The Guppy Sex Chromosome System and the Sexually Antagonistic Polymorphism Hypothesis for Y Chromosome Recombination Suppression

Abstract
:1. Introduction
1.1. Sex Chromosomes and Sex Determination
2. Evolution of Separate Sexes and Suppressed Recombination between Sex Determining Factors
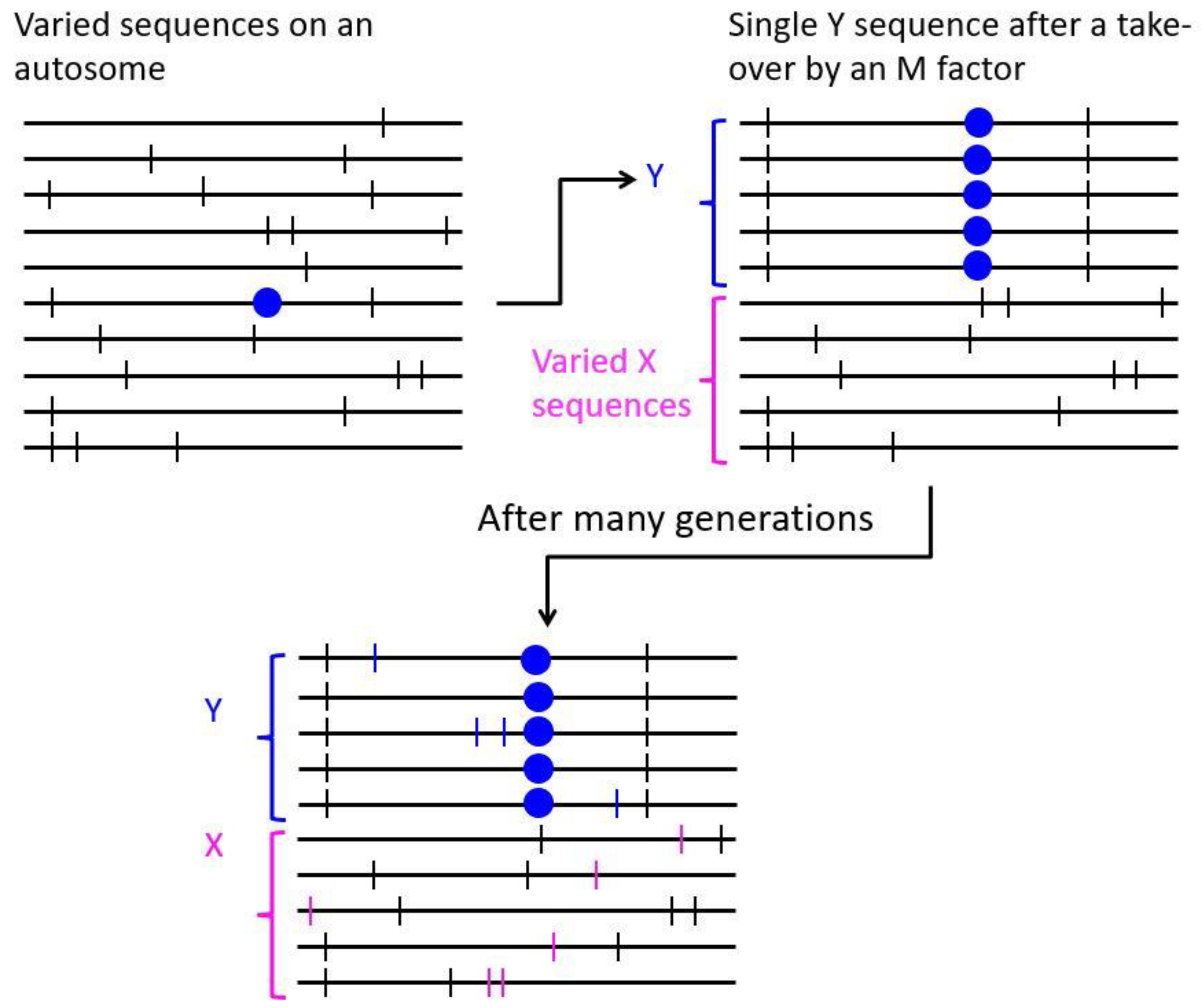
2.1. Turnovers Creating Single Gene Sex Determination
2.2. Control of Secondary Sexual Characters: The Sexually Antagonistic Selection Hypothesis
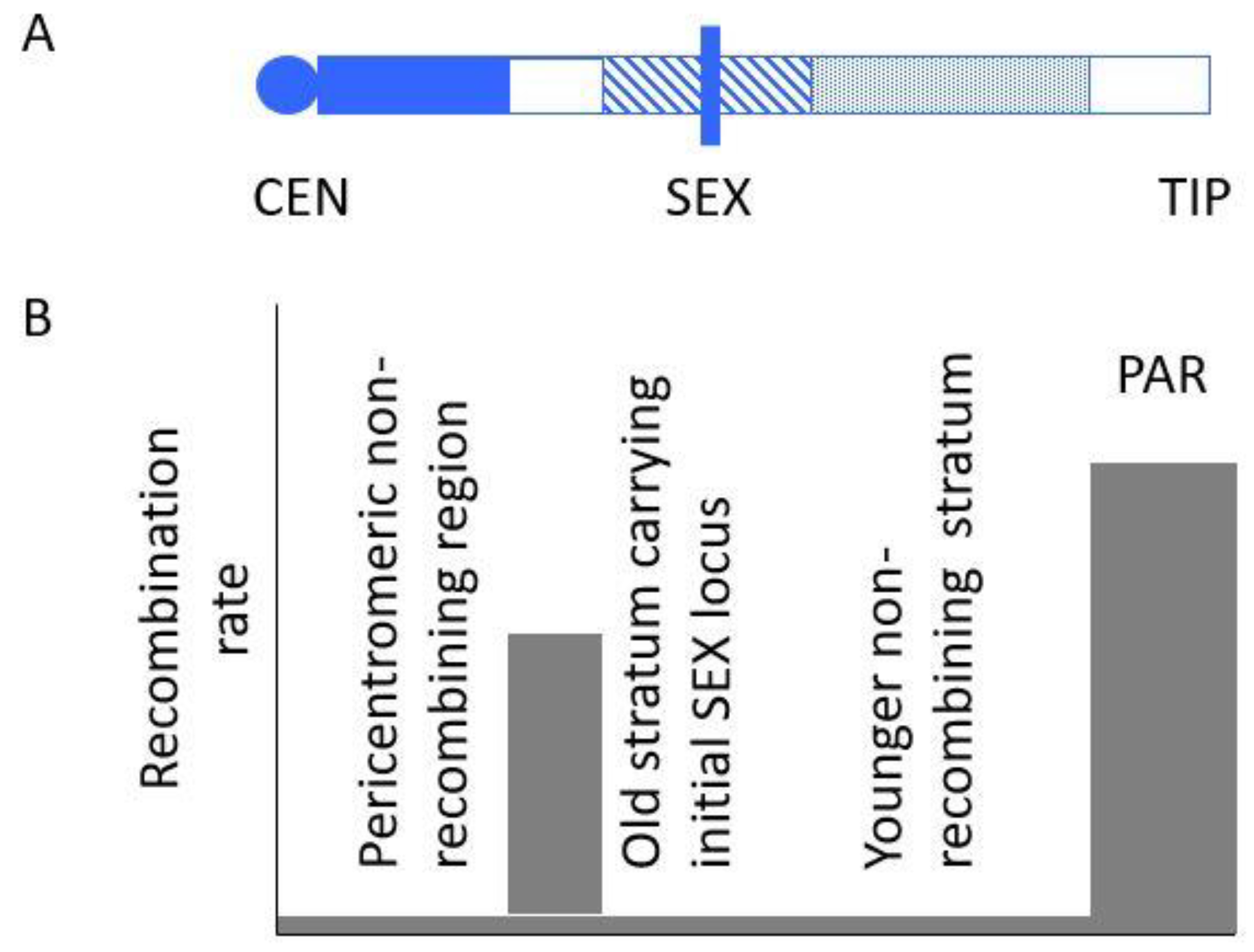
2.3. Evolutionary Strata and Sexually Antagonistic Selection
3. The Guppy System
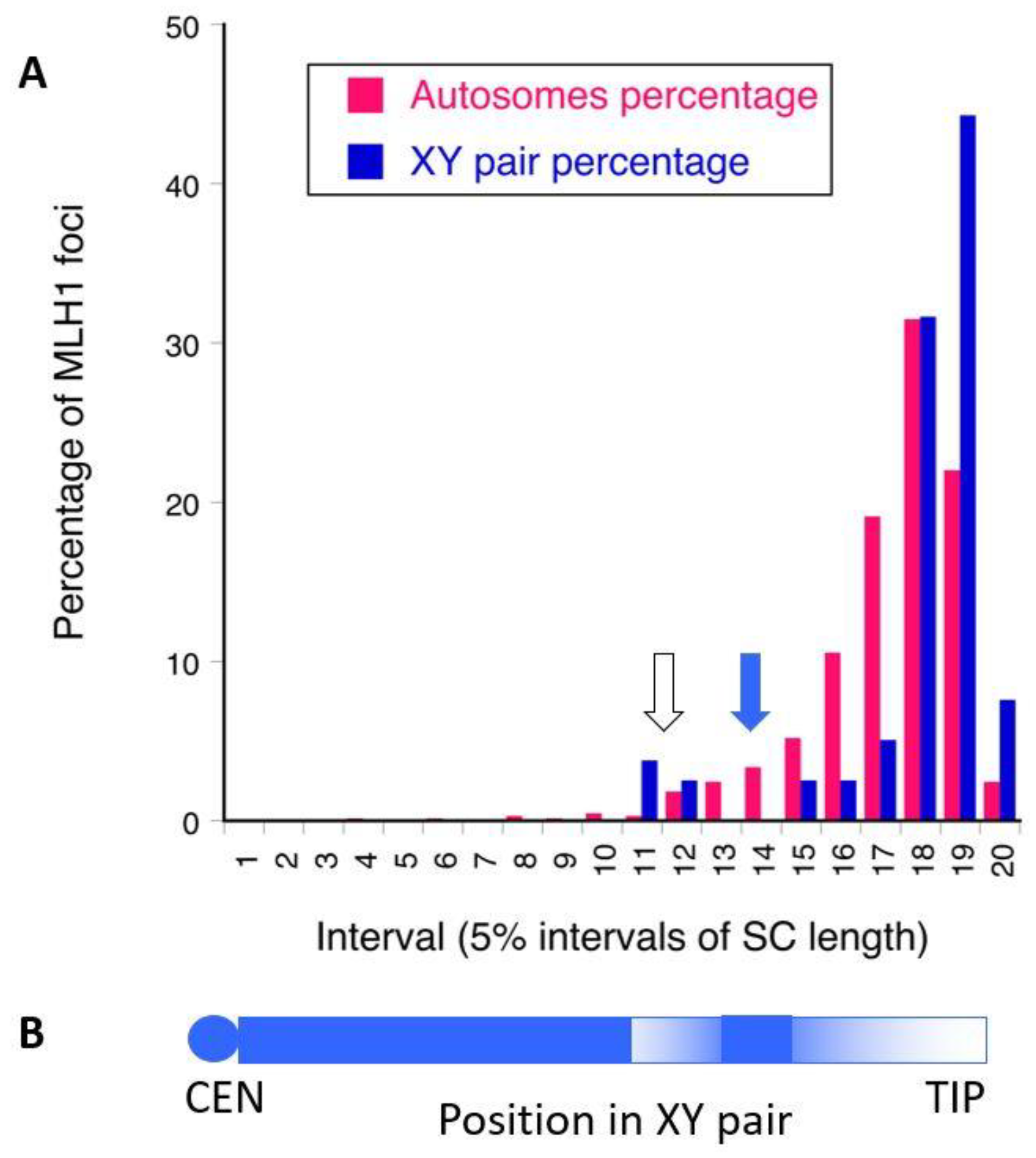
3.1. Recombination Patterns in Fish
3.2. Population Genomic Analysis of the Guppy XY Pair
3.3. Could Linkage Disequilibrium with Sexually Antagonistic Polymorphisms Create the Appearance of Sex Linkage?
3.4. Sex Reversal and the Evolution of Sex Chromosome Sequence Diversity and Divergence
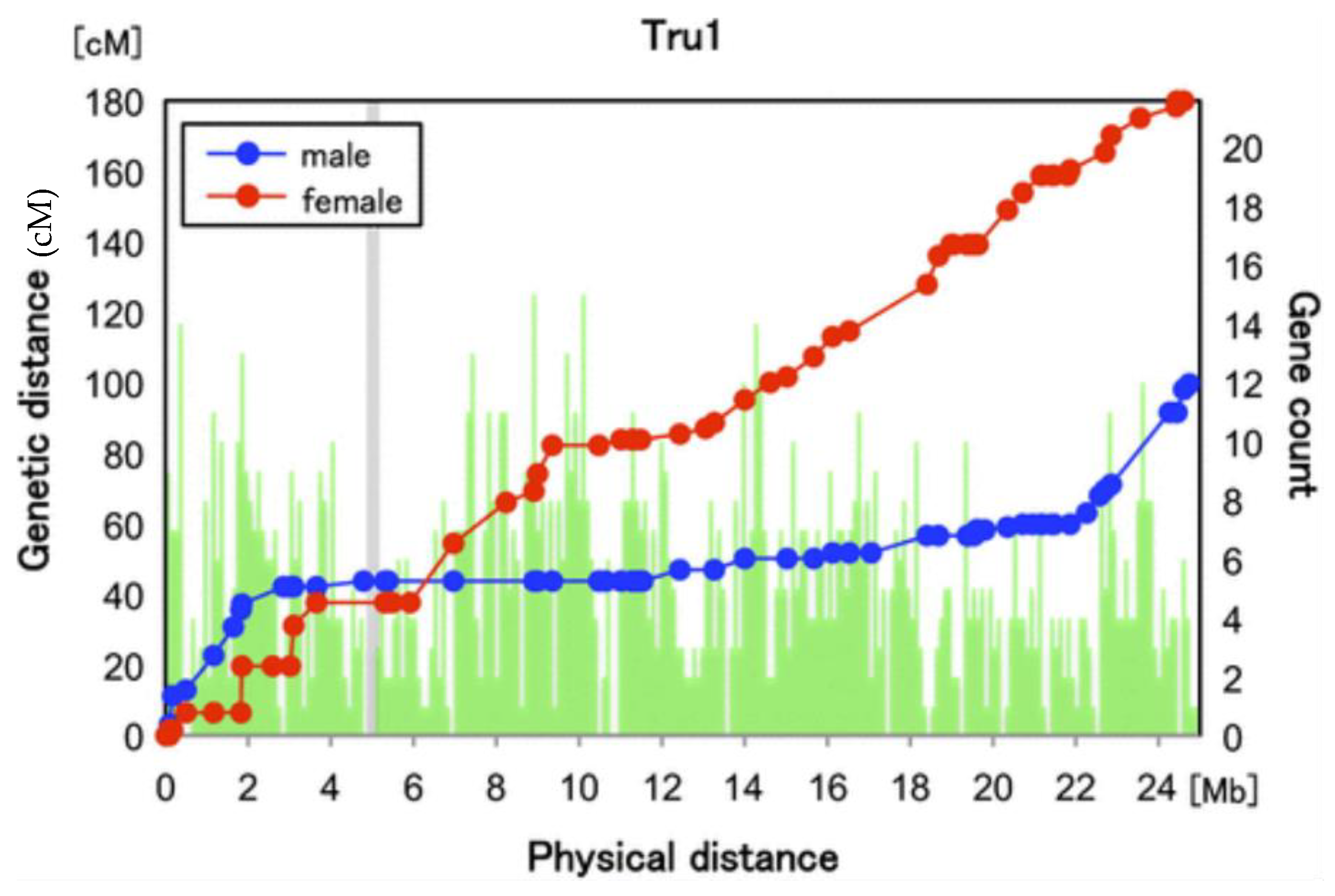
3.5. Evolution of Heterochiasmy
3.6. Male Coloration Factor Genetics and Sex-Limited Expression
Acknowledgments
Conflicts of Interest
References
- Nei, M. Linkage modification and sex difference in recombination. Genetics 1969, 63, 681–699. [Google Scholar] [PubMed]
- Muller, H.J. Some genetic aspects of sex. Am. Nat. 1932, 66, 118–138. [Google Scholar] [CrossRef]
- Westergaard, M. The mechanism of sex determination in dioecious plants. Adv. Genet. 1958, 9, 217–281. [Google Scholar] [PubMed]
- Charlesworth, B.; Charlesworth, D. A model for the evolution of dioecy and gynodioecy. Am. Nat. 1978, 112, 975–997. [Google Scholar] [CrossRef]
- Bull, J.J. Evolution of Sex Determining Mechanisms; Benjamin/Cummings: Menlo Park, CA, USA, 1983. [Google Scholar]
- Wai, C.; Moore, P.H.; Paull, R.E.; Ming, R.; Yu, Q. An integrated cytogenetic and physical map reveals unevenly distributed recombination spots along the papaya sex chromosomes. Chromosome Res. 2012, 20, 753–767. [Google Scholar] [CrossRef] [PubMed]
