3.1. Trait Expressed in Both Sexes
We first assume that both sexes fully express the trait that is under divergent ecological selection and used, in males, as a mating cue. An example of such a trait might be body size. We begin by concentrating on the case of secondary contact after a period of almost complete divergence in allopatry, to ask how preference strength and physical linkage will determine the trait and preference divergence that can be maintained after the commencement of gene flow. For this initial analysis, we assume that the preference and trait are in complete linkage disequilibrium at the start of secondary contact; this allows us to equate the case of no recombination (r = 0) between the preference and trait loci with phenotype matching.
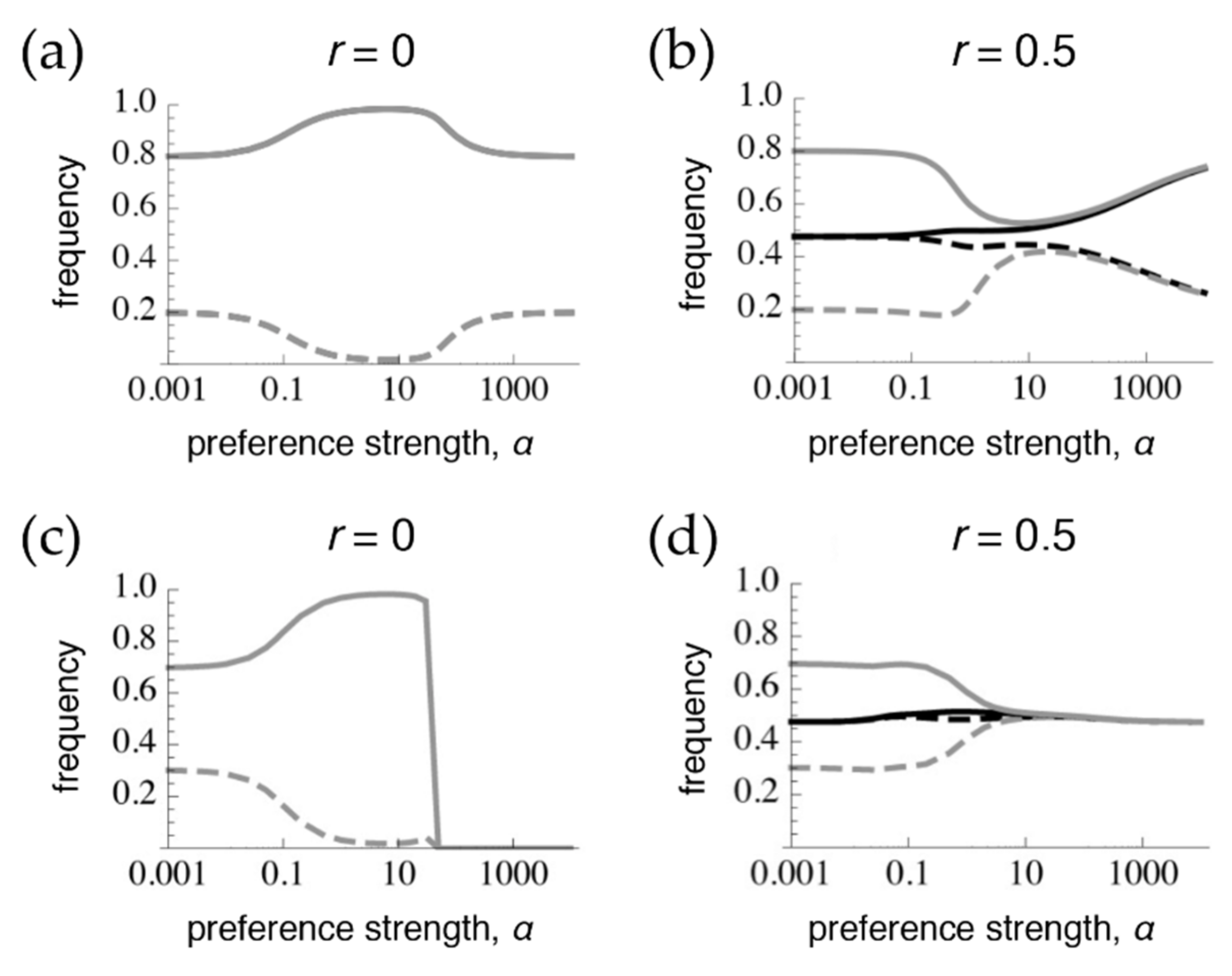
In order to assess whether tight linkage between the preference and trait can mimic pleiotropy, we first obtain, for comparison under these parameter values and starting conditions, the divergence that results after 1 million generations (at or close to equilibrium) at the extremes of pleiotropy (
Figure 1a, following [
11,
15]) versus the case where preferences and traits are on different chromosomes (
Figure 1b, following [
9]). The case of pleiotropy, or phenotype matching, results in the trait being at migration-selection balance both when preferences are very weak (low
α) and when preferences are very strong (high
α), such that all rare males will be chosen by a matching, rare female (
Figure 1a). However, at intermediate preference strengths (moderate
α), sexual selection, which is positive frequency-dependent, drives further divergence between populations than would exist due to viability selection alone. In contrast, when preferences and traits are on different chromosomes (
