Generation of A Mucor circinelloides Reporter Strain—A Promising New Tool to Study Antifungal Drug Efficacy and Mucormycosis

 ,
,  , ,
, , 
 and
and 
Abstract
:1. Introduction
2. Material and Methods
2.1. Fungal Strains, Plasmids, Media and Growth Conditions
2.2. Cloning Procedures
2.2.1. Amplification of the Firefly Luciferase Gene
2.2.2. Plasmid Construction
2.3. Transformation of Mucor circinelloides and Initial Screening
2.4. Genomic DNA Extraction and Southern Analysis
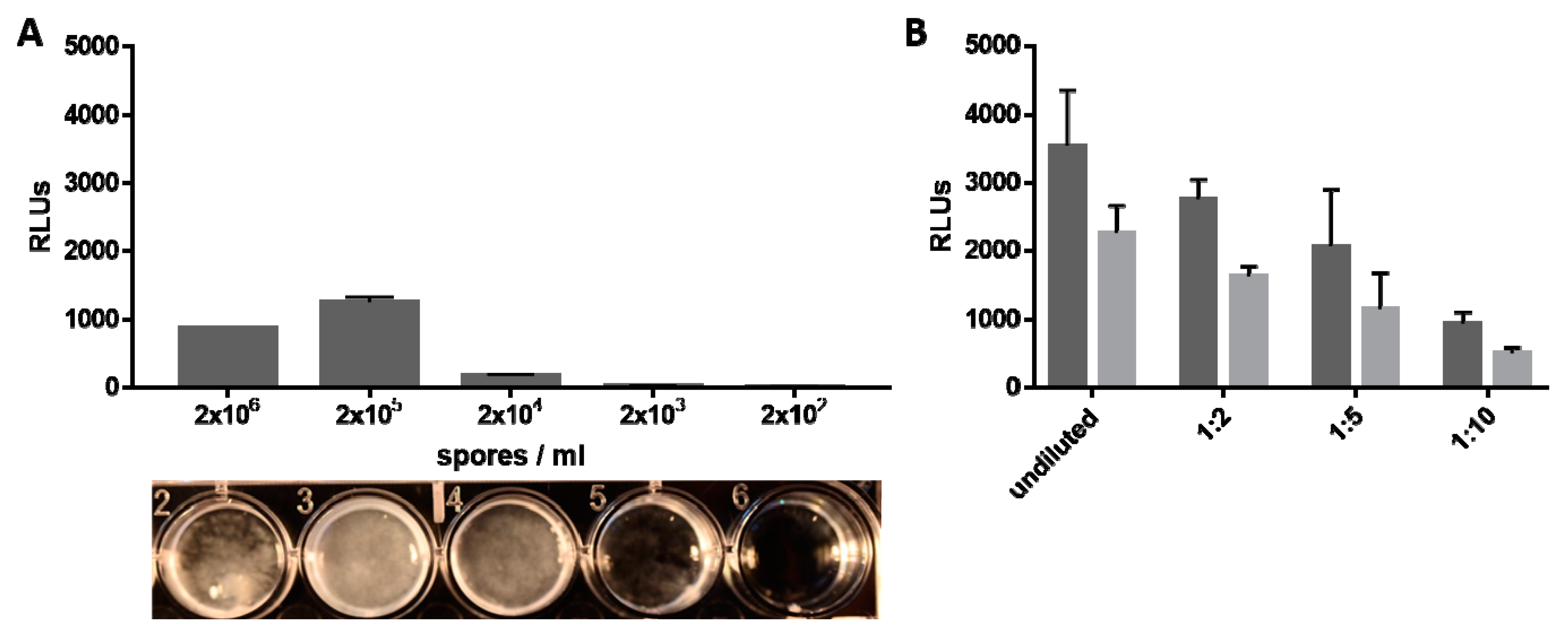
2.5. Luciferase Activity
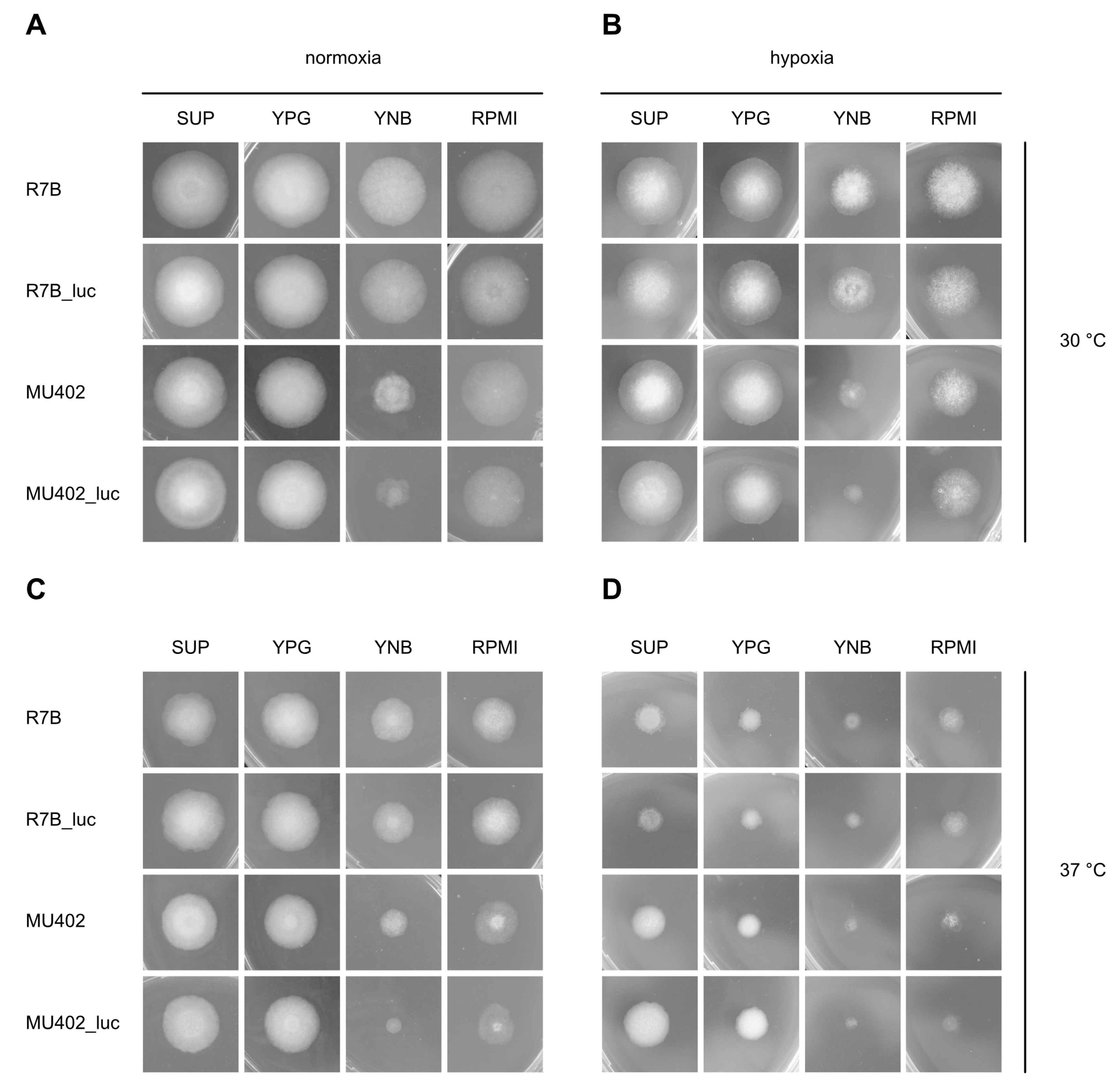
2.6. Phenotypic Analysis in Different Growth Conditions
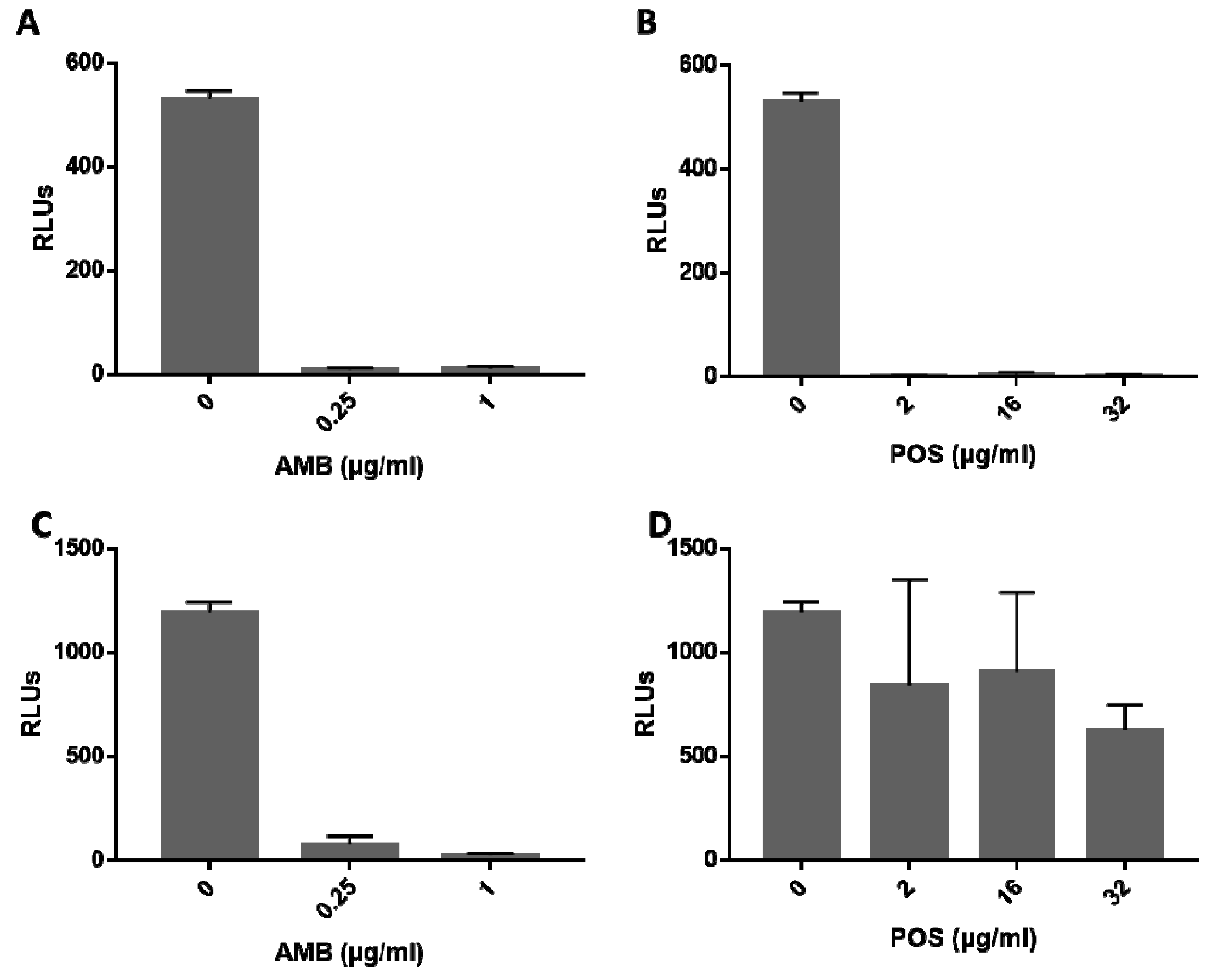
2.7. Antifungal Susceptibility Testing
2.8. Virulence Assay in Galleria mellonella
2.9. Histology of Larvae
3. Results and Discussion
3.1. Generation of Firefly Luciferase-Producing Mucor circinelloides Strains Resulted in Detectable Light Emission
3.2. In Vitro Characterization of Bioluminescent Mucor circinelloides Reporter Strains
3.2.1. Radial Growth Is Not Altered by Insertion of the Luciferase Gene
3.2.2. Light Emission Correlates with Fungal Biomass and Amount of Available Luciferase Substrate
3.3. Antifungal Susceptibility Testing
3.3.1. Genetic Manipulation Does Not Influence Antifungal Susceptibility Patterns of Mucor circinelloides Strains
3.3.2. Bioluminescent Strains Can Be Used to Evaluate Efficacy of Antifungal Drugs
3.4. Luciferase-Harboring Strains Exhibit Similar Virulence Potential as Recipient Strains in the Alternative Host Galleria mellonella
4. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Richardson, M. The ecology of the Zygomycetes and its impact on environmental exposure. Clin. Microbiol. Infect. 2009, 15, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Ingold, C.T. The Biology of Mucor and Its Allies; Studies in biology no. 88; E. Arnold: London, UK, 1978. [Google Scholar]
