mCherry-Labeled Verticillium dahliae Could Be Utilized to Investigate Its Pathogenicity Process in Nicotiana benthamiana

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plants and Fungal Strain
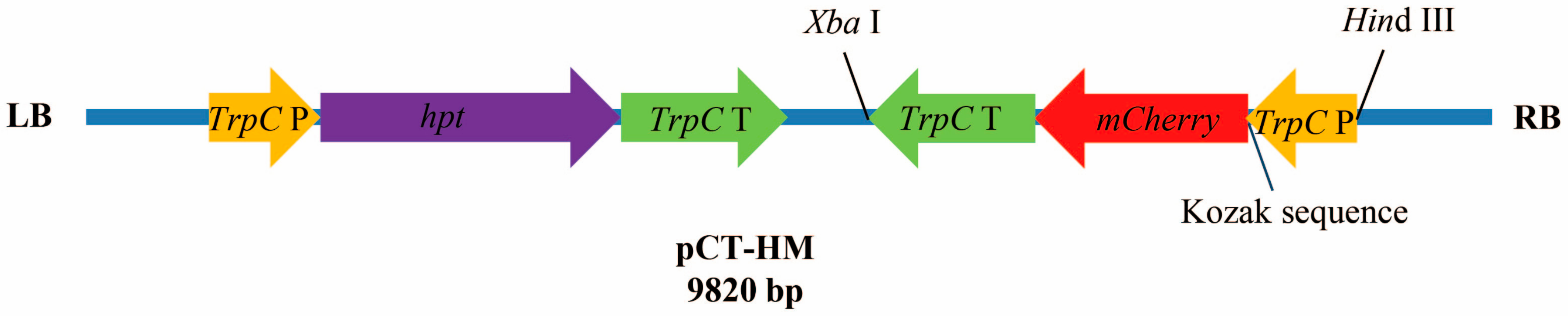
2.2. Codon Optimization of mCherry and Construction of Expression Plasmid
2.3. Fungal Transformation
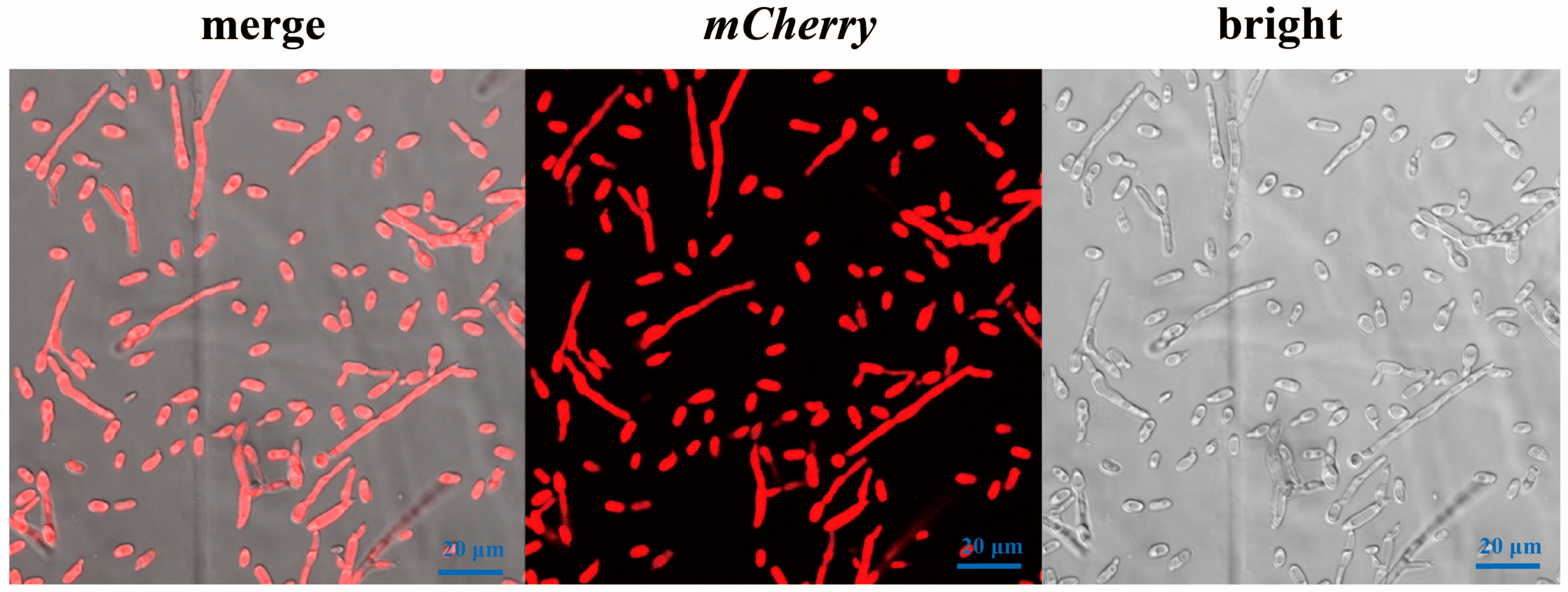
2.4. Screening of mCherry-Labeled Verticillium dahliae (Vd-m)
2.5. Characterization of Vd-m
2.6. Plant Inoculation
2.7. Microscopic Observation of Pathogenic Process
2.8. Quantification of Fungal Biomass
2.9. Statistical Analysis
3. Results
3.1. Optimization of mCherry Gene and Plasmid Construction
3.2. Confirmation of Vd-m Isolates
3.3. Analysis of Biological Characteristics of Vd-m
3.4. Attachment and Colonization of Vd-m on Roots
3.5. Advanced Penetration Stage of Vd-m in Roots
3.6. Vd-m Extensively Colonized Root Tissues
3.7. Microscopic Examination of Leaves
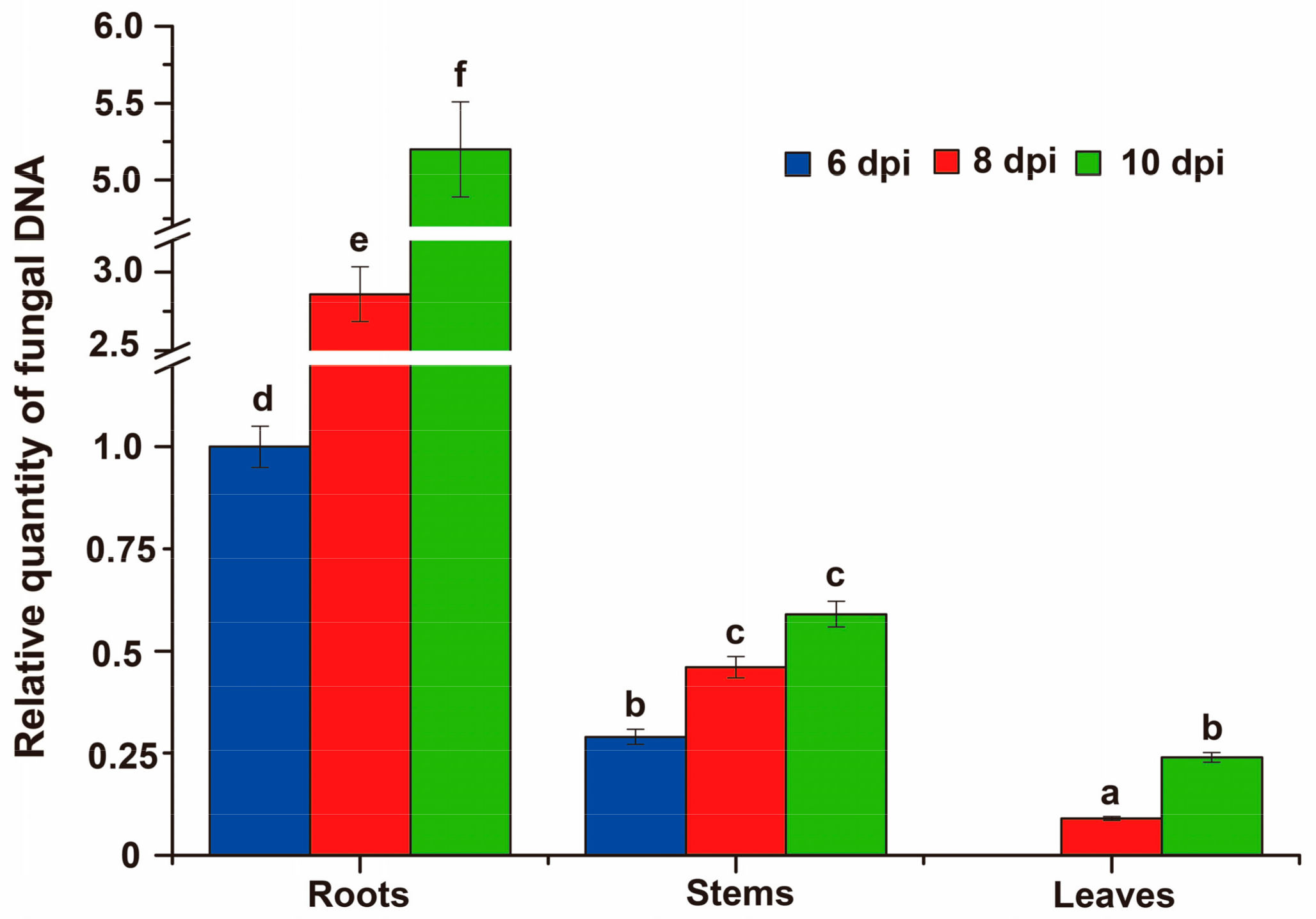
3.8. Fungal Biomass at Different Stages
4. Discussion
5. Conclusion
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Pegg, G.F.; Brady, B.L. Verticillium Wilts; CABI Publishing: Oxford, UK, 2002; pp. 142–166. [Google Scholar]
- Hu, D.; Wang, C.; Tao, F.; Cui, Q.; Xu, X.; Shang, W.; Hu, X. Whole genome wide expression profiles on germination of Verticillium dahliae microsclerotia. PLoS ONE 2014, 9, e100046. [Google Scholar] [CrossRef] [PubMed]
- Farley, J.D.; Wilhelm, S.; Snyder, W.C. Repeated germination and sporulation of microsclerotia of Verticillium albo-atrum in soil. Phytopathology 1971, 61, 260–264. [Google Scholar] [CrossRef]
- Duressa, D.; Rauscher, G.; Koike, S.T.; Mou, B.; Hayes, R.J.; Maruthachalam, K.; Subbarao, K.V.; Klosterman, S.J. A real-time PCR assay for detection and quantification of Verticillium dahliae in spinach seed. Phytopathology 2012, 102, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Vallad, G.E.; Bhat, R.G.; Koike, S.T.; Ryder, E.J.; Subbarao, K.V. Weedborne reservoirs and seed transmission of Verticillium dahliae in lettuce. Plant Dis. 2007, 89, 317–324. [Google Scholar] [CrossRef]
- Bubici, G.; Cirulli, M. Verticillium wilt of olive: Current status, disease management and future prospects. Prot. Delle Colt. 2012, 4, 42–56. [Google Scholar]
