Transcriptomic Analysis of Flower Bud Differentiation in Magnolia sinostellata

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Morphological Observation
2.3. RNA Extraction and cDNA Library Construction
2.4. Illumina Sequencing, Assembly and Annotation
2.5. Differential Gene Expression Analysis
2.6. qRT-PCR Analysis
3. Results
3.1. Morphological Changes during the Different Stages of Flower Bud Differentiation
3.2. Differentiating Flower Bud Transcriptome Sequencing, Assembly, and Functional Annotation
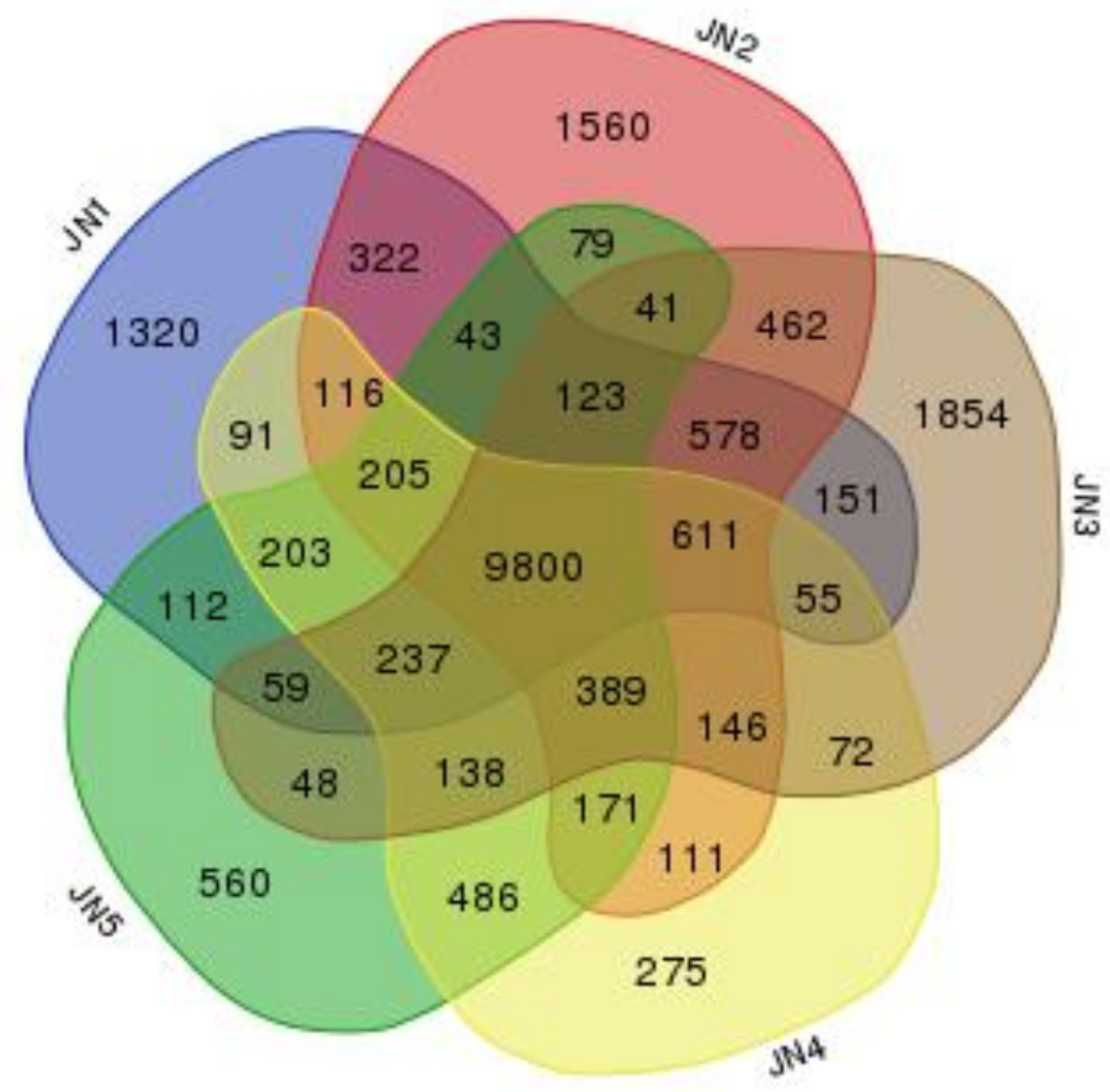
3.3. Genes Expression in Each Stage of Flower Bud Differentiation
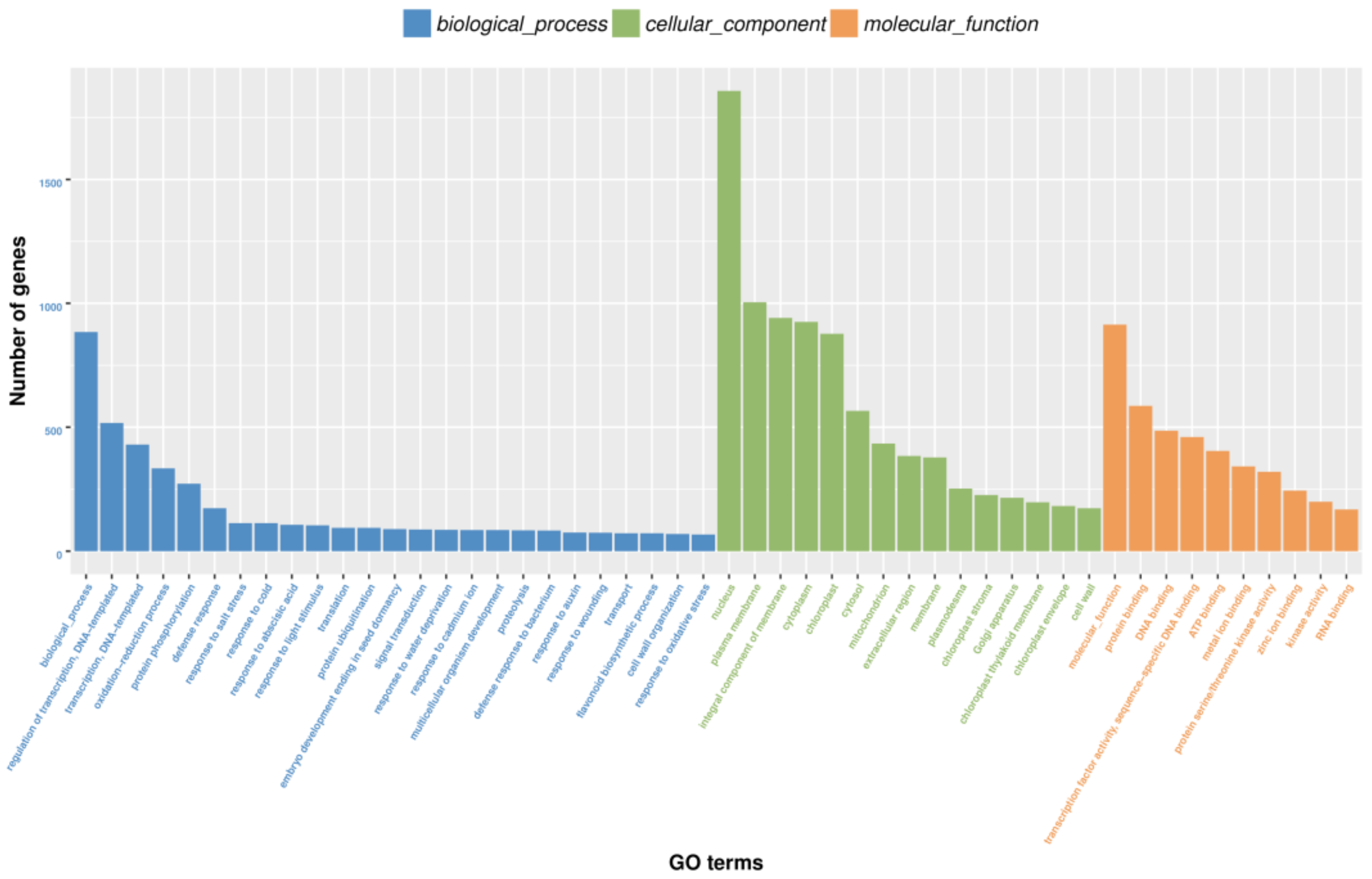
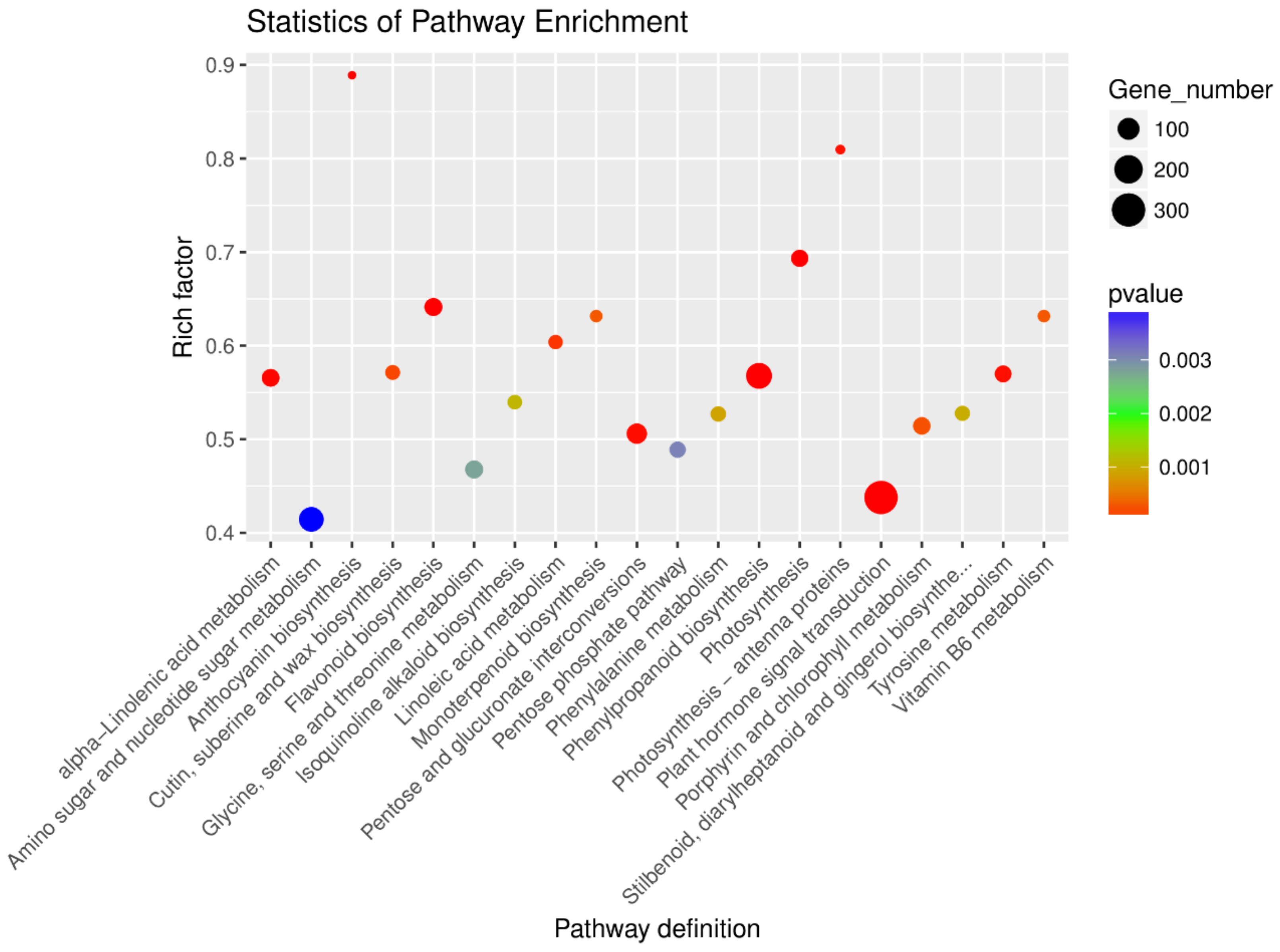
3.4. GO and KEGG Enrichment Analysis of Differentially Expressed Genes
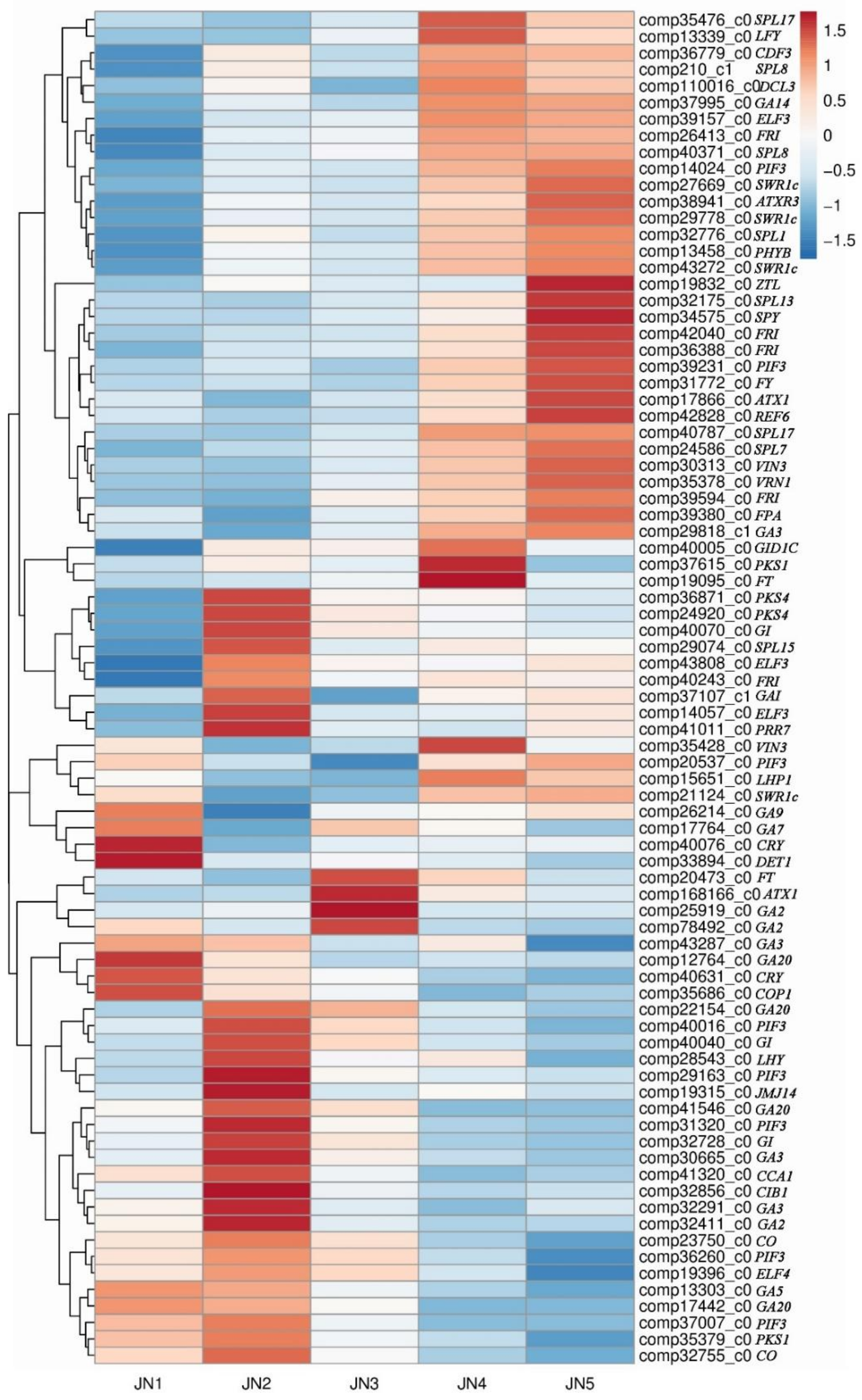
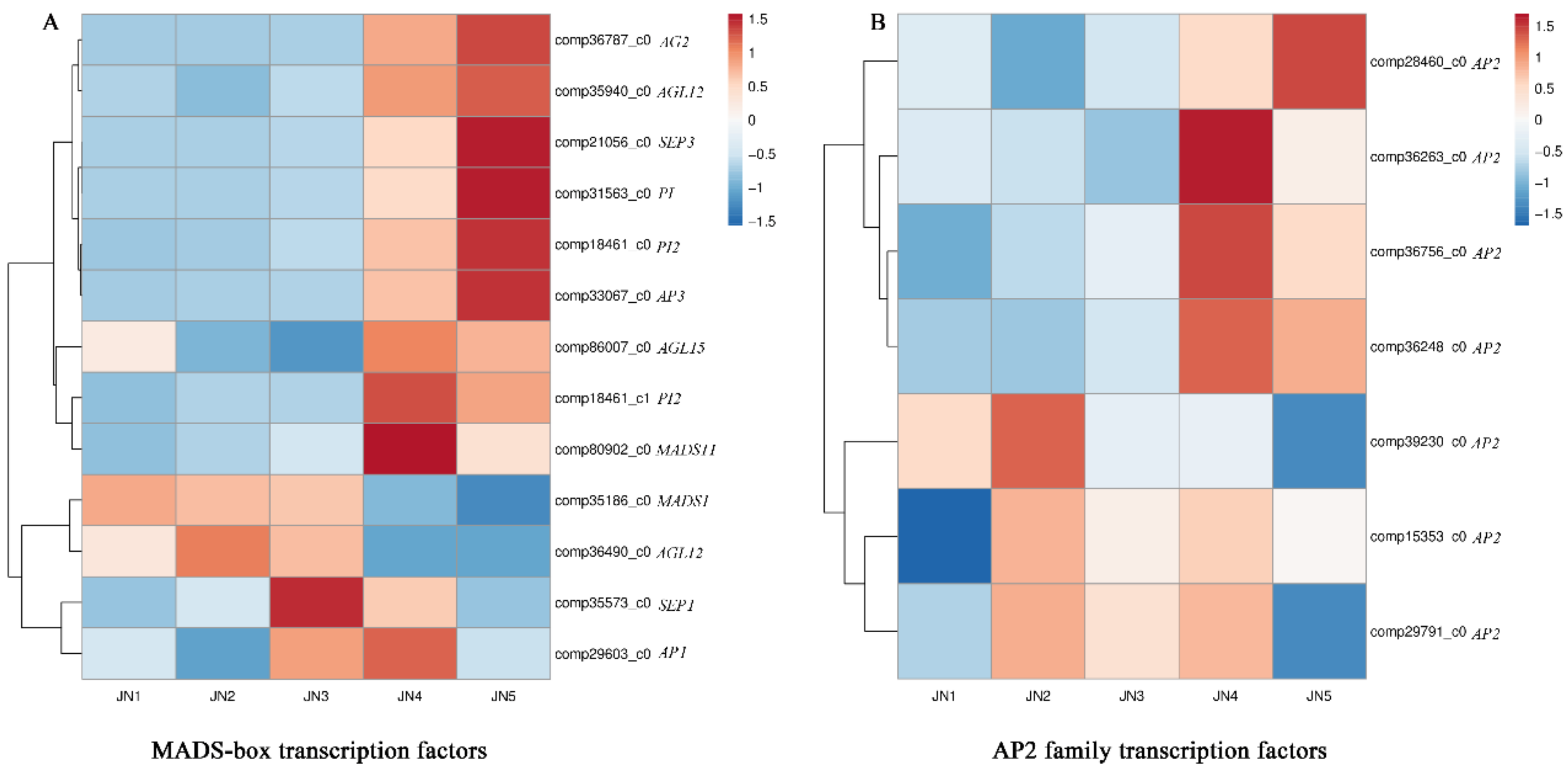
3.5. DEGs and Transcription Factors Associated with the Five Stages of Flower Bud Differentiation
3.6. Verification of Relative Gene Expression in Different Stages of Flower Bud Differentiation
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zhang, Y.; Zhou, Y.; Chen, Q.; Huang, X.; Tian, C. Molecular basis of flowering time regulation in Arabidopsis. Chin. Bull. Bot. 2014, 49, 469–482. [Google Scholar]
- Mouradov, A.; Cremer, F.; Coupland, G. Control of flowering time: Interacting pathways as a basis for diversity. Plant Cell 2002, 14, S111–S130. [Google Scholar] [CrossRef] [PubMed]
- Fornara, F.; de Montaigu, A.; Coupland, G. Snapshot: Control of flowering in Arabidopsis. Cell 2010, 141, 550. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Lee, I. Regulation and function of SOC1, a flowering pathway integrator. J. Exp. Bot. 2010, 61, 2247–2254. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, O.; Lee, I.; Blázquez, M.A.; Weigel, D. Flowering-time genes modulate the response to leafy activity. Genetics 1998, 150, 403–410. [Google Scholar] [PubMed]
- Wagner, D.; Sablowski, R.W.; Meyerowitz, E.M. Transcriptional activation of APETALA1 by LEAFY. Science 1999, 285, 582–584. [Google Scholar] [CrossRef] [PubMed]