Figure 1b), with weak preferences the preference frequencies are homogenized between populations and the trait is at migration-selection balance. With moderate preferences, the preference alleles are still at only slightly divergent frequencies and so exert sexual selection on the traits that draws their frequencies closer together. With strong preferences, preference frequencies are able to build up more divergence, leading to more divergence in trait frequencies as well [
9,
11]. Given that preference divergence is never greater than trait divergence, sexual selection generally tends to homogenize trait frequencies across the populations (at least within
α < 10,000, which would arguably already correspond to separate species).
We next assess the effects of reduced recombination between preferences and traits (
r = 0.001, 0.01, 0.1). With these starting conditions, the results at equilibrium are very similar, to the point of being visually indistinguishable, to those shown in
Figure 1b. In other words, recombination rates as low as
r = 0.001, which would be infeasible to distinguish from pleiotropy using mapping techniques, lead to equilibrium preference and trait frequencies that are far more similar to those of preference and trait loci on separate chromosomes, than they are to the pattern of matching occurring under pleiotropy (
r = 0,
Figure 1a).
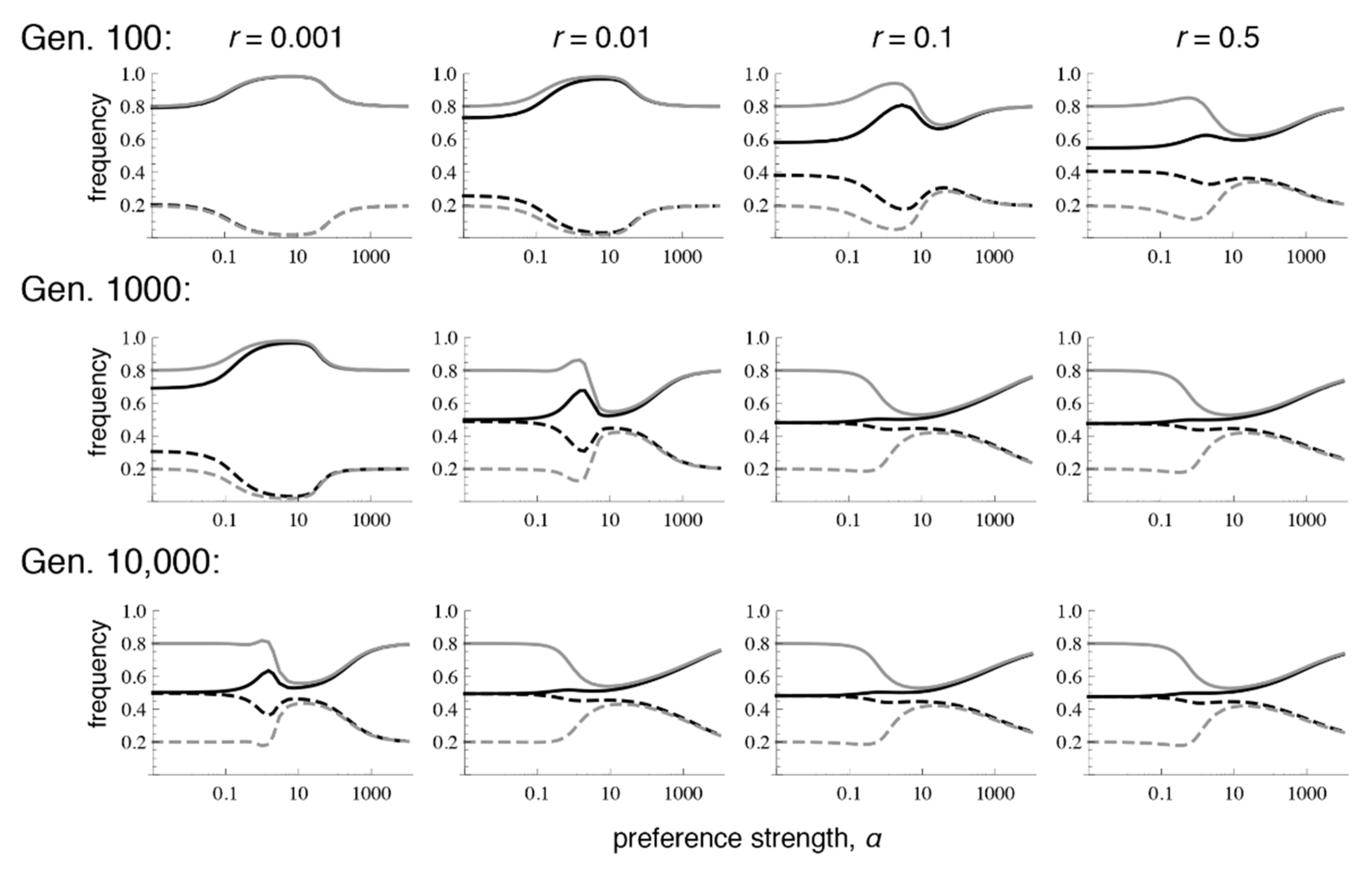
Patterns of differentiation that depend conspicuously on the recombination rate emerge, however, at lower numbers of generations after contact. Each row of
Figure 2 shows the amount of preference and trait divergence that would be observed for a given preference strength if the populations were assessed at a certain number of generations after the onset of migration (assuming the same allele frequencies at secondary contact as in
Figure 1). As can be seen in the figure, when