- Kontoyiannis, D.P.; Lionakis, M.S.; Lewis, R.E.; Chamilos, G.; Healy, M.; Perego, C.; Safdar, A.; Kantarjian, H.; Champlin, R.; Walsh, T.J.; et al. Zygomycosis in a tertiary-care cancer center in the era of Aspergillus-active antifungal therapy: A case-control observational study of 27 recent cases. J. Infect. Dis. 2005, 191, 1350–1360. [Google Scholar] [CrossRef] [PubMed]
- Lanternier, F.; Sun, H.Y.; Ribaud, P.; Singh, N.; Kontoyiannis, D.P.; Lortholary, O. Mucormycosis in organ and stem cell transplant recipients. Clin. Infect. Dis. 2012, 54, 1629–1636. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.E.; Kontoyiannis, D.P. Epidemiology and treatment of mucormycosis. Future Microbiol. 2013, 8, 1163–1175. [Google Scholar] [CrossRef] [PubMed]
- Chamilos, G.; Marom, E.M.; Lewis, R.E.; Lionakis, M.S.; Kontoyiannis, D.P. Predictors of pulmonary zygomycosis versus invasive pulmonary aspergillosis in patients with cancer. Clin. Infect. Dis. 2005, 41, 60–66. [Google Scholar] [CrossRef]
- Hammond, S.P.; Baden, L.R.; Marty, F.M. Mortality in hematologic malignancy and hematopoietic stem cell transplant patients with mucormycosis, 2001 to 2009. Antimicrob. Agents Chemother. 2011, 55, 5018–5021. [Google Scholar] [CrossRef]
- Spellberg, B.; Edwards, J., Jr.; Ibrahim, A. Novel perspectives on mucormycosis: Pathophysiology, presentation, and management. Clin. Microbiol. Rev. 2005, 18, 556–569. [Google Scholar] [CrossRef]
- Caramalho, R.; Tyndal, J.D.A.; Monk, B.C.; Larentis, T.; Lass-Flörl, C.; Lackner, M. Intrinsic short-tailed azole resistance in mucormycetes is due to an evolutionary conserved aminoacid substitution of the lanosterol 14α-demethylase. Sci. Rep. 2017, 7, 15898. [Google Scholar] [CrossRef] [Green Version]
- Skiada, A.; Pagano, L.; Groll, A.; Zimmerli, S.; Dupont, B.; Lagrou, K.; Lass-Florl, C.; Bouza, E.; Klimko, N.; Gaustad, P.; et al. Zygomycosis in Europe: Analysis of 230 cases accrued by the registry of the European Confederation of Medical Mycology (ECMM) working group on Zygomycosis between 2005 and 2007. Clin. Microbiol. Infect. 2011, 17, 1859–1867. [Google Scholar] [CrossRef]