- Mimida, N.; Kidou, S.-I.; Iwanami, H.; Moriya, S.; Abe, K.; Voogd, C.; Varkonyi-Gasic, E.; Kotoda, N. Apple FLOWERING LOCUS T proteins interact with transcription factors implicated in cell growth and organ development. Tree Physiol. 2011, 31, 555–566. [Google Scholar] [CrossRef] [PubMed]
- Winterhagen, P.; Tiyayon, P.; Samach, A.; Hegele, M.; Wünsche, J.N. Isolation and characterization of fLOWERING LOCUS T subforms and APETALA1 of the subtropical fruit tree Dimocarpus longan. Plant Physiol. Biochem. 2013, 71, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, F.; Endo, T.; Shimada, T.; Fujii, H.; Shimizu, T.; Omura, M.; Ikoma, Y. Increased CiFT abundance in the stem correlates with floral induction by low temperature in Satsuma mandarin (Citrus unshiu Marc.). J. Exp. Bot. 2007, 58, 3915–3927. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.-Y.; Adams, J.P.; Kim, H.; No, K.; Ma, C.; Strauss, S.H.; Drnevich, J.; Vandervelde, L.; Ellis, J.D.; Rice, B.M. FLOWERING LOCUS T duplication coordinates reproductive and vegetative growth in perennial poplar. Proc. Natl. Acad. Sci. USA 2011, 108, 10756–10761. [Google Scholar] [CrossRef] [PubMed]
- Coen, E. Goethe and the ABC model of flower development. Comptes Rendus de l’Académie des Sciences-Series III-Sciences de la Vie 2001, 324, 523–530. [Google Scholar] [CrossRef]
- Coen, E.S.; Meyerowitz, E.M. The war of the whorls: Genetic interactions controlling flower development. Nature 1991, 353, 31. [Google Scholar] [CrossRef] [PubMed]
- Litt, A.; Kramer, E.M. Seminars in cell & developmental biology. In The abc Model and the Diversification of Floral Organ Identity; Elsevier: New York, NY, USA, 2010; pp. 129–137. [Google Scholar]
- Angenent, G.C.; Franken, J.; Busscher, M.; van Dijken, A.; van Went, J.L.; Dons, H.; van Tunen, A.J. A novel class of MADS box genes is involved in ovule development in petunia. Plant Cell 1995, 7, 1569–1582. [Google Scholar] [CrossRef] [PubMed]
- Pelaz, S.; Ditta, G.S.; Baumann, E.; Wisman, E.; Yanofsky, M.F. B and c floral organ identity functions require sepallata MADS-box genes. Nature 2000, 405, 200. [Google Scholar] [CrossRef] [PubMed]