- Renner, S. Pathways for making unisexual flowers and unisexual plants: Moving beyond the “two mutations linked on one chromosome” model. Am. J. Bot. 2016, 103, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, K.; O’Meally, D.; Azad, B.; Georges, A.; Sarre, S.; Graves, J.; Matsuda, Y.; Ezaz, T. Amplification of microsatellite repeat motifs is associated with the evolutionary differentiation and heterochromatinization of sex chromosomes in Sauropsida. Chromosoma 2016, 125, 111–123. [Google Scholar] [CrossRef] [PubMed]
- Vuilleumier, S.; Lande, R.; van Alphen, J.J.M.; Seehausen, O. Invasion and fixation of sex-reversal genes. J. Evol. Biol. 2007, 20, 913–920. [Google Scholar] [CrossRef] [PubMed]
- Van Doorn, G.; Kirkpatrick, M. Turnover of sex chromosomes induced by sexual conflict. Nature 2007, 449, 909–912. [Google Scholar] [CrossRef] [PubMed]
- Blackmon, H.; Brandvain, Y. Long-term fragility of Y chromosomes is dominated by short-term resolution of sexual antagonism. Genetics 2017, 207, 1621–1629. [Google Scholar] [CrossRef] [PubMed]
- Veller, C.; Muralidhar, P.; Constable, G.W.A.; Nowak, M.A. Drift-induced selection between male and female heterogamety. Genetics 2017, 207, 711–727. [Google Scholar] [CrossRef] [PubMed]
- Bull, J.J.; Charnov, E.L. Changes in the heterogametic mechanism of sex determination. Heredity 1977, 39, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Myosho, T.; Takehana, Y.; Hamaguchi, S.; Sakaizumi, M. Turnover of sex chromosomes in Celebensis group medaka fishes. G3-Genes Genomes Genet. 2015, 5, 2685–2691. [Google Scholar] [CrossRef] [PubMed]
- Myosho, T.; Otake, H.; Masuyama, H.; Matsuda, M.; Kuroki, Y.; Fujiyama, A.; Naruse, K.; Hamaguchi, S.; Sakaizumi, M. Tracing the emergence of a novel sex-determining gene in medaka, Oryzias luzonensis. Genetics 2012, 191, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Ieda, R.; Hosoya, S.; Tajima, S.; Atsumi, K.; Kamiya, T.; Nozawa, A.; Aoki, Y.; Tasumi, S.; Koyama, T.; Nakamura, O.; et al. Identification of the sex-determining locus in grass puffer (Takifugu niphobles) provides evidence for sex-chromosome turnover in a subset of Takifugu species. PLoS ONE 2018, 13, e0190635. [Google Scholar] [CrossRef] [PubMed]
- Quinn, A.; Sarre, S.; Ezaz, T.; Graves, J.A.M.; Georges, A. Evolutionary transitions between mechanisms of sex determination in vertebrates. Biol. Lett. 2011, 7, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Vicoso, B.; Bachtrog, D. Numerous transitions of sex chromosomes in Diptera. PLoS Biol. 2015, 13, e1002078. [Google Scholar] [CrossRef] [PubMed]