- Antoniadou, A. Outbreaks of zygomycosis in hospitals. Clin. Microbiol. Infect. 2009, 15, 55–59. [Google Scholar] [CrossRef]
- Duffy, J.; Harris, J.; Gade, L.; Sehulster, L.; Newhouse, E.; O’Connell, H.; Noble-Wang, J.; Rao, C.; Balajee, S.A.; Chiller, T. Mucormycosis outbreak associated with hospital linens. Pediatr. Infect. Dis. J. 2014, 33, 472–476. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Hermoso, D.; Criscuolo, A.; Lee, S.C.; Legrand, M.; Chaouat, M.; Denis, B.; Lafaurie, M.; Rouveau, M.; Soler, C.; Schaal, J.-V.; et al. Outbreak of invasive wound mucormycosis in a burn unit due to multiple strains of Mucor circinelloides f. circinelloides resolved by whole-genome sequencing. MBio 2018, 9, e00573-18. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.C.; Billmyre, R.B.; Li, A.; Carson, S.; Sykes, S.M.; Huh, E.Y.; Mieczkowski, P.; Ko, D.C.; Cuomo, C.A.; Heitman, J. Analysis of a food-borne fungal pathogen outbreak: Virulence and genome of a Mucor circinelloides isolate from yogurt. MBio 2014, 5, e01390-14. [Google Scholar] [CrossRef] [PubMed]
- Magan, N.; Olsen, M. Mycotoxins in Food: Detection and Control; Woodhead Publishing Ltd.: Cambridge, UK, 2004. [Google Scholar]
- Torres-Martinez, S.; Ruiz-Vázquez, R.M.; Garre, V.; López-García, S.; Navarro, E.; Vila, A. Molecular tools for carotenogenesis analysis in the zygomycete Mucor circinelloides. Methods Mol. Biol. 2012, 898, 85–107. [Google Scholar] [PubMed]
- Vellanki, S.; Navarro-Mendoza, M.I.; Garcia, A.; Murcia, L.; Perez-Arques, C.; Garre, V.; Nicolas, F.E.; Lee, S.C. Mucor circinelloides: Growth, maintenance, and genetic manipulation. Curr. Protoc. Microbiol. 2018, 49, e53. [Google Scholar] [CrossRef] [PubMed]