- Bowman, J.L.; Smyth, D.R.; Meyerowitz, E.M. The ABC model of flower development: Then and now. Development 2012, 139, 4095–4098. [Google Scholar] [CrossRef] [PubMed]
- Theißen, G. Development of floral organ identity: Stories from the MADS house. Curr. Opin. Plant Biol. 2001, 4, 75–85. [Google Scholar] [CrossRef]
- Kim, S.; Koh, J.; Yoo, M.J.; Kong, H.; Hu, Y.; Ma, H.; Soltis, P.S.; Soltis, D.E. Expression of floral MADS-box genes in basal angiosperms: Implications for the evolution of floral regulators. Plant J. 2005, 43, 724–744. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.-Z.; Hilu, K.; Wang, Y.-L. From leaf and branch into a flower: Magnolia tells the story. Bot. Stud. 2014, 55, 28. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Rudall, P. Comparative floral anatomy and ontogeny in magnoliaceae. Plant Syst. Evol. 2006, 258, 1–15. [Google Scholar] [CrossRef]
- Wu, Z.; Hu, J.; Si, H.; Tang, J.; Sun, J.; Hu, Y. An observation on differentiation of Magnolia denudata Desr. flower buds. China J. Chin. Mater. Med. 1991, 2, 4. [Google Scholar]
- Jing, D.; Liu, Z.; Zhang, B.; Ma, J.; Han, Y.; Chen, F. Two ancestral APETALA3 homologs from the basal angiosperm Magnolia wufengensis (Magnoliaceae) can affect flower development of Arabidopsis. Gene 2014, 537, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Chen, F.; Jing, D.; Liu, Z.; Ma, L. Isolation and characterization of an AGAMOUS-like gene from Magnolia wufengensis (Magnoliaceae). Plant Mol. Biol. Rep. 2012, 30, 690–698. [Google Scholar] [CrossRef]
- Shi, S.-G.; Yang, M.; Zhang, M.; Wang, P.; Kang, Y.-X.; Liu, J.-J. Genome-wide transcriptome analysis of genes involved in flavonoid biosynthesis between red and white strains of Magnolia sprengeri pamp. BMC Genom. 2014, 15, 706. [Google Scholar] [CrossRef] [PubMed]
- Dhandapani, S.; Jin, J.; Sridhar, V.; Sarojam, R.; Chua, N.-H.; Jang, I.-C. Integrated metabolome and transcriptome analysis of Magnolia champaca identifies biosynthetic pathways for floral volatile organic compounds. BMC Genom. 2017, 18, 463. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.-Z. Magnolia sinostellata and relatives (Magnoliaceae). Phytotaxa 2013, 154, 47–58. [Google Scholar]