- Rowe, R.C.; Davis, J.R.; Powelson, M.L.; Rouse, D.I. Potato early dying: Causal agents and management strategies. Plant Dis. 1987, 71, 482–489. [Google Scholar] [CrossRef]
- López-Escudero, F.J.; Mercado-Blanco, J. Verticillium wilt of olive: A case study to implement an integrated strategy to control a soil-borne pathogen. Plant Soil 2010, 344, 1–50. [Google Scholar] [CrossRef] [Green Version]
- Rehman, L.; Su, X.; Li, X.; Qi, X.; Guo, H.; Cheng, H. FreB is involved in the ferric metabolism and multiple pathogenicity-related traits of Verticillium dahliae. Curr. Genet. 2018, 64, 645–659. [Google Scholar] [CrossRef] [PubMed]
- Mol, L.; Riessen, H.W.V. Effect of plant roots on the germination of microsclerotia of Verticillum dahliae. Eur. J. Plant Pathol. 1995, 101, 673–678. [Google Scholar] [CrossRef]
- Fradin, E.F.; Thomma, B.P. Physiology and molecular aspects of Verticillium wilt diseases caused by V. dahliae and V. albo-atrum. Mol. Plant Pathol. 2006, 7, 71–86. [Google Scholar] [CrossRef] [PubMed]
- Vallad, G.E.; Subbarao, K.V. Colonization of resistant and susceptible lettuce cultivars by a green fluorescent protein-tagged isolate of Verticillium dahliae. Phytopathology 2008, 98, 871–885. [Google Scholar] [CrossRef] [PubMed]
- Iakovoss, P.; Sotiriose, T.; Ioannisa, S.; Iordanis, C.; Epaminondasj, P. Mode of action of a non-pathogenic Fusarium oxysporum strain against Verticillium dahliae using Real Time QPCR analysis and biomarker transformation. Biol. Control 2009, 50, 30–36. [Google Scholar] [CrossRef]
- Huisman, O. Interrelations of root growth dynamics to epidemiology of root-invading fungi. Annu. Rev. Phytopathol. 1982, 20, 303–327. [Google Scholar] [CrossRef]
- Evans, G.; Gleeson, A.C.; Evans, G.; Gleeson, A.C. Observations on the origin and nature of Verticillium dahliae colonizing plant roots. Aust. J. Biol. Sci. 1973, 26, 151–162. [Google Scholar] [CrossRef]