- Nicolas, F.E.; de Haro, J.P.; Torres-Martínez, S.; Ruiz-Vázquez, R.M. Mutants defective in a Mucor circinelloides dicer-like gene are not compromised in siRNA silencing but display developmental defects. Fungal Genet. Biol. 2007, 44, 504–516. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Frometa, R.A.; Gutiérrez, A.; Torres-Martínez, S.; Garre, V. Malic enzyme activity is not the only bottleneck for lipid accumulation in the oleaginous fungus Mucor circinelloides. Appl. Microbiol. Biotechnol. 2013, 97, 3063–3072. [Google Scholar] [CrossRef] [PubMed]
- Brock, M. Application of bioluminescence imaging for in vivo monitoring of fungal infections. Int. J. Microbiol. 2012, 2012, 956794. [Google Scholar] [CrossRef] [PubMed]
- Delarze, E.; Ischer, F.; Sanglard, D.; Coste, A.T. Adaptation of a Gaussia princeps Luciferase reporter system in Candida albicans for in vivo detection in the Galleria mellonella infection model. Virulence 2015, 6, 684–693. [Google Scholar] [CrossRef]
- Jacobsen, I.D.; Lüttich, A.; Kurzai, O.; Hube, B.; Brock, M. In vivo imaging of disseminated murine Candida albicans infection reveals unexpected host sites of fungal persistence during antifungal therapy. J. Antimicrob. Chemother. 2014, 69, 2785–2796. [Google Scholar] [CrossRef]
- Brock, M.; Jouvion, G.; Droin-Bergère, S.; Dussurget, O.; Nicola, M.-A.; Ibrahim-Granet, O. Bioluminescent Aspergillus fumigatus, a new tool for drug efficiency testing and in vivo monitoring of invasive aspergillosis. Appl. Environ. Microbiol. 2008, 74, 7023–7035. [Google Scholar] [CrossRef] [PubMed]
- Donat, S.; Hasenberg, M.; Schäfer, T.; Ohlsen, K.; Gunzer, M.; Einsele, H.; Löffler, J.; Beilhack, A.; Krappmann, S. Surface display of Gaussia princeps luciferase allows sensitive fungal pathogen detection during cutaneous aspergillosis. Virulence 2012, 3, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim-Granet, O.; Jouvion, G.; Hohl, T.M.; Droin-Bergère, S.; Philippart, F.; Kim, O.Y.; Adib-Conquy, M.; Schwendener, R.; Cavaillon, J.M.; Brock, M.; et al. In vivo bioluminescence imaging and histopathopathologic analysis reveal distinct roles for resident and recruited immune effector cells in defense against invasive aspergillosis. BMC Microbiol. 2010, 10, 105. [Google Scholar] [CrossRef]