- Xia, N.H.; Liu, Y.H.; Nooteboom, H.P. Magnoliaceae. In Flora of China; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MO, USA, 2008; pp. 47–91. [Google Scholar]
- Fan, L.; Chen, M.; Wang, N.; Shen, Y.; Zhang, S. Flower bud differention of three species of magnolia. J. Northeast For. Univ. 2018, 46, 27–30. [Google Scholar]
- Xu, Y.C.; Zhou, L.H.; Hu, S.Q.; Hao, R.M.; Huang, C.j.; Zhao, H.-B. The differentiation and development of pistils of hermaphrodites and pistillodes of males in androdioecious Osmanthus fragrans L. and implications for the evolution to androdioecy. Plant Syst. Evol. 2014, 300, 843–849. [Google Scholar] [CrossRef]
- Andrews, S. Fastqc: A Quality Control Tool for High Throughput Sequence Data; Babraham Institute: Cambridge, UK, 2010. [Google Scholar]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M. De novo transcript sequence reconstruction from rna-seq using the trinity platform for reference generation and analysis. Nat. Protocols 2013, 8, 1494–1512. [Google Scholar] [CrossRef] [PubMed]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621. [Google Scholar] [CrossRef] [PubMed]
- Srikanth, A.; Schmid, M. Regulation of flowering time: All roads lead to Rome. Cell. Mol. Life Sci. 2011, 68, 2013–2037. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔct method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Boeckmann, B.; Bairoch, A.; Apweiler, R.; Blatter, M.C.; Estreicher, A.; Gasteiger, E.; Martin, M.J.; Michoud, K.; O’donovan, C.; Phan, I. The SWISS-PROT protein knowledgebase and its supplement TrEMBL in 2003. Nucleic Acids Res. 2003, 31, 365–370. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Godzik, A. Cd-hit: A fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S. Kegg: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Krylov, D.M.; Wolf, Y.I.; Rogozin, I.B.; Koonin, E.V. Gene loss, protein sequence divergence, gene dispensability, expression level and interactivity are correlated in eukaryotic evolution. Genome Res. 2003, 13, 2229–2235. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Zhang, Y.; Xu, Z.; Luan, A.; Mao, Q.; Feng, J.; Xie, T.; Gong, X.; Wang, X.; Chen, H. Transcriptome profiling of the pineapple under low temperature to facilitate its breeding for cold tolerance. PLoS ONE 2016, 11, e0163315. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xiu, Y.; Bi, K.; Ou, J.; Gu, W.; Wang, W.; Meng, Q. Integrated analysis of mrna-seq in the haemocytes of eriocheir sinensis in response to spiroplasma eriocheiris infection. Fish Shellfish Immunol. 2017, 68, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Bateman, A.; Coin, L.; Durbin, R.; Finn, R.D.; Hollich, V.; Griffiths-Jones, S.; Khanna, A.; Marshall, M.; Moxon, S.; Sonnhammer, E.L. The pfam protein families database. Nucleic Acids Res. 2004, 32, D138–D141. [Google Scholar] [CrossRef] [PubMed]
- Consortium, G.O. The Gene Ontology (GO) database and informatics resource. Nucleic Acids Res. 2004, 32, D258–D261. [Google Scholar] [CrossRef] [PubMed]
- Peng, F.; Tian, M.; Wang, C. Morphological and anatomical characteristics along with nutrient content changes during flower bud differentiation in oncidium. J. Zhejiang Agric. Univ. 2012, 29, 7–11. [Google Scholar]
- Pang, F.; Zhao, M.; Wang, Y.; Yu, H.; Xia, J. Studies on floral bud differentiation and biochemical changes of strawberry ‘ningyu’. J. Fruit Sci. 2014, 31, 1117–1122. [Google Scholar]
- Niu, H.; Zhang, H.; Yuan, B. Flower formation and endogenous hormones dynamic in chinese jujube. Acta Hortic. Sin. 2015, 42, 655–664. [Google Scholar]
- Liu, L.Q.; Qin, W.; Liao, K.; He, F.-J.; Zhang, D.-H.; Xu, L.; Fan, W.-M. Study on the floral morphodifferentiation of some apricot cultivars in Xinjiang. Xinjiang Agric. Sci. 2007, 5, 004. [Google Scholar]
- Chao, W.; Long, Z.; Ni, P.; Xin, Z. Morphological differentiation of flower buds of Cerasus tianschanica Pojark. Acta Agric. Boreali-Occident. Sin. 2015, 5, 20. [Google Scholar]