- Zhao, P.; Zhao, Y.L.; Jin, Y.; Zhang, T.; Guo, H.S. Colonization process of Arabidopsis thaliana roots by a green fluorescent protein-tagged isolate of Verticillium dahliae. Protein Cell 2014, 5, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, J.; Gao, J.; Zhang, G.; Yu, Y.; Zhou, H.; Chen, W.; Zhao, J. The colonization process of sunflower by a green fluorescent protein-tagged isolate of Verticillium dahliae and its seed transmission. Plant Dis. 2018, 102, 1772–1778. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xiao, S.; Xiong, D.; Tian, C. Genetic transformation, infection process and qPCR quantification of Verticillium dahliae on smoke-tree Cotinus coggygria. Australas. Plant Pathol. 2012, 42, 33–41. [Google Scholar] [CrossRef]
- Zhang, W.; Jiang, T.; Cui, X.; Qi, F.; Jian, G. Colonization in cotton plants by a green fluorescent protein labelled strain of Verticillium dahliae. Eur. J. Plant Pathol. 2013, 135, 867–876. [Google Scholar] [CrossRef]
- Kendall, J.; Badminton, M. Aequorea victoria bioluminescence moves into an exciting new era. Trends Biotechnol. 1998, 16, 216–224. [Google Scholar] [CrossRef]
- Meyer, A.J.; Dick, T.P. Fluorescent protein-based redox probes. Antioxid. Redox Signal. 2010, 13, 621–650. [Google Scholar] [CrossRef] [PubMed]
- Lubeck, M.; Imb, K.; Jensen, B.; Thrane, U.; Janvier, C.; Jensen, D.F. GUS and GFP transformation of the biocontrol strain Clonostachys rosea IK726 and the use of these marker genes in ecological studies. Mycol. Res. 2002, 106, 815–826. [Google Scholar] [CrossRef]
- Weid, I.V.D.; Artursson, V.; Seldin, L.; Jansson, J.K. Antifungal and root surface colonization properties of GFP-tagged Paenibacillus brasilensis PB177. World J. Microbiol. Biotechnol. 2005, 21, 1591–1597. [Google Scholar] [CrossRef]
- Mansouri, S. Developing Novel Molecular Tools to Study the Fusarium virguliforme-Soybean Interaction. Master’s Thesis, Southern Illinois University, Carbondale, IL, USA, 2007. [Google Scholar]
- Huh, W.K.; Falvo, J.V.; Gerke, L.C.; Carroll, A.S.; Howson, R.W.; Weissman, J.S.; O’Shea, E.K. Global analysis of protein localization in budding yeast. Nature 2003, 425, 686–691. [Google Scholar] [CrossRef] [PubMed]
- Skadsen, R.W.; Hohn, T.M. Use of Fusarium graminearum transformed with GFP to follow infection patterns in barley and Arabidopsis. Physiol. Mol. Plant Pathol. 2004, 64, 45–53. [Google Scholar] [CrossRef]
