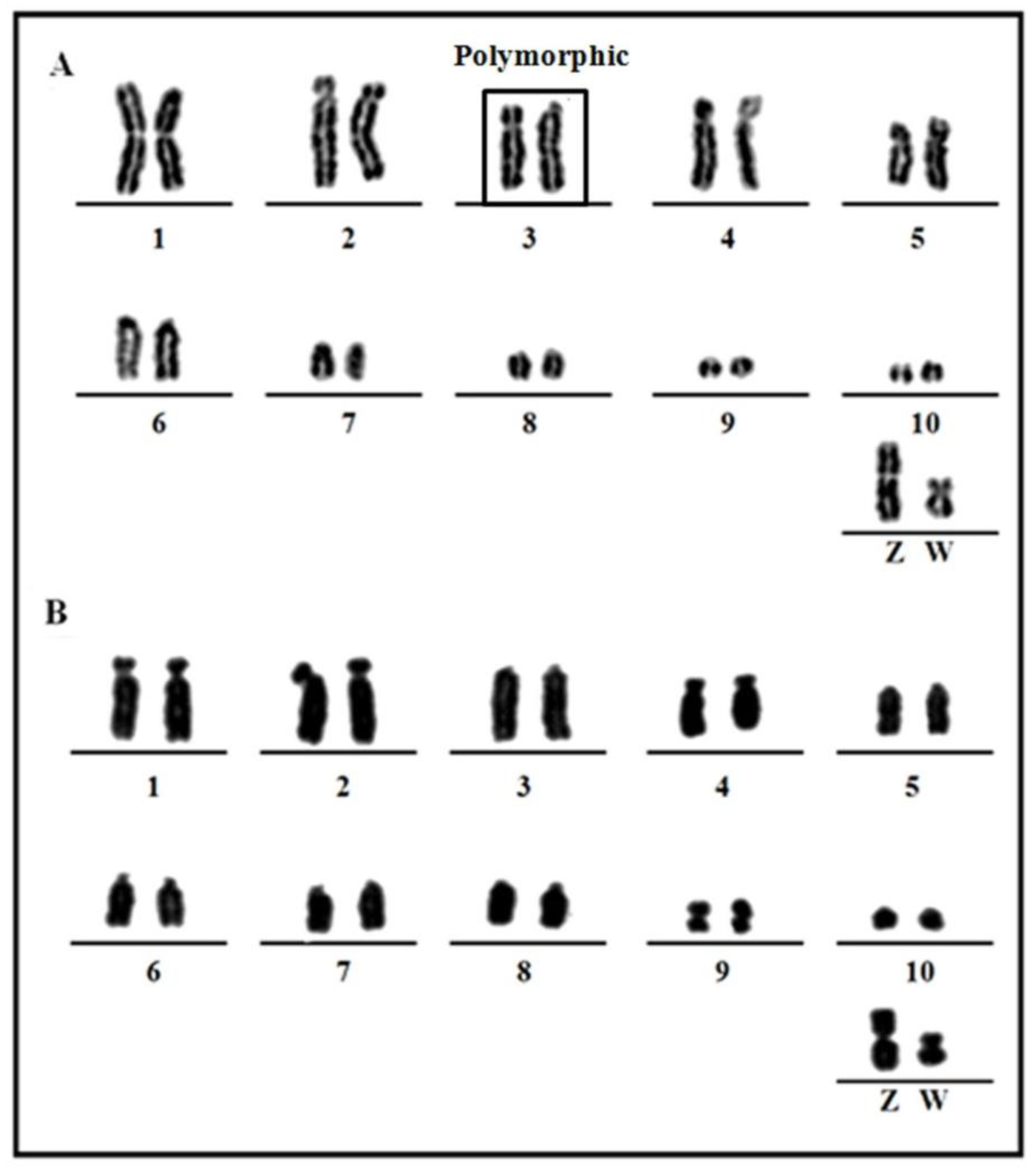
3.1. Karyotype Analysis
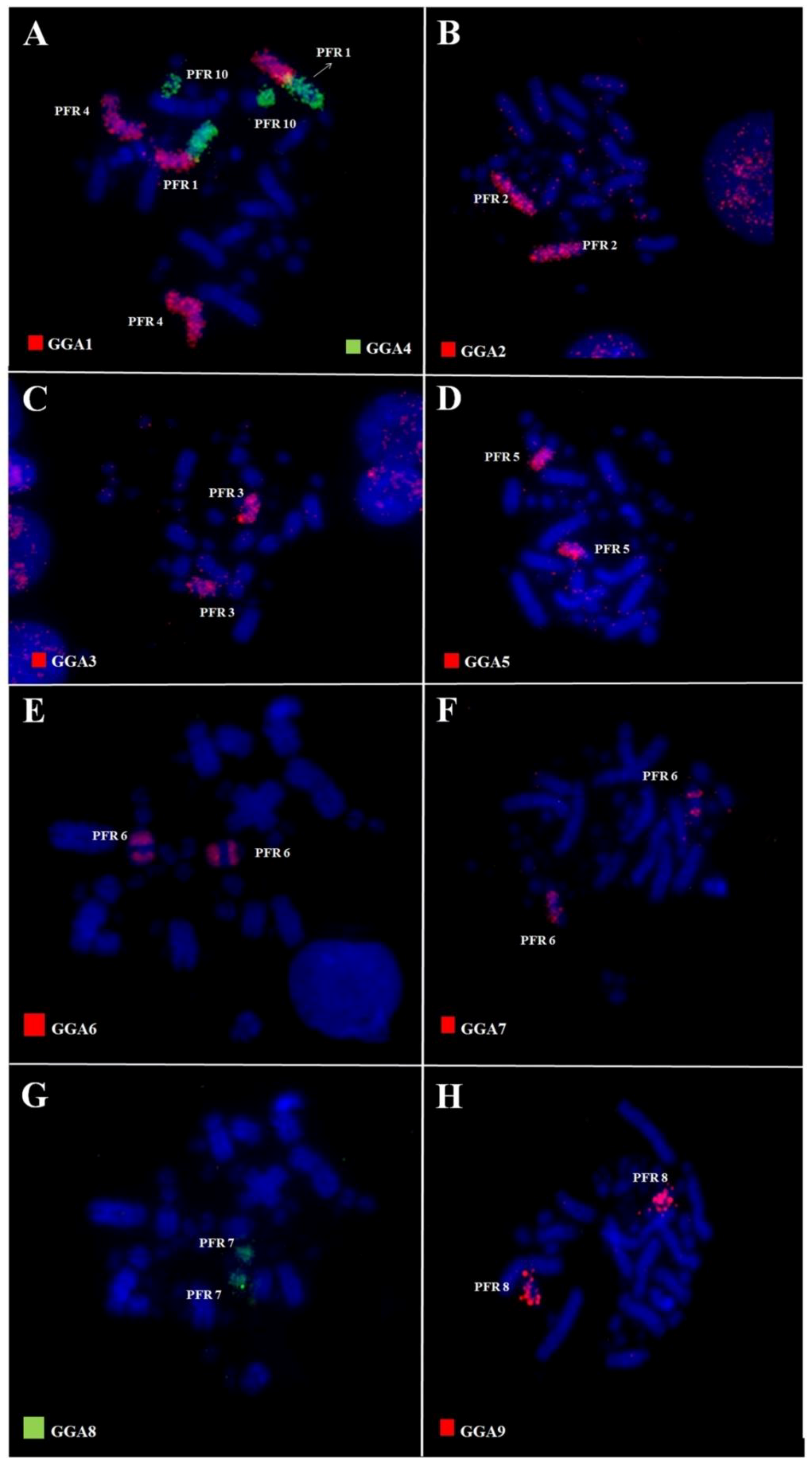
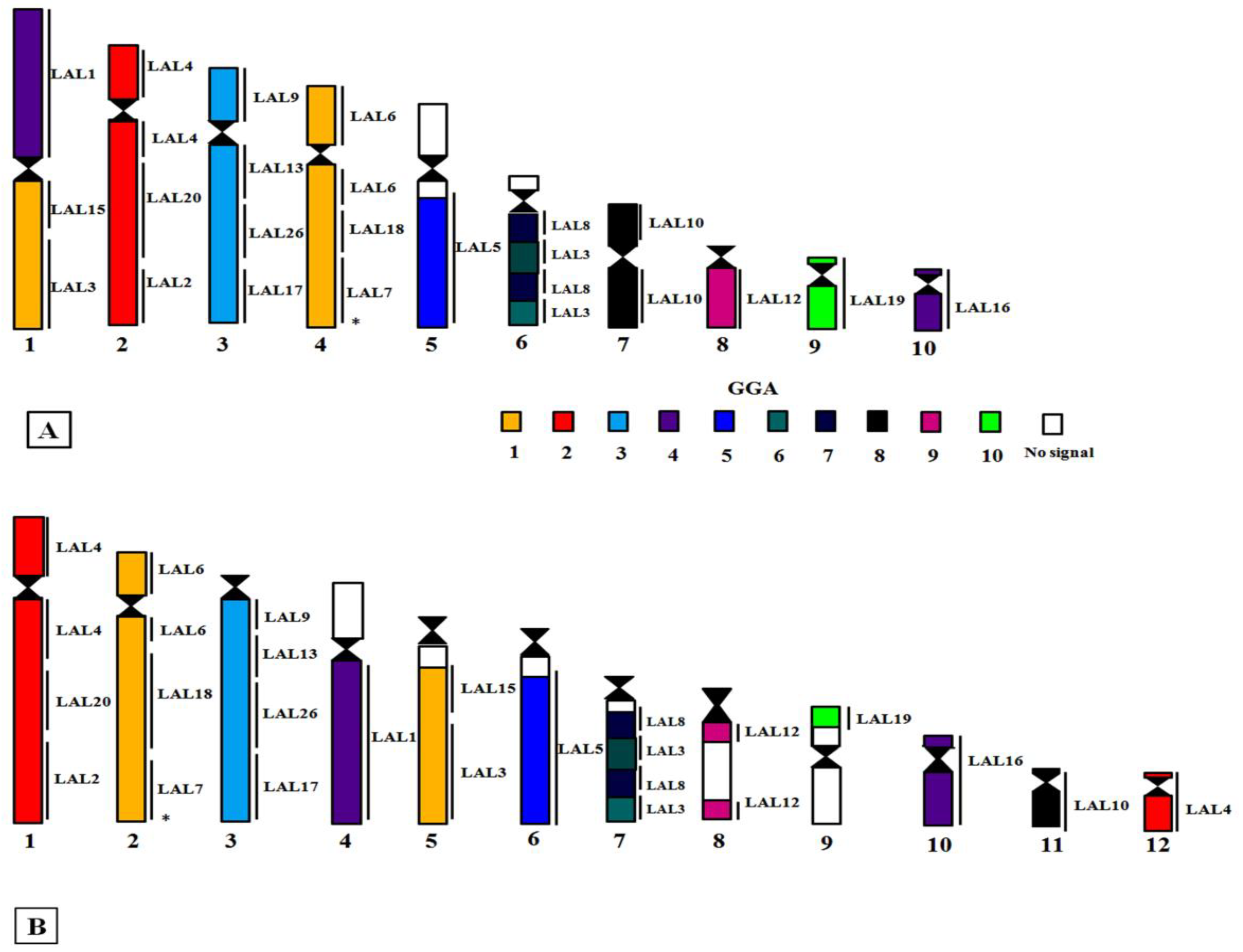
3.2. Comparative Chromosome Painting
4. Discussion
Phylogenetic Analysis
Supplementary Material
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Forshaw, J.W. Parrots of the World, 3rd ed.; Landsdowne Editions: Willoughby, Australia, 1989; p. 336. ISBN 9780691142852. [Google Scholar]
- Collar, N.J. Family Psittacidae (parrots). In Handbook of the Birds of the World; del Hoyo, J., Elliot, A., Sargatal, J., Eds.; Lynx Editions: Barcelona, Spain, 1997; pp. 280–477. [Google Scholar]
- Rowley, I. Family Cacatuidae (Cockatoos). In Handbook of the Birds of the World; del Hoyo, J., Elliot, A., Sargatal, J., Eds.; Lynx Editions: Barcelona, Spain, 1997; pp. 246–279. [Google Scholar]
- Wright, T.F.; Schirtzinger, E.E.; Matsumoto, T.; Eberhard, J.R.; Graves, G.R.; Sanchez, J.J.; Capelli, S.; Müller, H.; Scharpegge, J.; Chambers, G.K.; et al. A multilocus molecular phylogeny of the parrots (Psittaciformes): Support for a Gondwanan Origin during the Cretaceous. Mol. Biol. Evol. 2008, 25, 2141–2156. [Google Scholar] [CrossRef] [PubMed]
- Tavares, E.S.; Baker, A.J.; Pereira, S.L.; Miyaki, C.Y. Phylogenetic relationships and historical biogeography of Neotropical parrots (Psittaciformes: Psittacidae: Arini) inferred from mitochondrial and nuclear DNA sequences. Syst. Biol. 2006, 55, 454–470. [Google Scholar] [CrossRef] [PubMed]
- Miyaki, C.Y.; Matioli, S.R.; Burke, T.; Wajntal, A. Parrot evolution and paleogeographical events: Mitochondrial DNA evidence. Mol. Biol. Evol. 1998, 15, 544–551. [Google Scholar] [CrossRef]
- Tavares, E.S.; Yamashita, C.; Miyaki, C.Y. Phylogenetic relationships among some Neotropical parrot genera (Psittacidae) based on mitochondrial sequences. Auk 2004, 121, 230–242. [Google Scholar] [CrossRef]