- Liu, H.; Sun, M.; Du, D.; Pan, H.; Cheng, T.; Wang, J.; Zhang, Q. Whole-transcriptome analysis of differentially expressed genes in the vegetative buds, floral buds and buds of Chrysanthemum morifolium. PLoS ONE 2015, 10, e0128009. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, N.; Wu, M.F.; Winter, C.M.; Berns, M.C.; Nole-Wilson, S.; Yamaguchi, A.; Coupland, G.; Krizek, B.A.; Wagner, D. A molecular framework for auxin-mediated initiation of flower primordia. Dev. Cell 2013, 24, 271. [Google Scholar] [CrossRef] [PubMed]
- Matsoukas, I.G. Interplay between sugar and hormone signaling pathways modulate floral signal transduction. Front. Genet. 2014, 5, 218. [Google Scholar] [CrossRef] [PubMed]
- Apel, L.; Kammerer, D.R.; Stintzing, F.C.; Spring, O. Comparative metabolite profiling of triterpenoid saponins and flavonoids in flower color mutations of Primula veris L. Int. J. Mol. Sci. 2017, 18, 153. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Tang, H.; Dong, X.; Hou, Y.; Luo, Y.; Jiang, Y.; Huang, Q. Progress in the study of plant MYB transcription factors. Genom. Appl. Biol. 2009, 28, 365–372. [Google Scholar]
- Dubos, C.; Stracke, R.; Grotewold, E.; Weisshaar, B.; Martin, C.; Lepiniec, L. MYB transcription factors in Arabidopsis. Trends Plant Sci. 2010, 15, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Palatnik, J.F.; Allen, E.; Wu, X.; Schommer, C.; Schwab, R.; Carrington, J.C.; Weigel, D. Control of leaf morphogenesis by microRNAs. Nature 2003, 425, 257. [Google Scholar] [CrossRef] [PubMed]
- Park, W.; Li, J.; Song, R.; Messing, J.; Chen, X. CARPEL factory, a dicer homolog and HEN1, a novel protein, act in microRNA metabolism in Arabidopsis thaliana. Curr. Biol. 2002, 12, 1484–1495. [Google Scholar] [CrossRef]
- Chen, N.; Jiang, J.; Cao, B.; Lei, J.; Chen, C. The latest progresses on plant NAC transcription factors function. Mol. Plant Breed. 2015, 13, 1407–1414. [Google Scholar]
- Rushton, P.J.; Somssich, I.E.; Ringler, P.; Shen, Q.J. WRKY transcription factors. Trends Plant Sci. 2010, 15, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Hayama, R.; Coupland, G. Shedding light on the circadian clock and the photoperiodic control of flowering. Curr. Opin. Plant Biol. 2003, 6, 13–19. [Google Scholar] [CrossRef]