- Connallon, T.; Cox, R.M.; Calsbeek, R. Fitness consequences of sex-specific selection. Evolution 2010, 64, 1671–1682. [Google Scholar] [CrossRef] [PubMed]
- Delph, L.F.; Knapczyk, F.; Taylor, D. Among-population variation and correlations in sexually dimorphic traits of Silene latifolia. J. Evol. Biol. 2002, 15, 1011–1020. [Google Scholar] [CrossRef]
- Grath, S.; Parsch, J. Sex-biased gene expression. Ann. Rev. Genet. 2016, 50, 29–44. [Google Scholar] [CrossRef] [PubMed]
- Ridout, K.E.; Veltsos, P.; Muyle, A.; Emery, O.; Rastas, P.; Marais, G.A.B.; Filatov, D.A.; Pannell, J.R. Hallmarks of early sex-chromosome evolution in the dioecious plant Mercurialis annua revealed by de novo genome assembly, genetic mapping and transcriptome analysis. BioArxiv 2017. [Google Scholar] [CrossRef]
- Carlson, C.; Choi, Y.; Chan, A.P.; Serapiglia, M.J.; Town, C.D.; Smart, L.B. Dominance and sexual dimorphism pervade the Salix purpurea L. transcriptome. Genome Biol. Evol. 2017, 9, 2377–2394. [Google Scholar] [CrossRef] [PubMed]
- Zemp, N.; Tavares, R.; Muyle, A.; Charlesworth, D.; Marais, G.; Widmer, A. Evolution of sex-biased gene expression in a dioecious plant. Nat. Plants 2016, 2, 16168. [Google Scholar] [CrossRef] [PubMed]
- Fry, J.D. The genomic location of sexually antagonistic variation: Some cautionary comments. Evolution 2010, 64, 1510–1516. [Google Scholar] [CrossRef] [PubMed]
- Bachtrog, D. A dynamic view of sex chromosome evolution. Curr. Opin. Genet. Dev. 2006, 16, 578–585. [Google Scholar] [CrossRef] [PubMed]
- Connallon, T.; Clark, A.G. Evolutionary inevitability of sexual antagonism. Proc. R. Soc. B Biol. Sci. 2014, 281, 20132123. [Google Scholar] [CrossRef] [PubMed]
- Jordan, C.; Charlesworth, D. The potential for sexually antagonistic polymorphism in different genome regions. Evolution 2012, 66, 505–516. [Google Scholar] [CrossRef] [PubMed]
- Vicoso, B.; Charlesworth, B. Evolution on the X chromosome: Unusual patterns and processes. Nat. Rev. Genet. 2006, 7, 645–653. [Google Scholar] [CrossRef] [PubMed]
- Cavoto, E.; Neuenschwander, S.; Goudet, J.; Perrin, N. Sex-antagonistic genes, XY recombination and feminized Y chromosomes. J. Evol. Biol. 2018, 31, 416–427. [Google Scholar] [CrossRef] [PubMed]