- Mcdougal, R.; Yang, S.; Schwelm, A.; Stewart, A.; Bradshaw, R. A novel GFP-based approach for screening biocontrol microorganisms in vitro against Dothistroma septosporum. J. Microbiol. Methods 2011, 87, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Li, E.; Wang, G.; Yang, Y.; Xiao, J.; Mao, Z.; Xie, B. Microscopic analysis of the compatible and incompatible interactions between Fusarium oxysporum f. sp. conglutinans and cabbage. Eur. J. Plant Pathol. 2014, 141, 597–609. [Google Scholar] [CrossRef]
- Su, X.; Rehman, L.; Guo, H.; Li, X.; Cheng, H. The oligosaccharyl transferase subunit STT3 mediates fungal development and is required for virulence in Verticillium dahliae. Curr. Genet. 2018, 64, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Sarrocco, S.; Falaschi, N.; Vergara, M.; Nicoletti, F.; Vannacci, G. Use of Fusarium oxysporum f. sp. dianthi transformed with marker genes to follow colonization of carnation roots. J. Plant Pathol. 2007, 89, 47–54. [Google Scholar] [CrossRef]
- Monteiro, R.A.; Schmidt, M.A.; Baura, V.A.; Balsanelli, E.; Wassem, R.; Yates, M.G.; Randi, M.A.; Pedrosa, F.O.; Souza, E.M. Early colonization pattern of maize (Zea mays L. Poales, Poaceae) roots by Herbaspirillum seropedicae (Burkholderiales, Oxalobacteraceae). Genet. Mol. Biol. 2008, 31, 932–937. [Google Scholar] [CrossRef]
- Ilgen, P.; Hadeler, B.; Maier, F.J.; Schäfer, W. Developing kernel and rachis node induce the trichothecene pathway of Fusarium graminearum during wheat head infection. Mol. Plant Microbe Interact. 2009, 22, 899–908. [Google Scholar] [CrossRef] [PubMed]
- Shaner, N.C.; Campbell, R.E.; Steinbach, P.A.; Giepmans, B.N.; Palmer, A.E.; Tsien, R.Y. Improved monomeric red, orange and yellow fluorescent proteins derived from Discosoma sp. red fluorescent protein. Nat. Biotechnol. 2004, 22, 1567–1572. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Qi, H.; Zhang, J.; Yangqi, G.; Tang, C. Transformation and selection of fluorescent protein gene labelled Verticillium dahliae. Cotton Sci. 2014, 26, 221–227. [Google Scholar]