- Slesiona, S.; Ibrahim-Granet, O.; Olias, P.; Brock, M.; Jacobsen, I.D. Murine infection models for Aspergillus terreus pulmonary aspergillosis reveal long-term persistence of conidia and liver degeneration. J. Infect. Dis. 2012, 205, 1268–1277. [Google Scholar] [CrossRef] [PubMed]
- Roncero, M.I.G. Enrichment Method for the isolation of auxotrophic mutants of mucor using the polyene antibiotic N-glycosyl-polifungin. Carlsberg Res. Commun. 1984, 49, 685–690. [Google Scholar] [CrossRef]
- Nicolas, F.E.; Navarro-Mendoza, M.I.; Pérez-Arques, C.; López-García, S.; Navarro, E.; Torres-Martínez, S.; Garre, V. Molecular tools for carotenogenesis analysis in the mucoral Mucor circinelloides. Methods Mol. Biol. 2018, 1852, 221–237. [Google Scholar]
- Maurer, E.; Hörtnagl, C.; Lackner, M.; Grässle, D.; Naschberger, V.; Moser, P.; Segal, E.; Semis, M.; Lass-Flörl, C.; Binder, U. Galleria mellonella as a model system to study virulence potential of mucormycetes and evaluation of antifungal treatment. Med. Mycol. 2018. [Google Scholar] [CrossRef]
- Arendrup, M.C.; Hope, W.W.; Lass-Flörl, C.; Cuenca-Estrella, M.; Arikan, S.; Barchiesi, F.; Bille, J.; Chryssanthou, E.; Groll, A.; et al. EUCAST technical note on the EUCAST definitive document EDef 7.2: Method for the determination of broth dilution minimum inhibitory concentrations of antifungal agents for yeasts EDef 7.2 (EUCAST-AFST). Clin. Microbiol. Infect. 2012, 18, E246–E247. [Google Scholar] [CrossRef]
- Fallon, J.; Kelly, J.; Kavanagh, K. Galleria mellonella as a model for fungal pathogenicity testing. Methods Mol. Biol. 2012, 845, 469–485. [Google Scholar]
- Maurer, E.; Browne, N.; Surlis, C.; Jukic, E.; Moser, P.; Kavanagh, K.; Lass-Flörl, C.; Binder, U. Galleria mellonella as a host model to study Aspergillus terreus virulence and amphotericin B resistance. Virulence 2015, 6, 591–598. [Google Scholar] [CrossRef]