- Ironside, J. No amicable divorce? Challenging the notion that sexual antagonism drives sex chromosome evolution. Bioessays 2010, 32, 718–726. [Google Scholar] [CrossRef] [PubMed]
- Charlesworth, D.; Charlesworth, B.; Marais, G. Steps in the evolution of heteromorphic sex chromosomes. Heredity 2005, 95, 118–128. [Google Scholar] [CrossRef] [PubMed]
- Lahn, B.T.; Page, D.C. Four evolutionary strata on the human X chromosome. Science 1999, 286, 964–967. [Google Scholar] [CrossRef] [PubMed]
- Skaletsky, H.; Kuroda-Kawaguchi, T.; Minx, P.J.; Cordum, H.S.; Hillier, L.; Brown, L.G.; Repping, S.; Pyntikova, T.; Ali, J.; Bieri, T.; et al. The male-specific region of the human Y chromosome is a mosaic of discrete sequence classes. Nature 2003, 423, 825–837. [Google Scholar] [CrossRef] [PubMed]
- Bellott, D.; Hughes, J.F.; Skaletsky, H.; Brown, L.G.; Pyntikova, T.; Cho, T.; Koutseva, N.; Zaghlul, S.; Graves, T.; Rock, S.; et al. Mammalian Y chromosomes retain widely expressed dosage-sensitive regulators. Nature 2014, 508, 494–499. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhang, J.; Yang, W.; An, N.; Zhang, P.; Zhang, G.; Zhou, Q. Temporal genomic evolution of bird sex chromosomes. BMC Evol. Biol. 2014, 14, 250. [Google Scholar] [CrossRef] [PubMed]
- White, M.; Kitano, J.; Peichel, C. Purifying selection maintains dosage-sensitive genes during degeneration of the threespine stickleback Y chromosome. Mol. Biol. Evol. 2015, 32, 1981–1995. [Google Scholar] [CrossRef] [PubMed]
- Bergero, R.; Forrest, A.; Kamau, E.; Charlesworth, D. Evolutionary strata on the X chromosomes of the dioecious plant Silene latifolia: Evidence from new sex-linked genes. Genetics 2007, 175, 1945–1954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reichwald, K.; Petzold, A.; Koch, P.; Downie, B.; Hartmann, N.; Pietsch, S.; Baumgart, M.; Chalopin, D.; Felder, M.; Bens, M.; et al. Insights into sex chromosome evolution and aging from the genome of a short-lived fish. Cell 2015, 163, 1527–1538. [Google Scholar] [CrossRef] [PubMed]
