Role of SdiA on Biofilm Formation by Atypical Enteropathogenic Escherichia coli

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Culture Conditions and Plasmids
2.2. Recombinant DNA Techniques
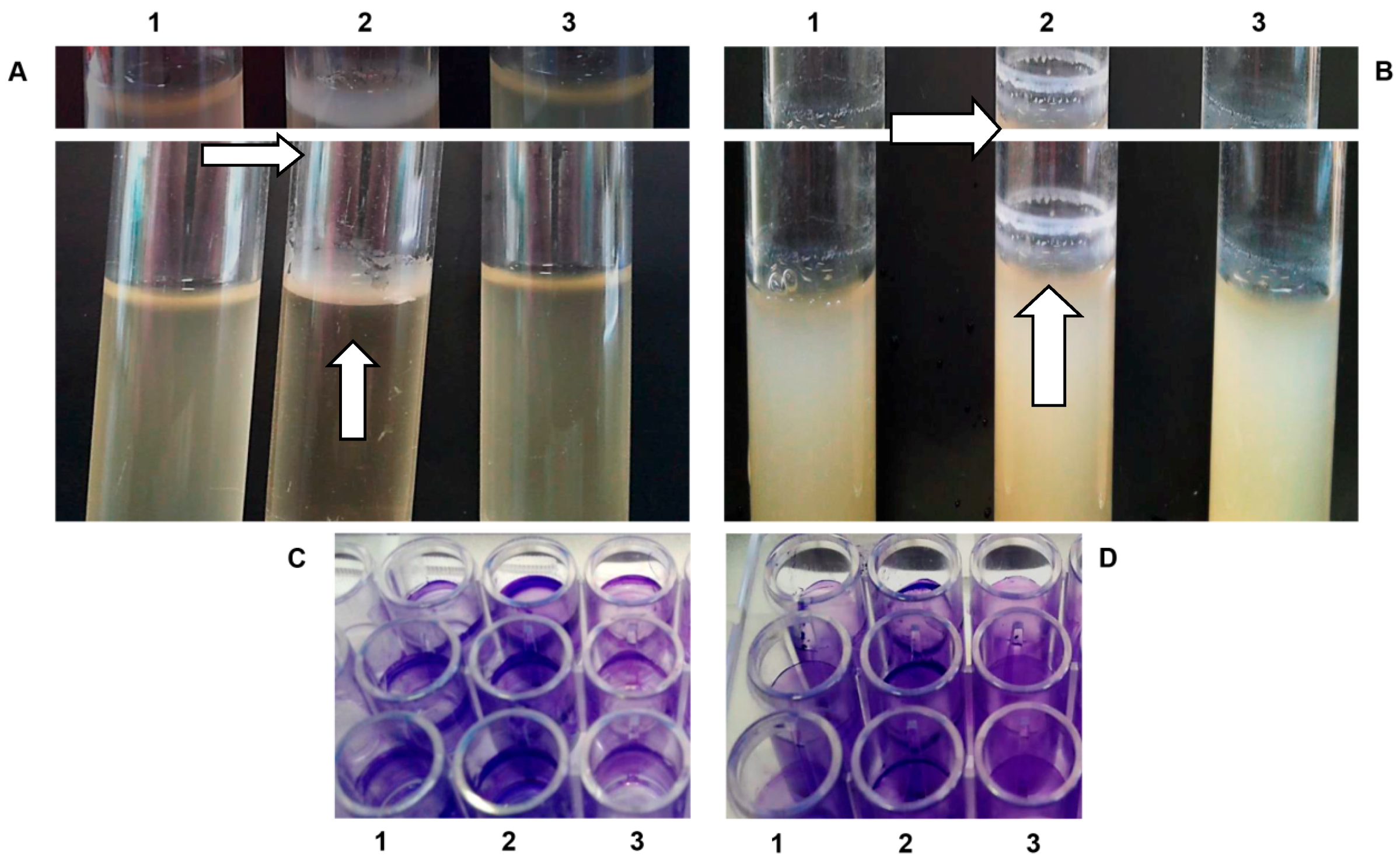
2.3. Congo Red Binding and Cellulose Assay
2.4. Pellicle and Ring-Like Structure Formation
2.5. Biofilm Assay with Crystal Violet
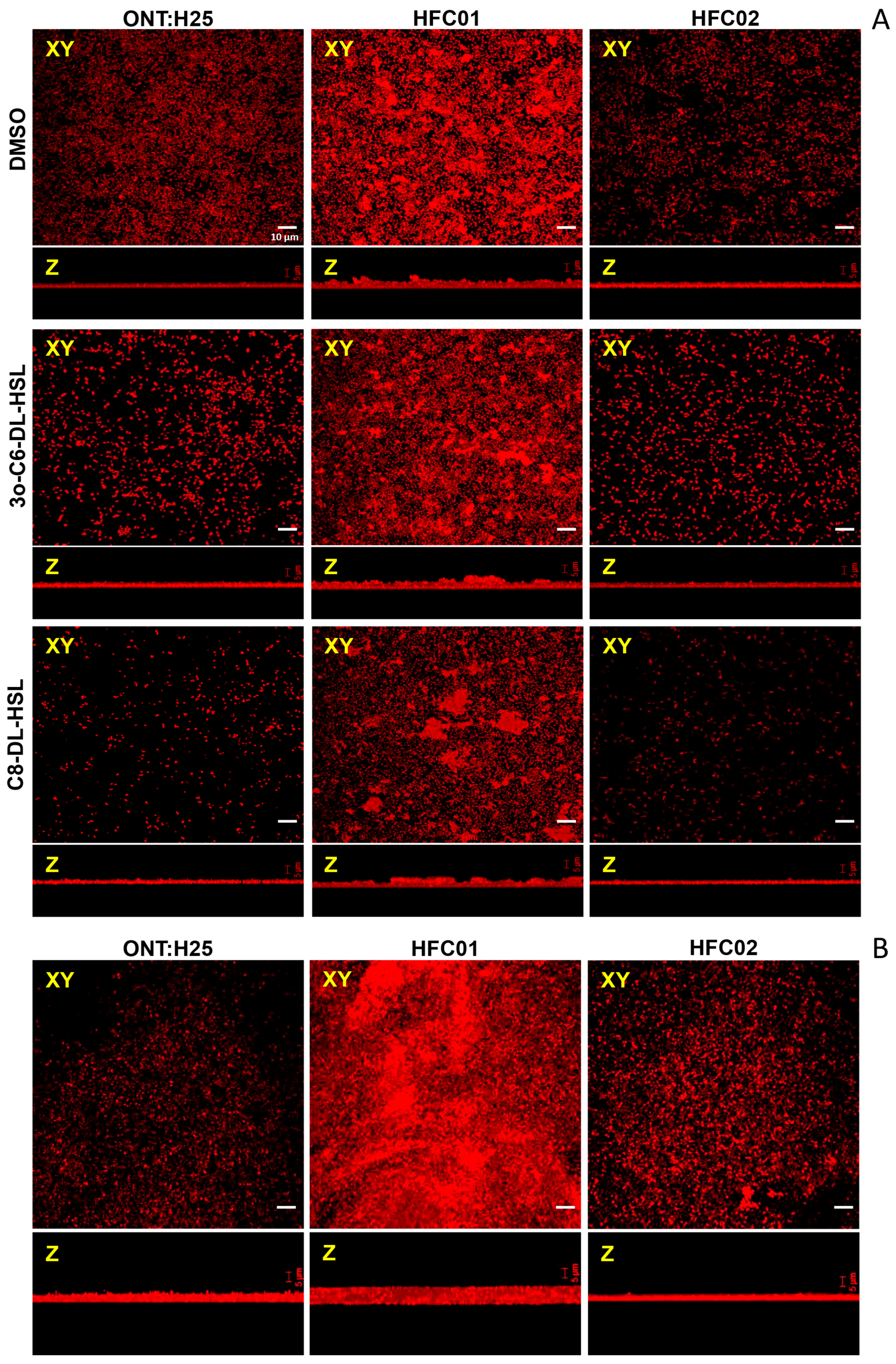
2.6. Confocal Scanning Laser Microscopy
2.7. Fluorescent Actin Staining
2.8. Confocal Scanning Laser Microscopy of Ring-Like Structure in Air-Liquid-Glass Interface
2.9. Motility Assays
2.10. RNA Extraction and Quantitative Real-Time PCR
2.11. Statistical Analysis
3. Results
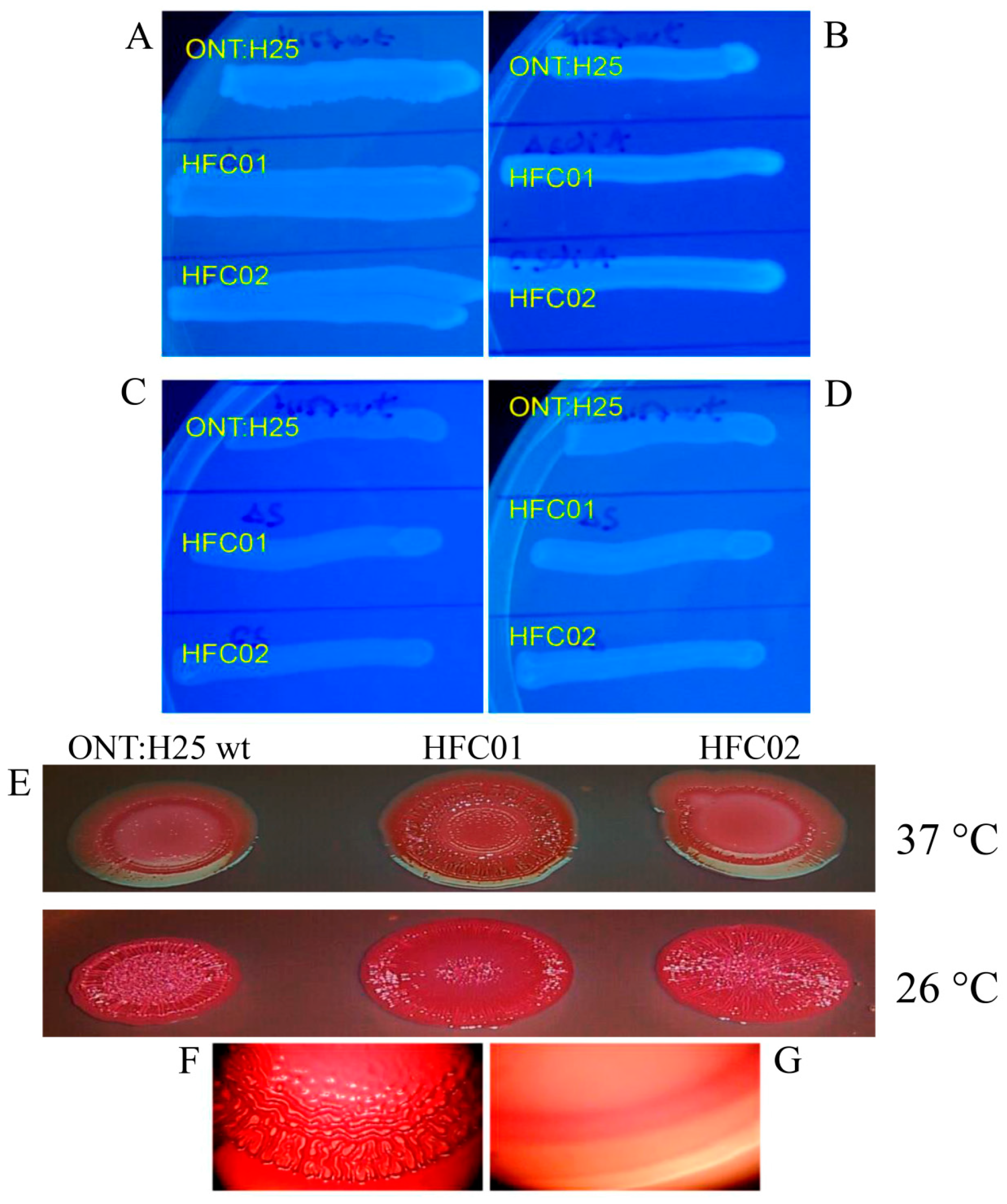
3.1. Deletion of sdiA Alters Curli Expression
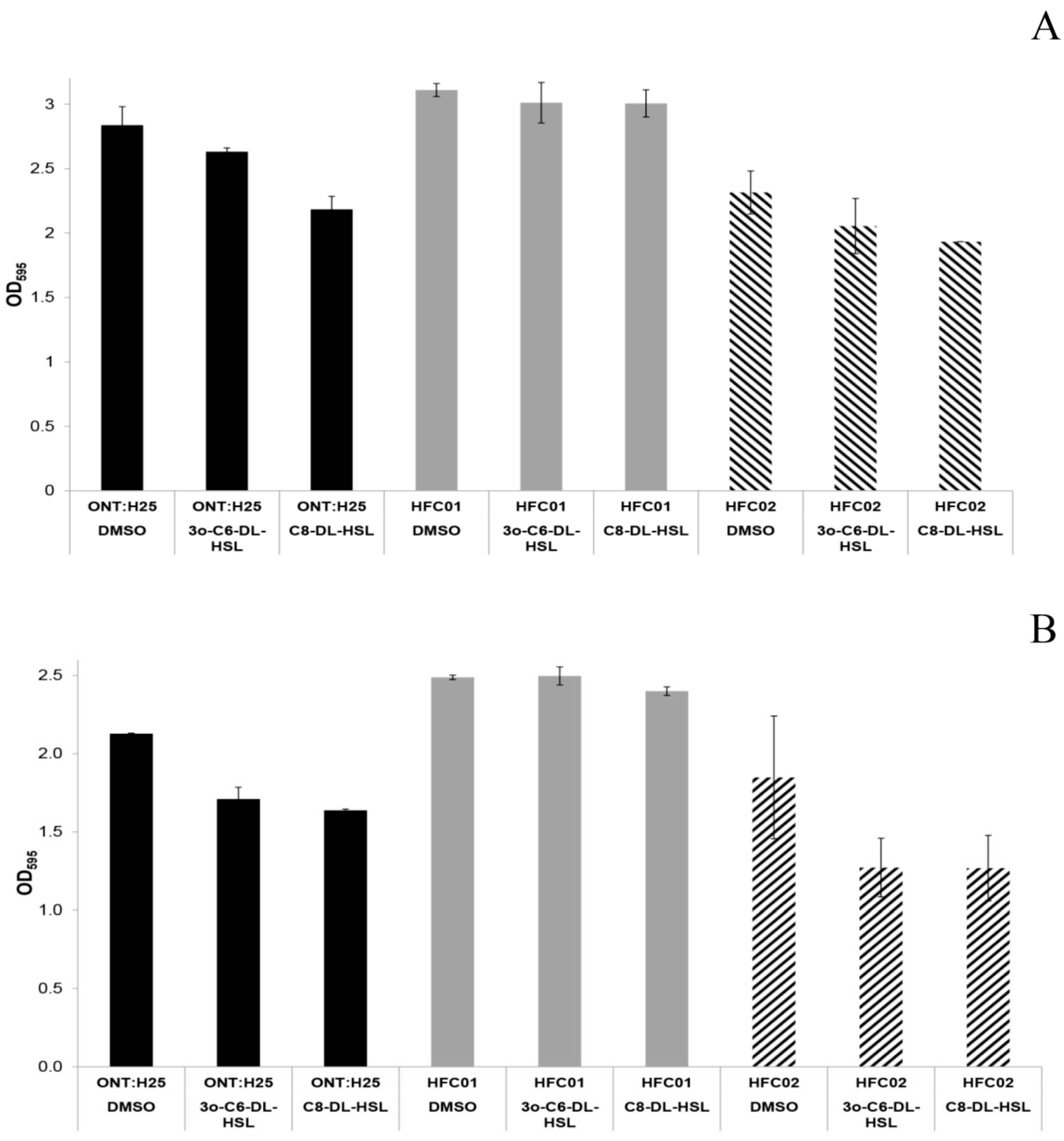
3.2. sdiA Suppresses Biofilm Formation
3.3. Deletion of sdiA Gene Increases Pellicle Formation in Air-Liquid Interface and Ring-Like Structure in Air-Liquid-Glass Interface
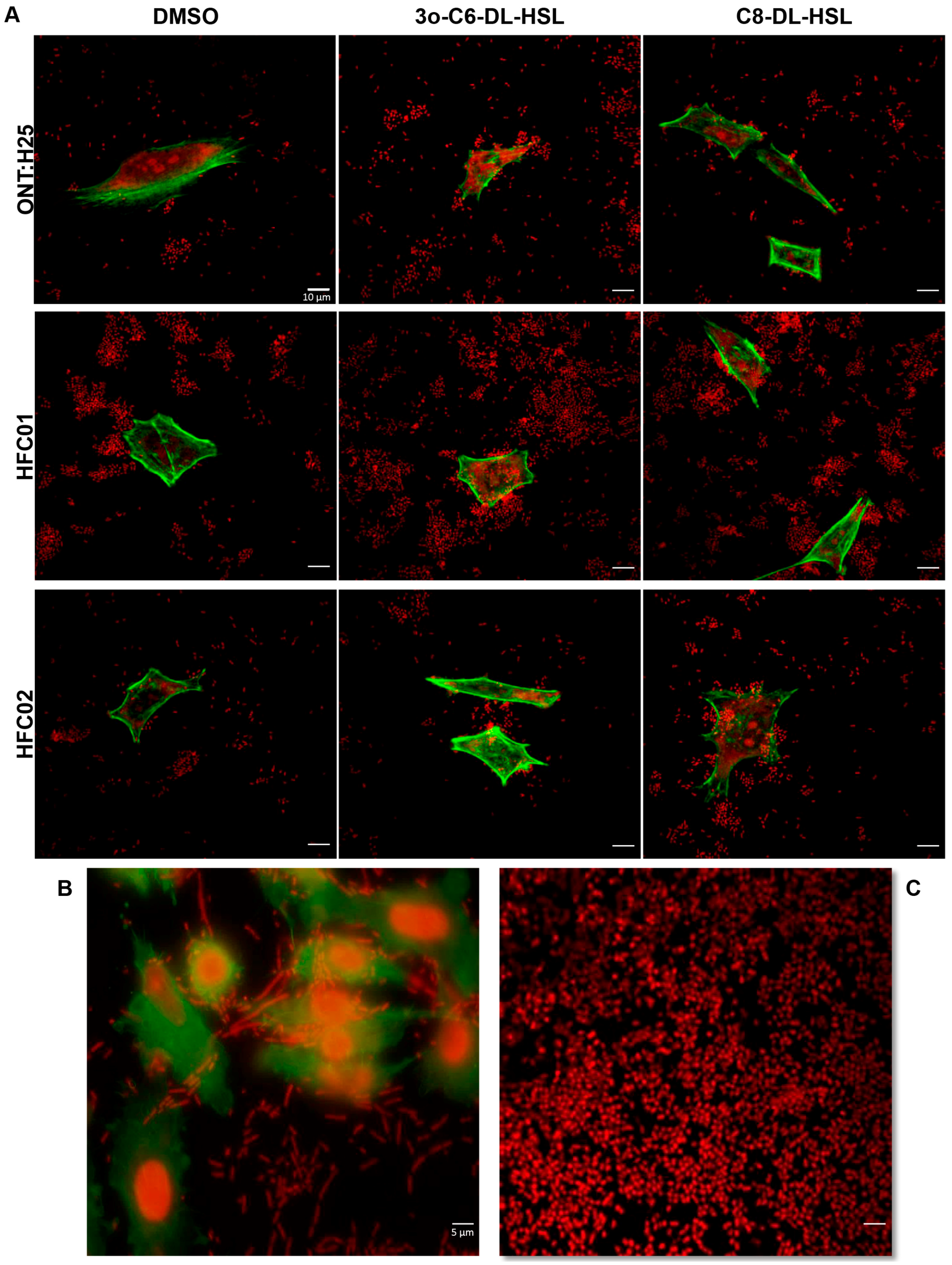
3.4. Presence of sdiA and AHL Result in Different Biofilm Architectures
3.5. AHL Addition Decreases aEPEC Adhesion
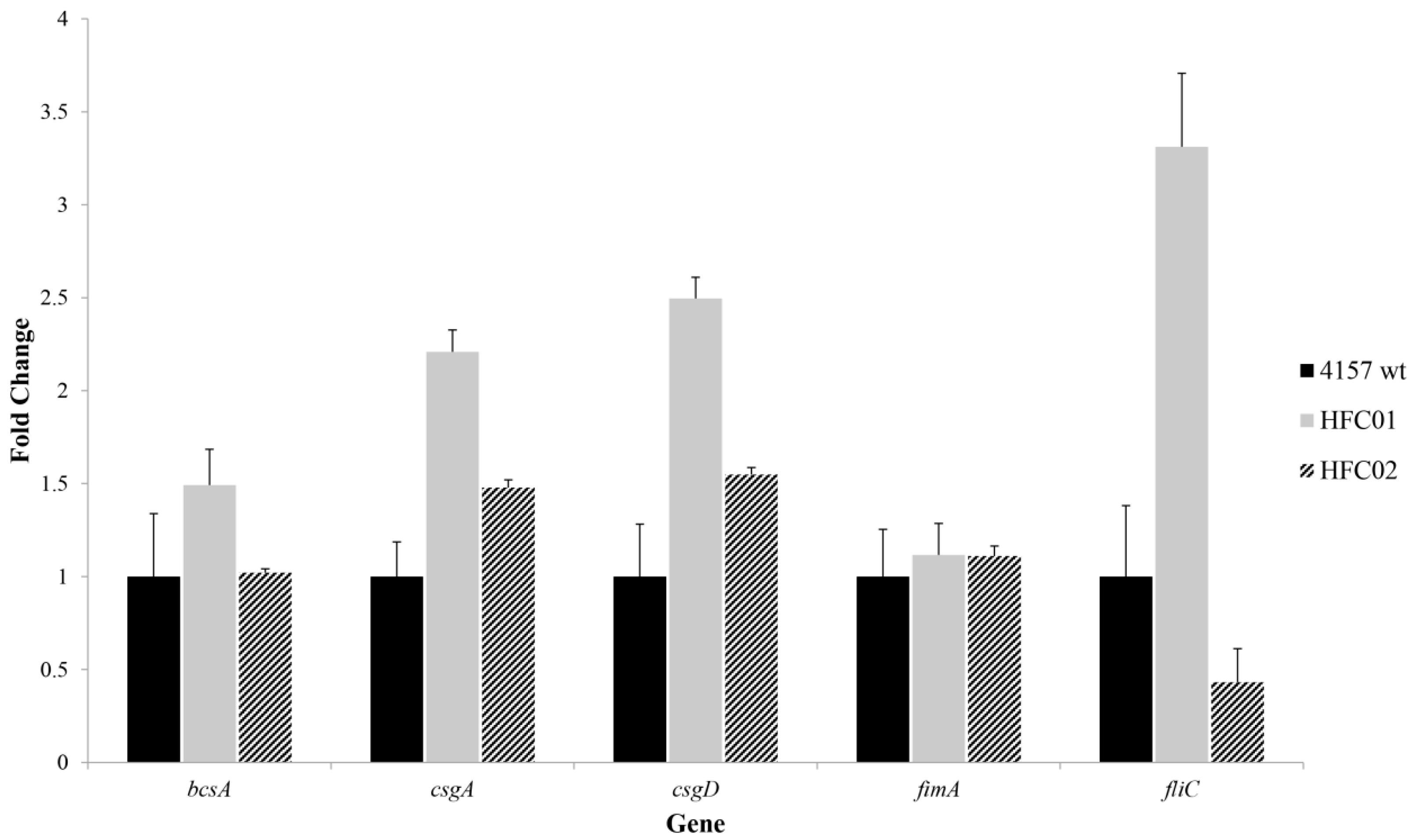
3.6. sdiA Diminishes csgD and csgA Transcription
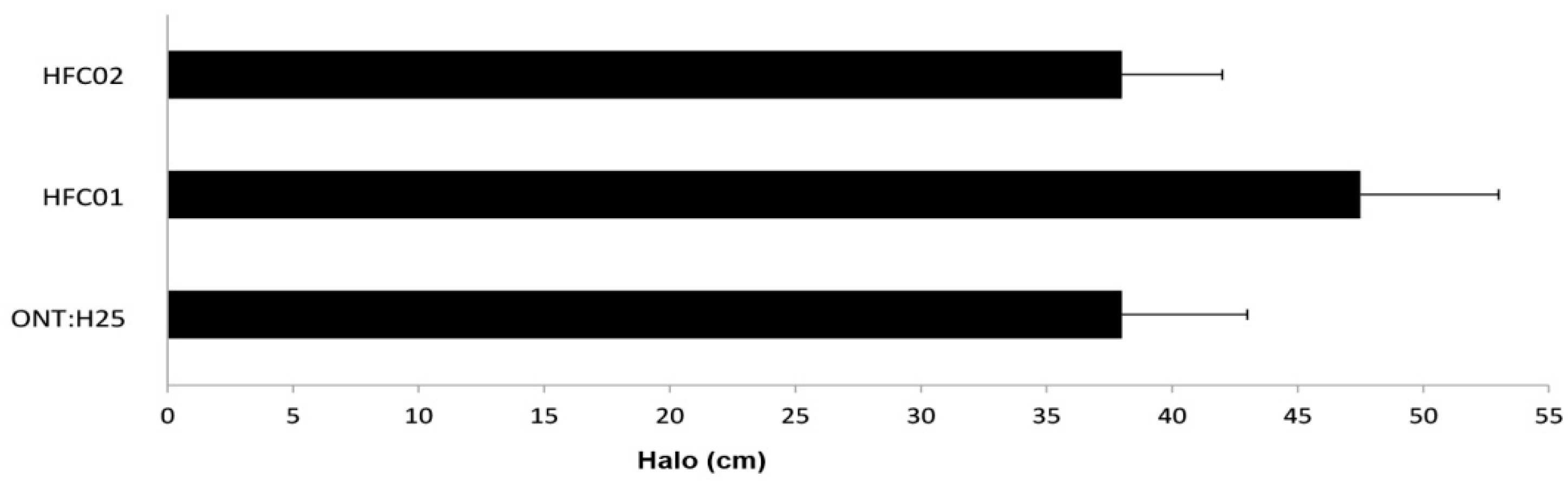
3.7. sdiA Negatively Regulates Motility
4. Discussion
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Wardlaw, T.; Salama, P.; Brocklehurst, C.; Chopra, M.; Mason, E. Diarrhoea: Why children are still dying and what can be done. Lancet 2010, 13, 870–872. [Google Scholar] [CrossRef]
- Ochoa, T.J.; Contreras, C.A. Enteropathogenic Escherichia coli infection in children. Curr. Opin. Infect. Dis. 2011, 24, 478–483. [Google Scholar] [CrossRef] [PubMed]
- Kaper, J.B. Defining enteropathogenic Escherichia coli. Rev. Microbiol. 1996, 27, 130–133. [Google Scholar]
- Hernandes, R.T.; Elias, W.P.; Vieira, M.A.M.; Gomes, T.A.T. An overview of atypical enteropathogenic Escherichia coli. FEMS Microbiol. Lett. 2009, 297, 137–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trabulsi, L.R.; Keller, R.; Gomes, T.A.T. Typical and atypical enteropathogenic Escherichia coli. Emerg. Infect. Dis. 2002, 8, 508–513. [Google Scholar] [CrossRef] [PubMed]