- Grahl, N.; Shepardson, K.M.; Chung, D.; Cramer, R.A. Hypoxia and fungal pathogenesis: To air or not to air? Eukaryot. Cell 2012, 11, 560–570. [Google Scholar] [CrossRef] [PubMed]
- Dannaoui, E. Antifungal resistance in mucorales. Int. J. Antimicrob. Agents 2017, 50, 617–621. [Google Scholar] [CrossRef] [PubMed]
- Espinel-Ingroff, A.; Chakrabarti, A.; Chowdhary, A.; Cordoba, S.; Dannaoui, E.; Dufresne, P.; Fothergill, A.; Ghannoum, M.; Gonzalez, G.M.; Guarro, J. Multicenter evaluation of MIC distributions for epidemiologic cutoff value definition to detect amphotericin B, posaconazole, and itraconazole resistance among the most clinically relevant species of Mucorales. Antimicrob. Agents Chemother. 2015, 59, 1745–1750. [Google Scholar] [CrossRef] [PubMed]
- Khan, Z.U.; Ahmad, S.; Brazda, A.; Chandy, R. Mucor circinelloides as a cause of invasive maxillofacial zygomycosis: an emerging dimorphic pathogen with reduced susceptibility to posaconazole. J. Clin. Microbiol. 2009, 47, 1244–1248. [Google Scholar] [CrossRef] [PubMed]
- Perkhofer, S.; Locher, M.; Cuenca-Estrella, M.; Rüchel, R.; Würzner, R.; Dierich, M.P.; Lass-Flörl, C. Posaconazole enhances the activity of amphotericin B against hyphae of zygomycetes in vitro. Antimicrob. Agents Chemother. 2008, 52, 2636–2638. [Google Scholar] [CrossRef]
- Salas, V.; Pastor, F.J.; Calvo, E.; Alvarez, E.; Sutton, D.A.; Mayayo, E.; Fothergill, A.W.; Rinaldi, M.G.; Guarro, J. In vitro and in vivo activities of posaconazole and amphotericin B in a murine invasive infection by Mucor circinelloides: poor efficacy of posaconazole. Antimicrob. Agents Chemother. 2012, 56, 2246–2250. [Google Scholar] [CrossRef]
- Caramalho, R.; Maurer, E.; Binder, U.; Araújo, R.; Dolatabadi, S.; Lass-Flörl, C.; Lackner, M. Etest cannot be recommended for in vitro susceptibility testing of mucorales. Antimicrob. Agents Chemother. 2015, 59, 3663–3665. [Google Scholar] [CrossRef] [PubMed]