- Lemaitre, C.; Braga, M.D.V.; Gautier, C.; Sagot, M.-F.; Tannier, E.; Marais, G.A.B. Footprints of inversions at present and past pseudoautosomal boundaries in human sex chromosomes. Genome Biol. Evol. 2009, 1, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Laere, A.-S.V.; Coppieters, W.; Georges, M. Characterization of the bovine pseudoautosomal boundary: Documenting the evolutionary history of mammalian sex chromosomes. Genome Res. 2008, 18, 1884–1895. [Google Scholar] [CrossRef] [PubMed]
- White, M.A.; Ikeda, A.; Payseur, B.A. A pronounced evolutionary shift of the pseudoautosomal region boundary in house mice. Mammalian Genome 2012, 23, 454–466. [Google Scholar] [CrossRef] [PubMed]
- Dumont, B.L. Variation and evolution of the meiotic requirement for crossing over in mammals. Genetics 2017, 205, 155–168. [Google Scholar] [CrossRef] [PubMed]
- Haskins, C.; Haskins, E.F.; McLaughlin, J.; Hewitt, R.E. Polymorphisms and population structure in Lebistes reticulatus, an ecological study. In Vertebrate Speciation; Blair, W.F., Ed.; University of Texas Press: Austin, TX, USA, 1961; pp. 320–395. [Google Scholar]
- Endler, J.A. Natural selection on color patterns in Poecilia reticulata. Evolution 1980, 34, 76–91. [Google Scholar] [CrossRef] [PubMed]
- Hughes, K.; Houde, A.; Price, A.; Rodd, F. Mating advantage for rare males in wild guppy populations. Nature 2013, 503, 108–110. [Google Scholar] [CrossRef] [PubMed]
- Olendorf, R.; Rodd, F.; Punzalan, D.; Houde, A.; Reznick, D.; Hughes, K. Frequency-dependent survival in natural guppy populations. Nature 2006, 441, 633–636. [Google Scholar] [CrossRef] [PubMed]
- Magurran, A.E. Evolutionary Ecology: The Trinidadian Guppy; Oxford University Press: Oxford, UK, 2005. [Google Scholar]
- Winge, O. The location of eighteen genes in Lebistes reticulatus. J. Genet. 1927, 18, 201–217. [Google Scholar] [CrossRef]
- Winge, O.; Ditlevsen, E. Colour inheritance and sex determination in Lebistes. Heredity 1947, 1, 65–83. [Google Scholar] [CrossRef]