- Knutton, S.; Baldwin, T.; Williams, P.H.; McNeish, A.S. Actin accumulation at sites of bacterial adhesion to tissue culture cells: Basis of a new diagnostic test for enteropathogenic and enterohemorrhagic Escherichia coli. Infect. Immun. 1989, 57, 1290–1298. [Google Scholar] [PubMed]
- Moon, H.W.; Whipp, S.C.; Argenzio, R.A.; Levine, M.M.; Giannella, R.A. Attaching and effacing activities of rabbit and human enteropathogenic Escherichia coli in pig and rabbit intestines. Infect Immun. 1983, 41, 1340–1351. [Google Scholar] [PubMed]
- Ochoa, T.J.; Barletta, F.; Contreras, C.; Mercado, E. New insights into the epidemiology of enteropathogenic Escherichia coli infection. Trans. R. Soc. Trop Med. Hyg. 2008, 102, 852–856. [Google Scholar] [CrossRef] [PubMed]
- Moreira, C.G.; Palmer, K.; Whiteley, M.; Sircili, M.P.; Trabulsi, L.R.; Castro, A.F.P.; Sperandio, V. Bundle-forming pili and EspA are involved in biofilm formation by enteropathogenic Escherichia coli. J. Bacteriol. 2006, 188, 3952–3961. [Google Scholar] [CrossRef] [PubMed]
- Weiss-Muszkat, M.; Shakh, D.; Zhou, Y.; Pinto, R.; Belausov, E.; Chapman, M.R.; Sela, S. Biofilm formation by and multicellular behavior of Escherichia coli O55:H7, an atypical enteropathogenic strain. Appl. Environ. Microb. 2010, 76, 1545–1554. [Google Scholar] [CrossRef] [PubMed]
- Culler, H.F.; Mota, C.M.; Abe, C.M.; Elias, W.P.; Sircili, M.P.; Franzolin, M.R. Atypical enteropathogenic Escherichia coli strains form Biofilm on abiotic surfaces regardless of their adherence pattern on cultured epithelial cells. BioMed. Res. Int. 2014, 2014, 845147:1–845147:10. [Google Scholar] [CrossRef] [PubMed]
- Hernandes, R.T.; Cruz, M.; Yamamoto, D.; Girón, J.A.; Gomes, T.A.T. Dissection of the role of pili and type 2 and 3 secretion systems in adherence and biofilm formation of an atypical enteropathogenic Escherichia coli strain. Infect. Immun. 2013, 81, 3793–3802. [Google Scholar] [CrossRef] [PubMed]