- Santos, L.P.; Gunski, R.J. Revisão de dados citogenéticos sobre a avifauna brasileira. Rev. Bras. Ornitol. 2006, 14, 35–45. [Google Scholar]
- Francisco, M.R.; Galetti, J.P.M. Cytotaxonomic considerations on Neotropical Psittacidae birds and description of three new Karyotypes. Hereditas 2001, 134, 225–228. [Google Scholar] [CrossRef] [PubMed]
- Seabury, C.M.; Dowd, S.E.; Seabury, P.M.; Raudsepp, T.; Brightsmith, D.J.; Liboriussen, P.; Halley, Y.; Fisher, C.A.; Owens, E.; Viswanathan, G.; et al. A multiplatform draft de novo genome assembly and comparative analysis for the scarlet macaw (Ara macao). PLoS ONE 2013, 8, e62415. [Google Scholar] [CrossRef] [PubMed]
- Furo, I.O.; Kretschmer, R.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; de Oliveira, E.H.C. Chromosomal diversity and karyotype evolution in South American macaws (Psittaciformes, Psittacidae). PLoS ONE 2015, 10, e0130157. [Google Scholar]
- Furo, I.O.; Monte, A.A.; dos Santos, M.S.; Tagliarini, M.M.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; de Oliveira, E.H. Cytotaxonomy of Eurypyga helias (Gruiformes, Eurypygidae): First karyotypic description and phylogenetic proximity with Rynochetidae. PLoS ONE 2015, 10, e0143982. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, E.H.C.; Tagliarini, M.M.; Rissino, J.D.; Pieczarka, J.C.; Nagamachi, C.Y.; O’Brien, P.C.M.; Ferguson-Smith, M.A. Reciprocal chromosome painting between white hawk (Leucopternis albicollis) and chicken reveals extensive fusions and fissions during karyotype evolution of Accipitridae (Aves, Falconiformes). Chromosome Res. 2010, 18, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Kretschmer, R.; Gunski, R.J.; Garnero, A.D.V.; Furo, I.O.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; de Oliveira, E.H.C. Molecular cytogenetic characterization of multiple intrachromosomal rearrangements in two representatives of the genus Turdus (Turdidae, Passeriformes). PLoS ONE 2014, 9, e103338. [Google Scholar] [CrossRef] [PubMed]