r = 0.5, the equilibrium pattern is approached relatively quickly. However, with lower recombination rates, the pattern of divergence can look very similar to the pattern expected from phenotype matching for many generations. This can inflate the preference and trait divergence far from that which will be reached at equilibrium, especially with moderate preference strengths, which may be the case most commonly of interest for empirical studies of speciation (i.e., there would be some premating isolation, but it would not be complete). More specifically, at these intermediate preference strengths, because preference divergence can remain inflated for a long time with tight linkage, sexual selection can temporarily allow trait divergence to remain above migration-selection balance. This has the potential to lead to the mistaken impression that sexual selection is “promoting” speciation, whereas when equilibrium is eventually reached, sexual selection will actually have inhibited trait divergence (e.g., the dip in trait frequencies that occurs with intermediate preferences in the bottom row).
Next, we obtain a more thorough picture of the equilibrium conditions by expanding our analysis to consider a broader range of starting conditions, concentrating on the case where population 1 is fixed for T
1 and P
1 at the start of contact, while population 2 is polymorphic, and there is a moderately high strength of mating preference. We have shown elsewhere [
9] that with separate preference and trait loci (and thus
r > 0), symmetric starting conditions between the two populations (i.e.,
p2,1 = 1 −
p2,2,
t2,1 = 1 −
t2,2, and
D1 =
D2) lead to the existence of a stable polymorphic equilibrium that is independent of the recombination rate
r and the starting conditions. With non-symmetric starting conditions, this is not the case.