- Prigent-Combaret, C.; Brombacher, E.; Vidal, O.; Ambert, A.; Lejeune, P.; Landini, P.; Dorel, C. complex regulatory network controls initial adhesion and biofilm formation in Escherichia coli via regulation of the csgD Gene. J. Bacteriol. 2001, 183, 7213–7223. [Google Scholar] [CrossRef] [PubMed]
- Zogaj, X.; Bokranz, W.; Nimtz, M.; Römling, U. Production of cellulose and curli fimbriae by members of the family Enterobacteriaceae isolated from the human gastrointestinal tract. Infect. Immun. 2003, 71, 4151–4158. [Google Scholar] [CrossRef] [PubMed]
- Pratt, L.A.; Kolter, R. Genetic analysis of Escherichia coli biofilm formation: Roles of flagella, motility, chemotaxis and type I pili. Mol. Microbiol. 1998, 30, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Prigent-Combaret, C.; Prensier, G.; le Thi, T.T.; Vidal, O.; Lejeune, P.; Dorel, C. Developmental pathway for biofilm formation in curli-producing Escherichia coli strains: Role of flagella, curli and colanic acid. Environ. Microbiol. 2000, 2, 450–464. [Google Scholar] [CrossRef] [PubMed]
- Saldaña, Z.; Xicohtencatl-Cortes, J.; Avelino, F.; Phillips, A.D.; Kaper, J.B.; Puente, J.L.; Girón, J.A. Synergistic role of curli and cellulose in cell adherence and biofilm formation of attaching and effacing Escherichia coli and identification of Fis as a negative regulator of curli. Environ. Microbiol. 2009, 11, 992–1006. [Google Scholar] [CrossRef] [PubMed]
- Zogaj, X.; Nimtz, M.; Rohde, M.; Bokranz, W.; Römling, U. The multicellular morphotypes of Salmonella typhimurium and Escherichia coli produce cellulose as the second component of the extracellular matrix. Mol. Microbiol. 2001, 39, 1452–1463. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, H.H.; Silva, L.E.P.; Souza, R.T.; Silva, N.P.; Scaletsky, I.C.A. Phenotypic and genotypic characteristics associated with biofilm formation in clinical isolates of atypical enteropathogenic Escherichia coli (aEPEC) strains. BMC Microbiol. 2014, 14, 184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antunes, L.C.; Ferreira, R.B.; Buckner, M.M.; Finlay, B.B. Quorum sensing in bacterial virulence. Microbiology 2010, 156, 2271–2282. [Google Scholar] [CrossRef] [PubMed]