- Bachtrog, D. Y-chromosome evolution: Emerging insights into processes of Y-chromosome degeneration. Nat. Rev. Genet. 2012, 14, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Lindholm, A.; Breden, F. Sex chromosomes and sexual selection in Poeciliid fishes. Am. Nat. 2002, 160, S214–S224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Künstner, A.; Hoffmann, M.; Fraser, B.A.; Kottler, V.A.; Sharma, E.; Weigel, D.; Dreyer, C. The genome of the Trinidadian guppy, Poecilia reticulata, and variation in the Guanapo population. PLoS ONE 2017, 11, e0169087. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.P.; López-Sepulcre, A.; Reznick, D.N. Predation-associated differences in sex-linkage of wild guppy coloration. Evolution 2012, 66, 912–918. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.P.; López-Sepulcre, A.; Rumbo, D.; Reznick, D.N. Rapid changes in the sex linkage of male coloration in introduced guppy populations. Am. Nat. 2017. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, J.; Carmo, A.; Rosse, I.; Vimieira, A.; Ludwig, S.; Fachin, S.; Pereira, A.; Brandão-Dias, P.; Abreu, N.; Kalapothakis, E. High-Throughput Sequencing Strategy for Microsatellite Genotyping Using Neotropical Fish as a Model. Front. Genet. 2018. [Google Scholar] [CrossRef] [PubMed]
- Natri, H.M.; Shikano, T.; Merilä, J. Progressive recombination suppression and differentiation in recently evolved neo-sex chromosomes. Mol. Biol. Evol. 2013, 30, 1131–1144. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, N.; Hoffmann, M.; Willing, E.; Lanz, C.; Weigel, D.; Dreyer, C. Genetic linkage map of the guppy, Poecilia reticulata, and quantitative trait loci analysis of male size and colour variation. Proc. R. Soc. B 2009, 276, 2195–2208. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, N.; Hoffmann, M.; Weigel, D.; Dreyer, C. Linkage analysis reveals the independent origin of Poeciliid sex chromosomes and a case of atypical sex inheritance in the guppy (Poecilia reticulata). Genetics 2009, 182, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Nanda, I.; Feichtinger, W.; Schmid, M.; Schröder, J.; Zischler, H.; Epplen, J. Simple repetitive sequences are associated with differentiation of the sex-chromosomes in the guppy fish. J. Mol. Evol. 1990, 30, 456–462. [Google Scholar] [CrossRef]