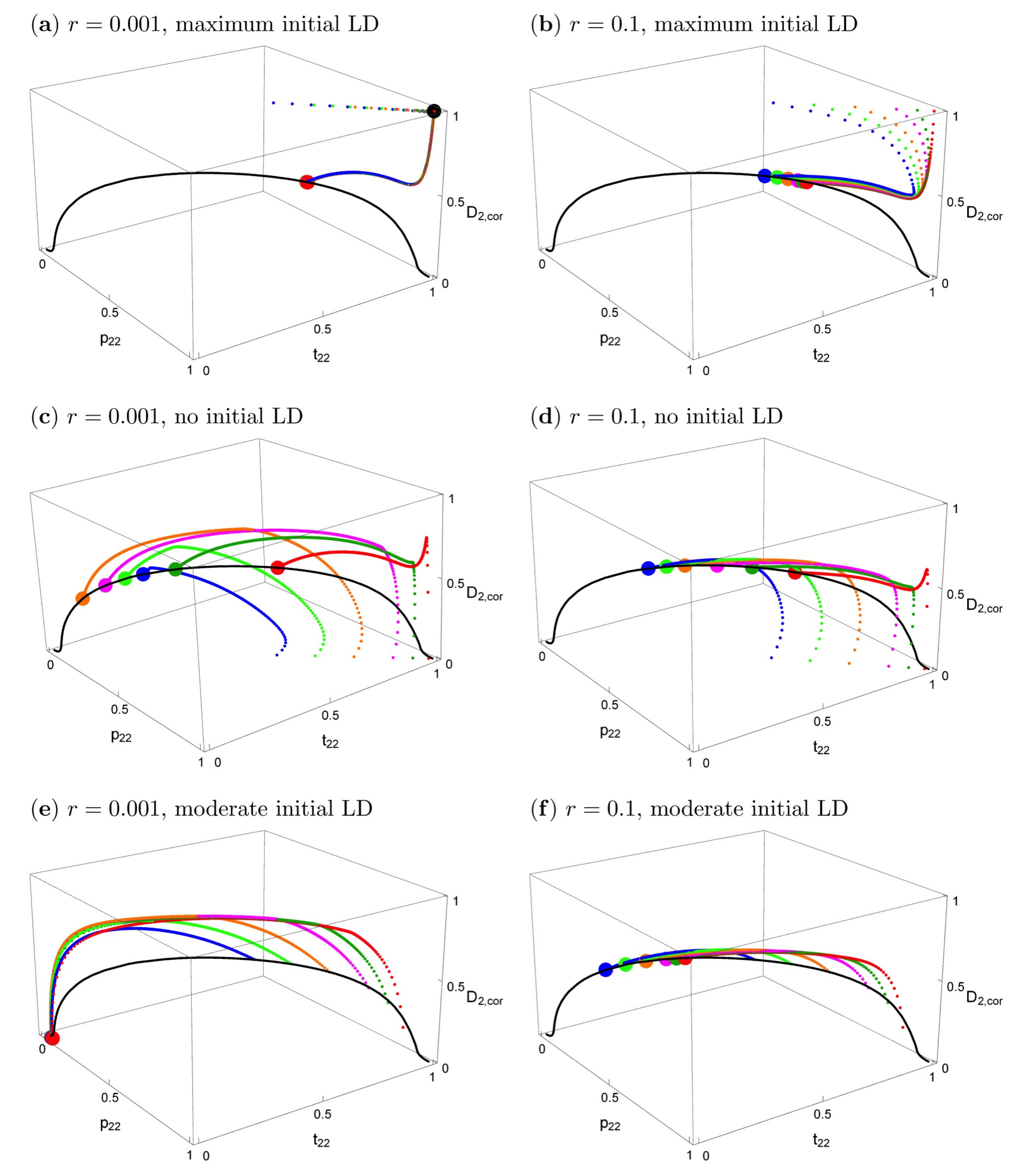
Figure 3 shows the effects of the starting conditions and recombination rate on the evolutionary trajectories. In all panels, the black solid curve represents the stable equilibria. The particular equilibrium point that is reached depends on
r and on the starting conditions, but the curve is the same for every
r > 0. Small deviations from symmetric initial conditions lead to small deviations from the symmetric equilibrium (
Supplementary Materials File S3, Section 3.2), which essentially coincides with the big red dot in
Figure 3a,b, the deviation being too small to be visible.
In the top row of
Figure 3, the starting conditions are consistent with what would be expected under phenotype matching, i.e.,
p2,k =
t2,k,
Dk =
t2,k(1 −
t2,k),
k = 1,2. Therefore, initially, only the two extreme gametes are present (maximum linkage disequilibrium) and the loci are completely correlated.
Figure 3a shows that with tight linkage and after secondary contact, the evolutionary trajectories first approach the phenotype-matching equilibrium (the big black dot) for about 20–100 generations, i.e., divergence between the two population increases. However, in contrast to phenotype matching, or pleiotropy (
r = 0), eventually very slow convergence to a very different equilibrium (the big red dot) occurs (on the order of 10,000–50,000 generations for the given parameters). This equilibrium depends very slightly on initial conditions (hardly visible in this figure because the deviations are so small). A similar evolutionary pattern occurs even for loosely linked loci (
Figure 3b). First, there is again some tendency to increase divergence, but after a few generations, evolution takes a different path and convergence to equilibria on the black curve occurs. It is clearly visible that the final evolutionary outcome with very tightly linked loci (
r = 0.001, red dot in
Figure 3a) is much closer to that for separate preference and trait loci (
r = 0.1, colored dots in
Figure 3b) than to phenotype matching (black dot in
Figure 3a). A detailed time course of a typical evolutionary trajectory with tightly linked loci is shown in
Figure S1a.