- Sircili, M.P.; Walters, M.; Trabulsi, L.R.; Sperandio, V. Modulation of enteropathogenic Escherichia coli virulence by Quorum sensing. Infect. Immun. 2004, 72, 2329–2337. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.N.; Ahmer, B.M.M. Detection of other microbial species by Salmonella: Expression of the SdiA Regulon. J. Bacteriol. 2003, 185, 1357–1366. [Google Scholar] [CrossRef] [PubMed]
- Ahmer, B.M.M. Cell-to-cell signalling in Escherichia coli and Salmonella enterica. Mol. Microbiol. 2004, 52, 933–945. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Martinez-Yamout, M.A.; Dickerson, T.J.; Brogan, A.P.; Wright, P.E.; Dyson, H.J. Structure of the Escherichia coli Quorum sensing protein SdiA: Activation of the folding switch by acyl homoserine lactones. J. Mol. Biol. 2006, 355, 262–273. [Google Scholar] [CrossRef] [PubMed]
- Soares, J.A.; Ahmer, B.M.M. Detection of acyl-homoserine lactones by Escherichia and Salmonella. Curr. Opin. Microbial. 2011, 14, 188–193. [Google Scholar] [CrossRef] [PubMed]
- Shankar, M.; Ponraj, P.; Illakkiam, D.; Rajendhran, J.; Gunasekaran, P. Inactivation of the transcriptional regulator-encoding gene sdiA enhances rice root colonization and biofilm formation in Enterobacter cloacae GS1. J. Bacteriol. 2013, 195, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Maeda, T.; Hong, S.H.; Wood, T.K. Reconfiguring the Quorum-sensing regulator SdiA of Escherichia coli to control biofilm formation via indole and N-acylhomoserine lactones. Appl. Environ. Microb. 2009, 75, 1703–1716. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Jayaraman, A.; Wood, T.K. Indole is an inter-species biofilm signal mediated by SdiA. BMC Microbiol. 2007, 7, 42. [Google Scholar] [CrossRef] [PubMed]
- Houdt, R.; Aertsen, A.; Moons, P.; Vanoirbeek, K.; Michiels, C.W. N-acyl-l-homoserine lactone signal interception by Escherichia coli. FEMS Microbiol. Lett. 2006, 256, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.; Duong, T.; Wu, C.; Choi, J.; Lan, N.; Kang, S.W.; Lokanath, N.K.; Shin, D.; Hwang, H.Y.; Kim, K.K. Structural insights into the molecular mechanism of Escherichia coli SdiA, a Quorum-sensing receptor. Acta Crystallogr. D Biol. Crystallogr. 2014, 70, 694–707. [Google Scholar] [CrossRef] [PubMed]