- Vitale, R.G.; de Hoog, G.S.; Schwarz, P.; Dannaoui, E.; Deng, S.; Machouart, M.; Voigt, K.; van de Sande, W.W.; Dolatabadi, S.; Meis, J.F.; et al. Antifungal susceptibility and phylogeny of opportunistic members of the order mucorales. J. Clin. Microbiol. 2012, 50, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, M.M.; Pastor, F.J.; Sutton, D.A.; Calvo, E.; Fothergill, A.W.; Salas, V.; Rinaldi, M.G.; Guarro, J. Correlation between in vitro activity of posaconazole and in vivo efficacy against Rhizopus oryzae infection in mice. Antimicrob. Agents Chemother. 2010, 54, 1665–1669. [Google Scholar] [CrossRef]
- D’Enfert, C.; Diaquin, M.; Delit, A.; Wuscher, N.; Debeaupuis, J.P.; Huerre, M.; Latge, J.P. Attenuated virulence of uridine-uracil auxotrophs of Aspergillus fumigatus. Infect. Immun. 1996, 64, 4401–4405. [Google Scholar]
- Galiger, C.; Brock, M.; Jouvion, G.; Savers, A.; Parlato, M.; Ibrahim-Granet, O. Assessment of efficacy of antifungals against Aspergillus fumigatus: Value of real-time bioluminescence imaging. Antimicrob. Agents Chemother. 2013, 57, 3046–3059. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Genotypes/Characteristics | Reference | |
|---|---|---|---|
| Mucor circinelloides | R7B | leuA1 (mutant alelle of leuA gene) | Roncero et al., 1984 [27] |
| R7B_luc | carRP::leu | Obtained in this study | |
| R7B_luc1 | carRP::leu | Obtained in this study | |
| MU402 | leuA1, pyrG− | Nicolas et al., 2007 [18] | |
| MU402_luc | pyrG−, carRP::leuA | Obtained in this study | |
| MU402_luc1 | pyrG−, carRP::leuA | Obtained in this study | |
| DH5α | ampicillin resistance | Thermo Fisher (Germering, Germany) | |
| Escherichia coli | pMAT1477 | Pzrt1, leuA | Rodriguez-Frometa et al., 2013 [19] |
| pGL3 basic vector | firefly luciferase reporter vector | Commercially available, Promega (Fitchburg, WI, USA) | |
| Plasmids | pMAT1903 | pMAT1477 + luciferase | Obtained in this study |