The two panels in the second row of
Figure 3 are parallel to those in the top row. All parameters and the initial allele frequencies are the same, but here, linkage disequilibrium is initially absent in population 2 at the time of secondary contact, instead of maximal. All evolutionary trajectories converge to the curve of equilibria. There is not an initial increase of trait divergence in this case (for a detailed time course, see
Figure S1b). Again, the final evolutionary outcome for very tightly linked versus loosely linked loci is quite similar. Interestingly, the latter case leads to somewhat higher equilibrium frequencies of
p2,2 and
t2,2, and also to slightly higher divergence between the two subpopulations (
Supplementary Materials File S3, Section 3.1.2). Although the allele frequencies in the two populations differ from one another and are thus technically “divergent”, it can be seen in
Figure 3c,d that the frequencies of T
2 and P
2 in population 2 are often below 0.5, and in population 1 they are even lower. Therefore, the same alleles (P
1 and T
1) are characteristic of both populations at equilibrium under such starting conditions.
Finally, we considered populations that were at equilibrium when isolated in allopatry. Note that the assumptions of our model lead to a line of polymorphic equilibria for preference and trait frequencies within each allopatric population, as was found in [
23]). For a given equilibrium frequency of an ecologically favored trait T
2 that is prevalent in a population (
), the equilibrium frequency of the preference allele P
2 is lower than that of the trait (
) and is independent of the recombination rate
r (
Supplementary Materials File S3, Section 2.4;
Figure S2; see similar model in [
23]). In other words, there is a curve of polymorphic equilibria in such a population where, counterintuitively, the equilibrium preference frequency
will sometimes be so low that it is below 0.5; this is especially the case when preferences are weak (low
α,
Figure S2). When two populations that are highly divergent in their trait frequencies but are at equilibrium in allopatry contact one another, we observe equilibrium patterns similar to
Figure 1b, but without the misleading initial approach to the phenotype matching pattern that is apparent in
Figure 2 (see
Figure S2).
The two panels in the bottom row of
Figure 3 focus on the case where population 1 is fixed for its characteristic trait and preference, and population 2 is initially at an equilibrium on this curve of allopatric equilibrium values. Otherwise, the two panels are parallel to those in the top and middle row of
Figure 3. The trait and preference alleles eventually reach low frequencies or are lost; an outcome that is determined by the recombination rate (compare
Figure 3e,f). Trait and preference variation is likely to be lost altogether when
r is very low. This shows that the initially more frequent preference allele (across the total of both populations), here P
1, may drag “its” trait allele, here T
1, to fixation.
When populations at equilibrium in allopatry are instead fully symmetrical in their frequencies, we see convergence to the same point regardless of the amount of initial trait divergence, provided that
r > 0 (
Figure S3). Comparison of
Figure 3 with
Figure S3 shows that symmetric initial conditions (as assumed in [
9]) are indeed most conducive to maintaining divergence and local adaptation, i.e., asymmetric initial allele frequencies of the two populations at secondary contact usually lead to reduced divergence and often even to loss of local adaptation in one population, such that maladapted alleles are more common than adaptive ones, or even reach fixation.
With migration rates that are an order of magnitude smaller than above, the evolutionary dynamics are much slower. Both the period of initial approach to the phenotype matching pattern, or with moderate or no initial linkage disequilibrium the period of near constancy of allele frequencies, and the period of final convergence to the true equilibrium, may be elongated by up to an order of magnitude (see
Figure S1c,d and
Figure S4 which is otherwise parallel to
Figure 3).
3.2. Trait Expressed Only in Males
Next, we turn to the case in which the mating trait is expressed only in males. Such traits, which include sexually dimorphic ornaments, courtship song, or pheromones, are more commonly the focus of sexual selection research than are traits expressed in both sexes, and thus may often be the subjects of empirical studies of the effects of sexual selection on speciation. Importantly, even when a trait is only expressed in males, preference/trait pleiotropy is still possible; in
Laupala, for example, there is potential evidence of pleiotropy even though only males produce a song [
17,
18]. Such cases do not fit a literal reading of the term “phenotype matching” because females do not express the phenotype, but they do fit the technical definition of phenotype matching as a category of models because the preference direction and the trait are determined by a single locus.