- Shrivastava, S.; Poddar, R.; Shukla, P.; Mukhopadhyay, K. Study of codon bias perspective of fungal xylanase gene by multivariate analysis. Bioinformation 2009, 3, 425–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehman, L.; Su, X.; Guo, H.; Qi, X.; Cheng, H. Protoplast transformation as a potential platform for exploring gene function in Verticillium dahliae. BMC Biotechnol. 2016, 16, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Tzima, A.K.; Paplomatas, E.J.; Tsitsigiannis, D.I.; Kang, S. The G protein β subunit controls virulence and multiple growth- and development-related traits in Verticillium dahliae. Fungal Genet. Biol. 2012, 49, 271–283. [Google Scholar] [CrossRef] [PubMed]
- Lorang, J.M.; Tuori, R.P.; Martinez, J.P.; Sawyer, T.L.; Redman, R.S.; Rollins, J.A.; Wolpert, T.J.; Johnson, K.B.; Rodriguez, R.J.; Dickman, M.B. Green fluorescent protein is lighting up fungal biology. Appl. Environ. Microbiol. 2001, 67, 1987–1994. [Google Scholar] [CrossRef] [PubMed]
- Turgeon, B.G.; Garber, R.C.; Yoder, C.O. Development of a fungal transformation system based on selection of sequences with promoter activity. Mol. Cell. Biol. 1987, 7, 3297–3305. [Google Scholar] [CrossRef] [PubMed]
- Okuda, T.; Ando, A.; Sakuradani, E.; Kikukawa, H.; Kamada, N.; Ochiai, M.; Shima, J.; Ogawa, J. Selection and characterization of promoters based on genomic approach for the molecular breeding of oleaginous fungus Mortierella alpina 1S-4. Curr. Genet. 2014, 60, 183–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arazoe, T.; Miyoshi, K.; Yamato, T.; Ogawa, T.; Ohsato, S.; Arie, T.; Kuwata, S. Tailor-made CRISPR/Cas system for highly efficient targeted gene replacement in the rice blast fungus. Biotechnol. Bioeng. 2015, 112, 2543–2549. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Meng, X.; Wei, X.; Lu, L. Highly efficient CRISPR mutagenesis by microhomology-mediated end joining in Aspergillus fumigatus. Fungal Genet. Biol. 2016, 86, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wang, Z.; Ni, H.; Xu, Y.; Chen, Q.; Jiang, L. CRISPR/Cas9-mediated base-editing system efficiently generates gain-of-function mutations in Arabidopsis. Sci. China Life Sci. 2017, 60, 520–523. [Google Scholar] [CrossRef] [PubMed]
- Takenaka, Y.; Haga, N.; Harumoto, T.; Matsuura, T.; Mitsui, Y. Transformation of Paramecium caudatum with a novel expression vector harboring codon-optimized GFP gene. Gene 2002, 284, 233–240. [Google Scholar] [CrossRef]