- Dyszel, J.L.; Soares, J.A.; Swearingen, M.C.; Lindsay, A.; Smith, J.N.; Ahmer, B.M. E. coli K-12 and EHEC genes regulated by SdiA. PLoS ONE 2010, 5, e8946. [Google Scholar] [CrossRef] [PubMed]
- Hughes, D.T.; Terekhova, D.A.; Liou, L.; Hovde, C.J.; Sahl, J.W.; Patankar, A.V.; Gonzalez, J.E.; Edrington, T.S.; Rasko, D.A.; Sperandio, V. Chemical sensing in mammalian host-bacterial commensal associations. Proc. Natl. Acad. Sci. USA 2010, 107, 9831–9836. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, A.; Ahmer, B.M.M. Effect of sdiA on Biosensors of N-acylhomoserine lactones. J. Bacteriol. 2005, 187, 5054–5058. [Google Scholar] [CrossRef] [PubMed]
- Shimada, T.; Shimada, K.; Matsui, M.; Kitai, Y.; Igarashi, J.; Suga, H.; Ishihama, A. Roles of cell division control factor SdiA: Recognition of Quorum sensing signals and modulation of transcription regulation targets. Genes Cells 2014, 19, 405–418. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.K.; Bearson, S.M.D.; Bearson, B.L. Evaluation of the effects of sdiA, a luxR homologue, on adherence and motility of Escherichia coli O157 :H7. Microbiology 2010, 156, 1303–1312. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, Y.; Nguyen, N.X.; Rogers, J.L.; Liao, J.; MacMillan, J.B.; Jiang, Y.; Sperandio, V. Structural and mechanistic roles of novel chemical ligands on the SdiA Quorum-sensing transcription regulator. mBio 2015, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Bueris, V.; Sircili, M.P.; Taddei, C.R.; Santos, M.F.; Franzolin, M.R.; Martinez, M.B.; Ferrer, S.R.; Barreto, M.L.; Trabulsi, L.R. Detection of diarrheagenic Escherichia coli from children with and without diarrhea in Salvador, Bahia, Brazil. Memórias do Instituto Oswaldo Cruz 2007, 102, 839–844. [Google Scholar] [CrossRef] [PubMed]
- Abe, C.M.; Trabulsi, L.R.; Blanco, J.; Blanco, M.; Dahbi, G.; Blanco, J.E.; Mora, A.; Franzolin, M.R.; Taddei, C.R.; Martinez, M.B.; et al. Virulence features of atypical enteropathogenic Escherichia coli identified by the eae+ EAF-negative stx− genetic profile. Diagn. Microbiol. Infect. Dis. 2009, 64, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual; CSHL Press: Cold Spring Harbor, NY, USA, 2001. [Google Scholar]