- Kretschmer, R.; de Oliveira, E.H.C.; dos Santos, M.S.; Furo, I.O.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; Garnero, A.D.V.; Gunski, R.J. Chromosome mapping of the large elaenia (Elaenia spectabilis): Evidence for a cytogenetic signature for passeriform birds? Biol. J. Linn. Soc. 2015, 115, 391–398. [Google Scholar] [CrossRef]
- Kretschmer, R.; Furo, I.O.; Gunski, R.J.; Garnero, A.D.V.; Pereira, J.C.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; de Oliveira, E.H.C.; de Freitas, T.R.O. Comparative chromosome painting in Columbidae (Columbiformes) reinforces divergence in Passerea and Columbea. Chromosome Res. 2018, 26, 211–223. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, E.H.C.; Tagliarini, M.M.; dos Santos, M.S.; O’Brien, P.C.M.; Ferguson-Smith, M.A. Chromosome Painting in Three Species of Buteoninae: A Cytogenetic signature reinforces the monophyly of South American species. PLoS ONE 2013, 8, e70071. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, M.; Ikeuchi, T.; Maino, S. A feather pulp culture for avian chromosomes with notes on the chromosomes of the peafowl and the ostrich. Experientia 1968, 24, 1923–1929. [Google Scholar] [CrossRef]
- Griffin, D.K.; Robertson, L.B.; Tempest, H.G.; Skinner, B.M. The evolution of the avian genome as revealed by comparative molecular cytogenetics. Cytogenet. Genome Res. 2007, 117, 64–77. [Google Scholar] [CrossRef] [PubMed]
- Dobigny, G.; Ducroz, J.F.; Robinson, T.J.; Volobouev, V. Cytogenetics and cladistics. Syst. Biol. 2004, 53, 470–484. [Google Scholar] [CrossRef] [PubMed]