- Martinezjaramillo, E.; Garzamorales, R.; Loeraarias, M.J.; Saucedocardenas, O.; Montesdeocaluna, R.; Mcnally, L.R.; Gomezgutierrez, J.G. Development of Lactococcus lactis encoding fluorescent proteins, GFP, mCherry and iRFP regulated by the nisin-controlled gene expression system. Stain Technol. 2017, 92, 167–174. [Google Scholar] [CrossRef]
- Wu, L.; Conner, R.L.; Wang, X.; Xu, R.; Li, H. Variation in growth, colonization of maize, and metabolic parameters of GFP- and DsRed-labeled Fusarium verticillioides strains. Phytopathology 2016, 106, 890–899. [Google Scholar] [CrossRef] [PubMed]
- Duncan, K.E.; Howard, R.J. Biology of maize kernel infection by Fusarium verticillioides. Mol. Plant-Microbe Interact. 2010, 23, 6–16. [Google Scholar] [CrossRef] [PubMed]
- Crespo-Sempere, A.; López-Pérez, M.; Martínez-Culebras, P.V.; González-Candelas, L. Development of a green fluorescent tagged strain of Aspergillus carbonarius to monitor fungal colonization in grapes. Int. J. Food Microbiol. 2011, 148, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, D.A. The development of lignitubers in roots after infection by Verticillium dahliae Kleb. Can. J. Microbiol. 1971, 17, 441–444. [Google Scholar] [CrossRef] [PubMed]
- Gerik, J.S.; Huisman, O.C. Study of field-grown cotton roots infected with Verticillium dahliae using an immunoenzymatic staining technique. Phytopathology 1988, 78, 1174–1178. [Google Scholar] [CrossRef]
- Eynck, C.; Koopmann, B.; Grunewaldt-Stoecker, G.; Karlovsky, P.; von Tiedemann, A. Differential interactions of Verticillium longisporum and V. dahliae with Brassica napus detected with molecular and histological techniques. Eur. J. Plant Pathol. 2007, 118, 259–274. [Google Scholar] [CrossRef]
- Reusche, M.; Truskina, J.; Thole, K.; Nagel, L.; Rindfleisch, S.; Tran, V.T.; Braus-Stromeyer, S.A.; Braus, G.H.; Teichmann, T.; Lipka, V. Infections with the vascular pathogens Verticillium longisporum and Verticillium dahliae induce distinct disease symptoms and differentially affect drought stress tolerance of Arabidopsis thaliana. Environ. Exp. Bot. 2014, 108, 23–37. [Google Scholar] [CrossRef]
- Zhao, Y.L.; Zhou, T.T.; Guo, H.S. Hyphopodium-epecific VdNoxB/VdPls1-dependent ROS-Ca2+ signaling is required for plant infection by Verticillium dahliae. PLoS Pathog. 2016, 12, e1005793. [Google Scholar] [CrossRef] [PubMed]