- Datsenko, K.A.; Wanner, B.L. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. USA 2000, 97, 6640–6645. [Google Scholar] [CrossRef] [PubMed]
- Habdas, B.J.; Smart, J.; Kaper, J.B.; Sperandio, V. The LysR-Type transcriptional regulator QseD alters type three secretion in enterohemorrhagic Escherichia coli and motility in K-12 Escherichia coli. J. Bacteriol. 2010, 192, 3699–3712. [Google Scholar] [CrossRef] [PubMed]
- Njoroge, J.; Sperandio, V. Enterohemorrhagic Escherichia coli virulence regulation by two bacterial adrenergic kinases, QseC and QseE. Infect. Immun. 2012, 80, 688–703. [Google Scholar] [CrossRef] [PubMed]
- Sheikh, J.; Hicks, S.; Dall’Agnol, M.; Phillips, A.D.; Nataro, J.P. Roles for Fis and YafK in biofilm formation by enteroaggregative Escherichia coli. Mol. Microbiol. 2001, 41, 983–997. [Google Scholar] [CrossRef] [PubMed]
- Sperandio, V.; Torres, A.G.; Kaper, J.B. Quorum sensing Escherichia coli regulators B and C (QseBC): A novel two-component regulatory system involved in the regulation of flagella and motility by Quorum sensing in E. coli. Mol. Microbiol. 2002, 43, 809–821. [Google Scholar] [CrossRef] [PubMed]
- Culler, H.F. Envolvimento dos genes qseC e SdiA na formação de biofilme por Escherichia coli enteropatogênica atípica. Ph.D. Thesis, Butantan Institute, University of Campinas, Campinas, SP, Brazil, 2015. [Google Scholar]
- Hedberg, C.W.; Savarino, S.J.; Besser, J.M.; Paulus, C.J.; Thelen, V.M.; Myers, L.J.; Cameron, D.N.; Barrett, T.J.; Kaper, J.B.; Osterholm, M.T. An outbreak of foodborne illness caused by Escherichia coli O39:NM, an agent not fitting into the existing scheme for classifying diarrheogenic E. coli. J. Infect. Dis. 1997, 176, 1625–1628. [Google Scholar] [CrossRef] [PubMed]
- Afset, J.E.; Bevanger, L.; Romundstad, P.; Bergh, K. Association of atypical enteropathogenic Escherichia coli (EPEC) with prolonged diarrhoea. J. Med. Microbiol. 2004, 53, 1137–1144. [Google Scholar] [CrossRef] [PubMed]