- Aquino, R.; Ferrari, I. Chromosome study of Amazona amazonica and A. aestiva (Aves: Psittaciformes) determination of chromosome number and identification of sex chromosomes by C-banding methods. Genetica 1990, 81, 1–3. [Google Scholar]
- Duarte, J.M.B.; Caparroz, R. Cytotaxonomic analysis of Brazilian species of the genus Amazona (Psittacidae, Aves) and confirmation of the genus Salvatoria (Ribeiro, 1920). Braz. J. Genet. 1995, 18, 623–628. [Google Scholar]
- Seibold-Torres, C.; Owens, E.; Chowdhary, R.; Ferguson-Smith, M.A.; Tizard, I.; Raudsepp, T. Comparative cytogenetics of the Congo African grey parrot (Psittacus erithacus). Cytogenet. Genome Res. 2016, 147, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Guttenbach, M.; Nanda, I.; Feichtinger, W.; Masabanda, J.S.; Griffin, D.K.; Schmid, M. Comparative chromosome painting of chicken autosomal paints 1–9 in nine different bird species. Cytogenet. Genome Res. 2003, 103, 173–184. [Google Scholar] [CrossRef] [PubMed]
- Nanda, I.; Karl, E.; Volobouev, V.; Griffin, D.K.; Scharlt, M.; Schmid, M. Extensive gross genomic rearrangements between chicken and old world vultures (Falconiformes, Accipitridae). Cytogenet. Genome Res. 2006, 112, 286–295. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, E.H.; de Moura, S.P.; dos Anjos, L.J.; Nagamachi, C.Y.; Pieczarka, J.C.; O’Brien, P.C.M.; Ferguson-Smith, M.A. Comparative chromosome painting between chicken and spectacled owl (Pulsatrix perspicillata): Implications for chromosomal evolution in the Strigidae (Aves, Strigiformes). Cytogenet. Genome Res. 2008, 122, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Nanda, I.; Karl, E.; Griffin, D.K.; Schartl, M.; Schmid, M. Chromosome repatterning in three representative parrots (Psittaciformes) inferred from comparative chromosome painting. Cytogenet. Genome Res. 2007, 117, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Kretschmer, R.; Ferguson-Smith, M.A.; de Oliveira, E.H.C. Karyotype Evolution in birds: From conventional staining to chromosome painting. Genes 2018, 9, 181. [Google Scholar] [CrossRef] [PubMed]