- Nanda, I.; Schories, S.; Tripathi, N.; Dreyer, C.; Haaf, T.; Schmid, M.; Schartl, M. Sex chromosome polymorphism in guppies. Chromosoma 2014, 123, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Lisachov, A.; Zadesenets, K.; Rubtsov, N.; Borodin, P. Sex chromosome synapsis and recombination in male guppies. Zebrafish 2015, 12, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Russell, S.; Magurran, A. Intrinsic reproductive isolation between Trinidadian populations of the guppy, Poecilia reticulata. J. Evol. Biol. 2006, 19, 1294–1303. [Google Scholar] [CrossRef] [PubMed]
- Amores, A.; Catchen, J.; Nanda, I.; Warren, W.; Walter, R.; Schartl, M.; Postlethwait, J.H. A RAD-tag genetic map for the Platyfish (Xiphophorus maculatus) reveals mechanisms of karyotype evolution among Teleost fish. Genetics 2014, 197, 625–641. [Google Scholar] [CrossRef] [PubMed]
- Korunes, K.; Noor, M. Gene conversion and linkage: Effects on genome evolution and speciation. Mol. Ecol. 2017, 26, 351–364. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.; Smith, C.; Kazemi, Y.; Cockrell, A.; Arvanitakas, A.; Blumenstie, J.; Jaspersen, S.; Hawley, R. Whole-genome analysis of individual meiotic events in Drosophila melanogaster reveals that noncrossover gene conversions are insensitive to interference and the centromere effect. Genetics 2016, 203, 159–171. [Google Scholar] [CrossRef] [PubMed]
- Kai, W.; Kikuchi, K.; Fujita, M.; Fujiwara, A.; Yoshiura, Y.; Ototake, M.; Venkatesh, B.; Miyaki, K.; Suzuki, Y. A genetic linkage map for the Tiger Pufferfish, Takifugu rubripes. Genetics 2005, 171, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, T.; Danzmann, R.; Gharbi, K.; Howard, P.; Ozaki, A.; Khoo, S.; Woram, R.; Okamoto, N.; Ferguson, M.M.; Holm, L.; et al. A microsatellite linkage map of rainbow trout (Oncorhynchus mykiss) characterized by large sex-specific differences in recombination rates. Genetics 2000, 155, 1331–1345. [Google Scholar] [PubMed]
- Singer, A.; Perlman, H.; Yan, Y.; Walker, C.; Corley-Smith, G.; Brandhorst, B.; Postlethwait, J. Sex-specific recombination rates in zebrafish (Danio rerio). Genetics 2002, 160, 649–657. [Google Scholar] [CrossRef] [PubMed]
- Reid, D.; Smith, C.; Rommens, M.; Blanchard, B.; Martin-Robichaud, D.; Reith, M. A genetic linkage map of Atlantic halibut (Hippoglossus hippoglossus L.). Genetics 2007, 177, 1193–1205. [Google Scholar] [CrossRef] [PubMed]