When the trait is expressed only in males, trait divergence is lost at high preference strengths both during phenotype matching (T
2 is lost across both populations in
Figure 1c), and with the preference and trait on separate chromosomes (the traits have homogenized across populations in
Figure 1d). The loss of one of the alleles with very strong preferences during phenotype matching (
Figure 1c) occurs for the following reason: When selection on the trait only occurs in males, females have less divergent trait frequencies than males at the start of the sexual selection stage of the life cycle (immediately after divergent viability selection). Therefore, females that have very strong preferences for matching male phenotypes will generate sexual selection on males that will tend to initially cause the male traits to converge between the populations. This sexual selection can also lead to a (very slow) loss of polymorphism when frequencies are not sufficiently symmetrical between populations. Note that these effects contrast with those that occur when viability selection acts on both sexes, and when all males, regardless of their identity, have a matching female that strongly prefers them.
Given starting conditions that resemble phenotype matching, separate preference and trait loci with free recombination will homogenize across populations much more so when traits are expressed only in males than when females also express the trait (compare
Figure 1d with
Figure 1b). This occurs because when the trait is expressed in both sexes, females themselves undergo ecological differentiation, increasing trait divergence and subsequently preference divergence (through linkage disequilibrium), right before sexual selection acts in the life cycle (see [
9]). Preferences are thus more homogenized at the time of sexual selection when females do not express the trait, and consequently sexual selection has a more inhibitory effect on trait divergence in this case (note that this is not true with all starting conditions; see below). Although sexual selection does not promote trait differentiation with strong preferences in either the cases of matching or separate chromosomes, this inhibition occurs by different mechanisms (loss of one trait allele versus homogenization of the traits, respectively), so the patterns are still easily distinguishable from each other.
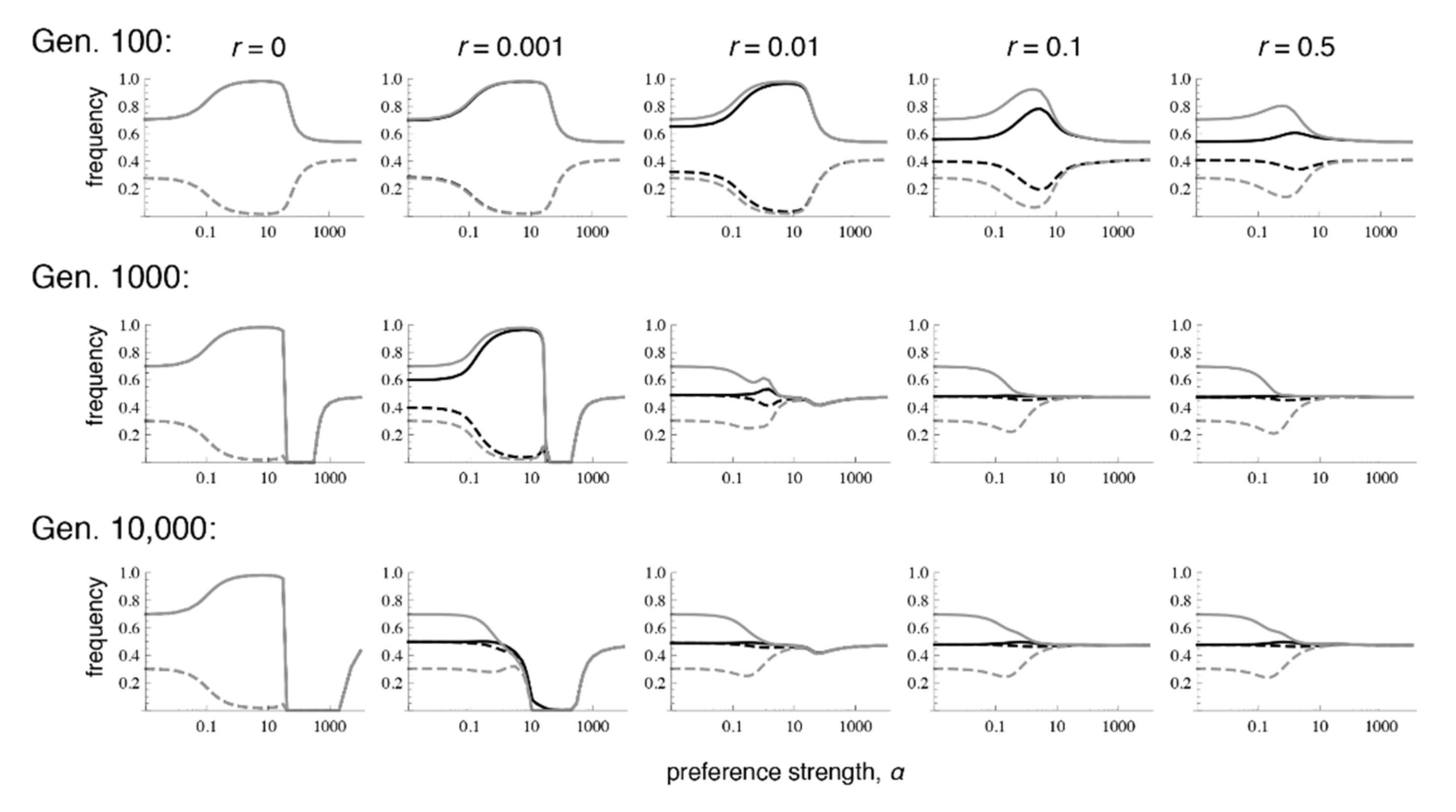
Similar to the case with selection in both sexes, when there is selection only in males, even very low recombination causes the equilibrium values to differ sharply from those that occur with phenotype matching. More specifically, with selection only in males, the polymorphic equilibrium is uniquely determined when
r > 0, and it is the same for every
r > 0 (
Supplementary Materials File S4, Section 2.2;
Figure 4 and
Figure 5; see also [
9]). However, the equilibrium that is reached can differ with the recombination rate and the initial frequencies, in that sometimes loss of trait variation occurs. In particular, we see that with very tight linkage trait variation is sometimes lost with intermediate to high preference strengths (bottom row of
Figure 4, where
r = 0.001;
Figure 5c,e; the pattern in