| Strains | RLUs (2 min) | SD | RLUs (30 min) | SD |
|---|---|---|---|---|
| R7B | 14 | 4 | 15 | 1 |
| R7B_luc | 4167 | 112 | 1444 | 129 |
| R7B_luc1 | 3117 | 73 | 1194 | 95 |
| MU402 | 10 | 1 | 7 | 3 |
| MU402_luc | 174 | 50 | 185 | 0 |
| MU402_luc1 | 20 | 2 | 15 | 1 |
| MIC (µg/mL) | ||||||||
|---|---|---|---|---|---|---|---|---|
| RPMI1640 | YNB | |||||||
| Strains | AMB | POS | ITRA | ISA | AMB | POS | ITRA | ISA |
| R7B | 2 | >16 | >8 | >4 | 1 | 2 | >8 | >4 |
| R7B_luc | 1 | >16 | >8 | >4 | 0.5 | 4 | >8 | >4 |
| MU402 | 2 | >16 | >8 | >4 | 1 | >16 | >8 | >4 |
| MU402_luc | 2 | >16 | >8 | >4 | 1 | >16 | >8 | >4 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Binder, U.; Navarro-Mendoza, M.I.; Naschberger, V.; Bauer, I.; Nicolas, F.E.; Pallua, J.D.; Lass-Flörl, C.; Garre, V. Generation of A Mucor circinelloides Reporter Strain—A Promising New Tool to Study Antifungal Drug Efficacy and Mucormycosis. Genes 2018, 9, 613. https://doi.org/10.3390/genes9120613
Binder U, Navarro-Mendoza MI, Naschberger V, Bauer I, Nicolas FE, Pallua JD, Lass-Flörl C, Garre V. Generation of A Mucor circinelloides Reporter Strain—A Promising New Tool to Study Antifungal Drug Efficacy and Mucormycosis. Genes. 2018; 9(12):613. https://doi.org/10.3390/genes9120613
Chicago/Turabian StyleBinder, Ulrike, Maria Isabel Navarro-Mendoza, Verena Naschberger, Ingo Bauer, Francisco E. Nicolas, Johannes D. Pallua, Cornelia Lass-Flörl, and Victoriano Garre. 2018. "Generation of A Mucor circinelloides Reporter Strain—A Promising New Tool to Study Antifungal Drug Efficacy and Mucormycosis" Genes 9, no. 12: 613. https://doi.org/10.3390/genes9120613
APA StyleBinder, U., Navarro-Mendoza, M. I., Naschberger, V., Bauer, I., Nicolas, F. E., Pallua, J. D., Lass-Flörl, C., & Garre, V. (2018). Generation of A Mucor circinelloides Reporter Strain—A Promising New Tool to Study Antifungal Drug Efficacy and Mucormycosis. Genes, 9(12), 613. https://doi.org/10.3390/genes9120613