- Yatsuyanagi, J.; Saito, S.; Miyajima, Y.; Amano, K.; Enomoto, K. Characterization of atypical enteropathogenic Escherichia coli Strains harboring the astA gene that were associated with a waterborne outbreak of diarrhea in Japan. J. Clin. Microbiol. 2003, 41, 2033–2039. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, R.N.; Taylor, L.S.; Tauschek, M.; Robins-Browne, R.M. Atypical enteropathogenic Escherichia coli infection and prolonged diarrhea in children. Emerg. Infect. Dis. 2006, 12, 597–603. [Google Scholar] [CrossRef] [PubMed]
- Bassler, B.L.; Losick, R. Bacterially Speaking. Cell 2006, 125, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Wang, X.; Weilbacher, T.; Pernestig, A.K.; Melefors, Ö.; Georgellis, D.; Babitzke, P.; Romeo, T. Regulatory circuitry of the CsrA/CsrB and BarA/UvrY systems of Escherichia coli. J. Bacteriol. 2002, 184, 5130–5140. [Google Scholar] [CrossRef] [PubMed]
- Michael, B.; Smith, J.N.; Swift, S.; Heffron, F.; Ahmer, B.M.M. SdiA of Salmonella entericaI is a LuxR homolog that detects mixed microbial communities. J. Bacteriol. 2001, 183, 5733–5742. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, Y.N.; Sheng, H.; Dakarapu, R.; Falck, J.R.; Hovde, C.J.; Sperandio, V. The acyl-homoserine lactone synthase YenI from Yersinia enterocolitica modulates virulence gene expression in enterohemorrhagic Escherichia coli O157:H7. Infect. Immun. 2013, 81, 4192–4199. [Google Scholar] [CrossRef] [PubMed]
- Kanamaru, K.; Kanamaru, K.; Tatsuno, I.; Tobe, T.; Sasakawa, C. SdiA, an Escherichia coli homologue of Quorum-sensing regulators, controls the expression of virulence factors in enterohaemorrhagic Escherichia coli O157:H7. Mol. Microbiol. 2000, 38, 805–816. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains and Plasmids | Description | Reference |
|---|---|---|
| Strains | ||
| aEPEC | aEPEC ONT:H25 | [37] |
| HFC01 | ONT:H25ΔsdiA::kan | Current study |
| HFC02 | HFC01 + pHFC01 | Current study |
| BW 25113 | E. coli K12 strain harboring plasmid pKD46 | [40] |
| BW 25141 | E. coli K12 strain harboring plasmid pKD3 | [40] |
| BW 25141 | E. coli K12 strain harboring plasmid pKD4 | [40] |
| Plasmids | ||
| pHFC01 | sdiA in pBAD/Myc-His A | Current study |
| pKD3 | λ Red template plasmid (Chloramphenicol) | [40] |