Figure 4 persists at 1 million generations post-contact). The pattern of trait loss for very tightly linked loci mimics that which is expected from phenotype matching in this range, and apparently occurs because the populations are following the phenotype matching pattern for a long enough period of time to enter this basin of attraction. Looking at patterns of divergence over time, it can again be seen that with low recombination rates, populations can arrive at a pattern of inflated divergence and remain there for some time, especially at intermediate preference strengths (
Figure 4).
We again examined a broader range of starting conditions by focusing in more detail on the case paralleling
Figure 3, of fixation in population 1 with a polymorphism in population 2 (again with
α = 10). Comparison of
Figure 5 with
Figure 3 shows that the short-term behavior of evolutionary trajectories is quite similar, but the long-term behavior differs strongly in most cases. If the trait is expressed only in males, the unique stable polymorphic equilibrium typically exhibits weak differentiation and weak local adaptation, i.e., the frequencies of the locally advantageous trait allele and its preference allele are only slightly above 0.5 (see also bottom row of
Figure 4 if
α > 1). In addition, convergence to the polymorphic equilibrium follows complex trajectories and is typically extremely slow. A typical trajectory is shown in
Figure S1e, in which the different phases are made clearly visible. However, depending on initial conditions and the recombination rate, loss of the trait and preference alleles can also occur. This occurs usually on a much shorter time scale (see also
Figure S1f).
If populations were strongly divergent and in linkage equilibrium before contact, or if they were at the allopatric equilibrium, again there are only two outcomes. We observe either convergence to one and the same polymorphic equilibrium, which maintains weak differentiation, or the loss of a trait allele (
Figure 4,
Figure 5 and
Figure S5). In these cases, there is no initial increase of differentiation (in contrast to the cases when the initial populations had maximum linkage disequilibrium), but the long-term evolution is again complex in many cases (see
Figure 5c–f and
Figure S1e,f) with phases of decreasing and increasing differentiation.
3.3. Asymmetric Migration, Selection and Preference Strength
An asymmetry in the migration rate has the tendency to increase the loss of variation at the trait and preference loci. The case of unidirectional migration (a continent-island model) was analyzed in depth by [
11], for both the phenotype matching model and the case of preferences and traits on separate chromosomes. They found that under phenotype matching, if the trait T
1 is fixed on the continent (our population 1), a polymorphic equilibrium will often exist on the island (our population 2), determined by a balance between viability and sexual selection increasing the frequency of T
2 and migration lowering it. When viability selection is weak, this polymorphic equilibrium representing trait divergence exists only at intermediate preference strengths (see also
Figure S6a or
Figure S7a, far left column). In a preference/trait model, however, Servedio and Bürger [
11] found that maintenance of an island trait is possible only when preferences are sufficiently weak (see also
Figure S6a or
Figure S7a, far right column), unless the preference allele P
2 is also present on the continent.