- Reyes, J.C.; Muro-Pastor, M.I.; Florencio, F.J. The GATA family of transcription factors in Arabidopsis and rice. Plant Physiol. 2004, 134, 1718–1732. [Google Scholar] [CrossRef] [PubMed]
- Wagner, D.; Meyerowitz, E.M. Switching on flowers: Transient LEAFY induction reveals novel aspects of the regulation of reproductive development in Arabidopsis. Front. Plant Sci. 2011, 2, 60. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ballard, H.E.; Sternberger, A.L.; Wyatt, S.E.; Stockinger, E.J.; Nadella, V. The potential role of two LEAFY orthologs in the chasmogamous/cleistogamous mixed breeding system of Viola pubescens (Violaceae). J. Torrey Bot. Soc. 2017, 144, 206–217. [Google Scholar] [CrossRef]
- Caicedo, A.L.; Stinchcombe, J.R.; Olsen, K.M.; Schmitt, J.; Purugganan, M.D. Epistatic interaction between Arabidopsis FRI and FLC flowering time genes generates a latitudinal cline in a life history trait. Proc. Natl. Acad. Sci. USA 2004, 101, 15670–15675. [Google Scholar] [CrossRef] [PubMed]
- He, Y. Control of the transition to flowering by chromatin modifications. Mol. Plant 2009, 2, 554–564. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.W.; Czech, B.; Weigel, D. miR156-regulated SPL transcription factors define an endogenous flowering pathway in Arabidopsis thaliana. Cell 2009, 138, 738–749. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, A.; Wu, M.F.; Yang, L.; Wu, G.; Poethig, R.S.; Wagner, D. The microRNA-regulated SBP-Box transcription factor SPL3 is a direct upstream activator of LEAFY, FRUITFULL and APETALA1. Dev. Cell 2009, 17, 268–278. [Google Scholar] [CrossRef] [PubMed]
- Galvão, V.C.; Horrer, D.; Küttner, F.; Schmid, M. Spatial control of flowering by DELLA proteins in Arabidopsis thaliana. Development 2012, 139, 4072–4082. [Google Scholar] [CrossRef] [PubMed]
- Pařenicová, L.; de Folter, S.; Kieffer, M.; Horner, D.S.; Favalli, C.; Busscher, J.; Cook, H.E.; Ingram, R.M.; Kater, M.M.; Davies, B. Molecular and phylogenetic analyses of the complete MADS-box transcription factor family in Arabidopsis. Plant Cell Online 2003, 15, 1538–1551. [Google Scholar] [CrossRef]