- Sick, H. Ornitologia Brasileira, 2nd ed.; Editora Nova Fronteira: Rio de Janeiro, Brazil, 1997; p. 862. ISBN 8520908160. [Google Scholar]
- Miyaki, C.Y.; Duarte, J.M.B.; Caparroz, R.; Nunes, A.L.V.; Wajntal, A. Sex identification of New World parrots (Psittacidae, Aves) using the human minisatellite probe 33.15. Auk 1997, 114, 516–520. [Google Scholar] [CrossRef]
- Montón, M. Algunos aspectos significativos en la adaptación de los psitácidos de Sudamérica. Revista del Parque Zoológico de Barcelona 1977, 30, 19–22. [Google Scholar]
- De Lucca, E.J.; de Marco, D.A. Chromosomal Polymorphism in Forpus Xanthopterygius (Psittaciformes: Aves). Caryologia 1983, 36, 355–361. [Google Scholar] [CrossRef]
- Lunardi, V.O.; Francisco, M.R.; Rocha, G.T.; Goldschmidt, B.; Galetti, J.P.M. Karyotype description of two Neotropical Psittacidae species: The endangered Hyacinth macaw, Anodorhynchus hyacinthinus, and the Hawk-headed Parrot, Deroptyus accipitrinus (Psittaciformes: Aves), and its significance for conservation plans. Genet. Mol. Biol. 2003, 26, 283–287. [Google Scholar] [CrossRef]
- De Lucca, E.J.; Shirley, L.R.; Lanier, C. Karyotype studies in twenty-two species of parrots (Psittaciformes: Aves). Genet. Mol. Biol. 1991, 14, 73–98. [Google Scholar]
- Barrowclough, G.F.; Groth, J.G.; Mertz, L.A. Phylogenetic relationships among parrots. In Proceedings of the One Hundred and Twenty-Second Stated Meeting of the American Ornithologists’ Union, Quebec City, QC, Canada, 16–21 August 2004. [Google Scholar]