| pKD4 | λ Red template plasmid (Kanamycin) | [40] |
| pKD46 | λ Red helper plasmid | [40] |
| Primers | Primer Sequence (5’-3’) | Ref. |
|---|---|---|
| sdiA-DT | F-CAGTAGCGGCCGCGTAACA | Current study |
| R-GAGAATGCGATGGCTTGCAAAAG | ||
| sdiA-DL | F-AGCAACCTGCGTCTTATTCGGTGCATTGATTTTTTTCTGCGTGTAGGCTGGAGCTGCTTC | Current study |
| R-TATCATTATAAATGATACTCACTCTCAGGGGCGTTGCGGTGAACTAAGGAGGATATTCATATG | ||
| pBAD | F-ATGCCATAGCATTTTTATCC | Current study |
| R-GATTTAATCTGTATCAGG | ||
| sdiA-XhoI | F-GTCGCTCGAGAATGCAGGATACGGATTTTTTC | Current study |
| sdiA-EcoRI | R-GTCGGAATTCTCAAATTAAGCCAGTAGCGG | |
| bcsA-RT | F-AGCTCGGCTTCCGTGGC | Current study |
| R-TCATTGTTGAGCCAAAGCCT | ||
| csgA-RT | F-GATGTTGGTCAGGGCTCAG | [35] |
| R-CCACCGAATTGTTTAACTGTC | ||
| csgD-RT | F-GGTAACTATCGTTATAACAGCA | Current study |
| R-TGCCCAGGAAACCGCTTG | ||
| fimA-RT | F-TGTCCCTCAGTTCTACAGCG | [35] |
| R-TCCTAACTGAACGGTTTGATC | ||
| fliC-RT | F-TCCATCGACAAATTCCGTTCT | [41] |
| R-TGGTGACTGCGGAATCCA | ||
| rpoA-RT | F-GCGCTCATCTTCTTCCGAAT | [42] |
| R-CGCGGTCGTGGTTATGTG |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Culler, H.F.; Couto, S.C.F.; Higa, J.S.; Ruiz, R.M.; Yang, M.J.; Bueris, V.; Franzolin, M.R.; Sircili, M.P. Role of SdiA on Biofilm Formation by Atypical Enteropathogenic Escherichia coli. Genes 2018, 9, 253. https://doi.org/10.3390/genes9050253
Culler HF, Couto SCF, Higa JS, Ruiz RM, Yang MJ, Bueris V, Franzolin MR, Sircili MP. Role of SdiA on Biofilm Formation by Atypical Enteropathogenic Escherichia coli. Genes. 2018; 9(5):253. https://doi.org/10.3390/genes9050253
Chicago/Turabian StyleCuller, Hebert F., Samuel C. F. Couto, Juliana S. Higa, Renato M. Ruiz, Min J. Yang, Vanessa Bueris, Marcia R. Franzolin, and Marcelo P. Sircili. 2018. "Role of SdiA on Biofilm Formation by Atypical Enteropathogenic Escherichia coli" Genes 9, no. 5: 253. https://doi.org/10.3390/genes9050253
APA StyleCuller, H. F., Couto, S. C. F., Higa, J. S., Ruiz, R. M., Yang, M. J., Bueris, V., Franzolin, M. R., & Sircili, M. P. (2018). Role of SdiA on Biofilm Formation by Atypical Enteropathogenic Escherichia coli. Genes, 9(5), 253. https://doi.org/10.3390/genes9050253