- Jaya, E.S.; Clemens, J.; Song, J.; Zhang, H.; Jameson, P.E. Quantitative expression analysis of meristem identity genes in Eucalyptus occidentalis: AP1 is an expression marker for flowering. Tree Physiol. 2009, 30, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.Q.; Li, Z.Y.; Zhao, D.Q.; Tao, J. Comparative analysis of flower-meristem-identity gene APETALA2 (AP2) codon in different plant species. J. Integr. Agric. 2017, 6, 60345–60347. [Google Scholar] [CrossRef]
- Kim, S.; Yoo, M.-J.; Albert, V.A.; Farris, J.S.; Soltis, P.S.; Soltis, D.E. Phylogeny and diversification of b-function MADS-box genes in angiosperms: Evolutionary and functional implications of a 260-million-year-old duplication. Am. J. Bot. 2004, 91, 2102–2118. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Number | Unigenes Annotated Using These Databases | ||||||
|---|---|---|---|---|---|---|---|
| Nr | Pfam | KEGG | KOG | Swiss-Prot | GO | ||
| Unigenes | 52,441 | 25,531 | 18,736 | 8612 | 20,977 | 15,850 | 14,055 |
| Percentage | 100% | 48.69% | 35.73% | 16.42% | 40.00% | 30.22% | 26.80% |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, L.; Chen, M.; Dong, B.; Wang, N.; Yu, Q.; Wang, X.; Xuan, L.; Wang, Y.; Zhang, S.; Shen, Y. Transcriptomic Analysis of Flower Bud Differentiation in Magnolia sinostellata. Genes 2018, 9, 212. https://doi.org/10.3390/genes9040212
Fan L, Chen M, Dong B, Wang N, Yu Q, Wang X, Xuan L, Wang Y, Zhang S, Shen Y. Transcriptomic Analysis of Flower Bud Differentiation in Magnolia sinostellata. Genes. 2018; 9(4):212. https://doi.org/10.3390/genes9040212
Chicago/Turabian StyleFan, Lijie, Mengqian Chen, Bin Dong, Ninghang Wang, Qin Yu, Xingli Wang, Lingjuan Xuan, Yaling Wang, Shouzhou Zhang, and Yamei Shen. 2018. "Transcriptomic Analysis of Flower Bud Differentiation in Magnolia sinostellata" Genes 9, no. 4: 212. https://doi.org/10.3390/genes9040212
APA StyleFan, L., Chen, M., Dong, B., Wang, N., Yu, Q., Wang, X., Xuan, L., Wang, Y., Zhang, S., & Shen, Y. (2018). Transcriptomic Analysis of Flower Bud Differentiation in Magnolia sinostellata. Genes, 9(4), 212. https://doi.org/10.3390/genes9040212