When examining the case of secondary contact of populations that have almost completely diverged in allopatry (starting with complete linkage disequilibrium) and assessing the role of tight physical linkage between the preference and trait, we find that with unidirectional migration, as with symmetrical migration, the equilibrium trait frequencies on the island resemble those with preferences and traits are on separate chromosomes (both when the trait is expressed only in males and when it is expressed in both sexes (
Figure S6a or
Figure S7a)). Inflated frequencies of the trait T
2 on the island can remain present with tight linkage for long periods of time, but particularly when preferences are very strong, and the trait is expressed in both sexes (
Figures S6a and S8). Strongly asymmetric migration rates, instead of unidirectional rates, exhibit similar findings, with the exception that when the trait is expressed in both sexes, trait and preference divergence can be maintained at very high preference strengths at equilibrium (
Figure S6b vs.
Figure S7b). Exploration of broader starting conditions with moderately strong preferences demonstrates that, as in the case with symmetric migration, high initial linkage disequilibrium is necessary for a temporary inflation of trait divergence to be observed (
Figure S9).
Asymmetry in the strength of preference of P
1 versus P
2 females can also have a strong effect on preference and trait frequencies, and indeed on the ability of divergence to be maintained across populations. The patterns observed, however, are consistent with the ways in which we understand the mechanisms of phenotype matching and separate preferences and trait to work. We examined
α2 >
α1 when preferences and traits started at highly diverged frequencies (both starting in linkage equilibrium and with complete linkage disequilibrium), and examined the cases of selection in both sexes (
Figure S10) and of selection on males alone (
Figure S11). Convergence to equilibrium is slowed considerably when preference strengths are asymmetric, so while we examined the pattern present at one million generations post-contact, this should be understood to be only partway to an equilibrium state.
We find that even when the asymmetry in preference strengths is quite low, such that P2 females have a slightly stronger preference for T2 males than P1 females do for T1 males (e.g., between 1% and 5% stronger with selection in both sexes and <1% stronger with selection only in males), the mating trait T2 can fix across both populations in regions of parameter space and time in which evolution mimics a phenotype matching mechanism with strong sexual selection. This occurs with low recombination and intermediate preference strengths; under these conditions, a higher α2 will favor T2 globally across the two-population system, due to the fact that sexual selection is positive frequency-dependent when the system behaves as under phenotype matching.
On the other hand, the system is expected to behave as under separate preferences and traits when either the recombination rate is high or enough generations have passed under low recombination (see above). Under these conditions, we find that a higher preference of P
2 females for T
2 males tends to paradoxically lead to the loss of T
2 if preferences are strong enough, but only if selection on T
2 occurs in both sexes. A detailed examination of the changes in genotype frequencies through the life cycle shows that when selection occurs in both sexes, this loss of T
2 can be explained by a complex interaction of viability selection in each population with frequency-dependent sexual selection, explained in full in
Supplementary Materials File S5. This explanation hinges in part on the fact that when preferences are more homogenized than traits, which is expected in models with separate preference and trait loci [
9], sexual selection tends to favor the rarer trait allele (though in this case of asymmetric preferences this is tempered by the differences in the preference strengths, see
Supplementary Materials File S5). When selection on the trait only occurs in males, we find that strong but slightly-asymmetric preferences instead lead to the evolution of trait frequencies to a point that is close to that which is found with symmetric preferences (recall that when selection occurs only in males there is a unique equilibrium point instead of the line of equilibria that is found when selection is in both sexes; see above).
As with the case of asymmetric preference strengths, asymmetric selection coefficients between the populations can also lead to some unexpected results. We examined the case where the selection coefficient in population 2 was twice as large as that in population 1 (
Figure S12), again looking at the patterns after 1 million generations. When selection occurs in both sexes, the increase in frequency in the locally adapted trait in females leads, within the viability selection stage of the life cycle, to an increase in the “local” preference due to linkage disequilibrium between the P and T loci (see [
9]). The patterns in this case are relatively straightforward; we see an increase in population 2 of T
2 (and P
2) over the frequencies of T
1 (and P
1) in population 1 (
Figure S12a). When selection occurs only in males, however, we see an unexpected dip in the frequencies of both preferences (P
1 and P
2) in population 2 when preferences are weak to moderate (
Figure S12b).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