- Ralhan, A.; Schottle, S.; Thurow, C.; Iven, T.; Feussner, I.; Polle, A.; Gatz, C. The vascular pathogen Verticillium longisporum requires a jasmonic acid-independent COI1 function in roots to elicit disease symptoms in Arabidopsis shoots. Plant Physiol. 2012, 159, 1192–1203. [Google Scholar] [CrossRef] [PubMed]
- Dimond, A.E. Biophysics and biochemistry of the vascular wilt syndrome. Annu. Rev. Phytopathol. 1970, 8, 301–322. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Sequence (5′-3′) |
|---|---|
| Ptrpc | atcAAGCTTTTGAAGGAGCATTTTTGGGCTTGGC |
| CTCGCCCTTGGAGACCATGGTGGCATCGATGCTTGGGTAG | |
| Pmch | ATGGTCTCCAAGGGCGAGGAGGACAAC |
| CTACTTGTAGAGCTCGTCCATGCCGCC | |
| Ttrpc | CATGGACGAGCTCTACAAGTAGAGTAGATGCCGACCGGGATCC |
| aacTCTAGATTATCTTTGCGAACCCAGGGGCTG | |
| qRT-VdITS | CCGCCGGTCCATCAGTCTCTCTGTTTATAC |
| CGCCTGCGGGACTCCGATGCGAGCTGTAAC | |
| qRT-Nbactin | GGACCTTTATGGAAACATTGTGCTCAGT |
| CCAAGATAGAACCTCCAATCCAGACAC | |
| Det-hpt | GAGGGCGAAGAATCTCGTGCTTTCA |
| TGTTATGCGGCCATTGTCCGTCAGG | |
| Det-mch | CTACGTTAAGCACCCCGCCGACATT |
| CTGCTCGACAATCGTGTAGTCCTCGT |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, X.; Lu, G.; Rehman, L.; Li, X.; Sun, L.; Guo, H.; Cheng, H. mCherry-Labeled Verticillium dahliae Could Be Utilized to Investigate Its Pathogenicity Process in Nicotiana benthamiana. Genes 2018, 9, 508. https://doi.org/10.3390/genes9100508
Su X, Lu G, Rehman L, Li X, Sun L, Guo H, Cheng H. mCherry-Labeled Verticillium dahliae Could Be Utilized to Investigate Its Pathogenicity Process in Nicotiana benthamiana. Genes. 2018; 9(10):508. https://doi.org/10.3390/genes9100508
Chicago/Turabian StyleSu, Xiaofeng, Guoqing Lu, Latifur Rehman, Xiaokang Li, Lu Sun, Huiming Guo, and Hongmei Cheng. 2018. "mCherry-Labeled Verticillium dahliae Could Be Utilized to Investigate Its Pathogenicity Process in Nicotiana benthamiana" Genes 9, no. 10: 508. https://doi.org/10.3390/genes9100508
APA StyleSu, X., Lu, G., Rehman, L., Li, X., Sun, L., Guo, H., & Cheng, H. (2018). mCherry-Labeled Verticillium dahliae Could Be Utilized to Investigate Its Pathogenicity Process in Nicotiana benthamiana. Genes, 9(10), 508. https://doi.org/10.3390/genes9100508