- De Kloet, R.S.; de Kloet, S.R. The evolution of the spindlin gene in birds: Sequence analysis of anintron of the spindlin W and Z gene revealed four major divisions of the Psittaciformes. Mol. Phylogenet. Evol. 2005, 36, 706–721. [Google Scholar] [CrossRef] [PubMed]








© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Furo, I.D.O.; Kretschmer, R.; O’Brien, P.C.M.; Pereira, J.C.; Garnero, A.D.V.; Gunski, R.J.; Ferguson-Smith, M.A.; De Oliveira, E.H.C. Chromosome Painting in Neotropical Long- and Short-Tailed Parrots (Aves, Psittaciformes): Phylogeny and Proposal for a Putative Ancestral Karyotype for Tribe Arini. Genes 2018, 9, 491. https://doi.org/10.3390/genes9100491
Furo IDO, Kretschmer R, O’Brien PCM, Pereira JC, Garnero ADV, Gunski RJ, Ferguson-Smith MA, De Oliveira EHC. Chromosome Painting in Neotropical Long- and Short-Tailed Parrots (Aves, Psittaciformes): Phylogeny and Proposal for a Putative Ancestral Karyotype for Tribe Arini. Genes. 2018; 9(10):491. https://doi.org/10.3390/genes9100491
Chicago/Turabian StyleFuro, Ivanete De Oliveira, Rafael Kretschmer, Patrícia C. M. O’Brien, Jorge C. Pereira, Analía Del Valle Garnero, Ricardo J. Gunski, Malcolm A. Ferguson-Smith, and Edivaldo Herculano Corrêa De Oliveira. 2018. "Chromosome Painting in Neotropical Long- and Short-Tailed Parrots (Aves, Psittaciformes): Phylogeny and Proposal for a Putative Ancestral Karyotype for Tribe Arini" Genes 9, no. 10: 491. https://doi.org/10.3390/genes9100491
APA StyleFuro, I. D. O., Kretschmer, R., O’Brien, P. C. M., Pereira, J. C., Garnero, A. D. V., Gunski, R. J., Ferguson-Smith, M. A., & De Oliveira, E. H. C. (2018). Chromosome Painting in Neotropical Long- and Short-Tailed Parrots (Aves, Psittaciformes): Phylogeny and Proposal for a Putative Ancestral Karyotype for Tribe Arini. Genes, 9(10), 491. https://doi.org/10.3390/genes9100491