- Sardell, J.; Cheng, C.; Dagilis, A.; Ishikawa, A.; Kitano, J.; Peichel, C.; Kirkpatrick, M. Sex differences in recombination in sticklebacks. G3-Genes Genomes Genet. 2018. [Google Scholar] [CrossRef] [PubMed]
- May, B.; Delany, M. Meiotic models to explain classical linkage, pseudolinkage, and chromosomal pairing in tetraploid derivative salmonid genomes: II. Wright is still right. J. Hered. 2015, 106, 762–766. [Google Scholar] [CrossRef] [PubMed]
- Kai, W.; Kikuchi, K.; Tohari, S.; Chew, A.; Tay, A.; Fujiwara, A.; Hosoya, S.; Suetake, H.; Naruse, K.; Brenner, S.; et al. Integration of the genetic map and genome assembly of fugu facilitates insights into distinct features of genome evolution in teleosts and mammals. Genome Biol. Evol. 2011, 3, 424–442. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, N.; Studer, T.; Dufresne, C.; Perrin, N. Sex-chromosome recombination in common frogs brings water to the fountain-of-youth. Mol. Biol. Evol. 2018, 35, 942–948. [Google Scholar] [CrossRef] [PubMed]
- Traut, W.; Winking, H. Meiotic chromosomes and stages of sex chromosome evolution in fish: Zebrafish, platyfish and guppy. Chromosome Res. 2001, 9, 659–672. [Google Scholar] [CrossRef] [PubMed]
- Gammerdinger, W.; Conte, M.; Sandkam, B.; Ziegelbecker, A.; Koblmüller, S.; Kocher, T. Novel sex chromosomes in 3 Cichlid fishes from lake Tanganyika. J. Hered. 2018, 15. [Google Scholar] [CrossRef] [PubMed]
- Taboada, X.; Hermida, M.; Vera, M.; Piferrer, F.; Viñas, A.; Bouza, C.; Martínez, P. Identification of the major sex-determining region of turbot (Scophthalmus maximus). G3-Genes Genomes Genet. 2014, 4, 1871–1880. [Google Scholar] [CrossRef]
- Wright, A.; Darolti, I.; Bloch, N.; Oostra, V.; Sandkam, B.; Buechel, S.; Kolm, N.; Breden, F.; Vicoso, B.; Mank, J. Convergent recombination suppression suggests a role of sexual conflict in guppy sex chromosome formation. Nat. Commun. 2017, 8, 14251. [Google Scholar] [CrossRef] [PubMed]
- Willing, E.-M.; Bentzen, P.; van Oosterhout, C.; Hoffmann, M.; Cable, J.; Breden, F.; Weigel, D.; Dreyer, C. Genome-wide single nucleotide polymorphisms reveal population history and adaptive divergence in wild guppies. Mol. Ecol. 2010, 19, 968–984. [Google Scholar] [CrossRef] [PubMed]
- Fraser, B.A.; Künstner, A.; Reznick, D.N.; Dreyer, C.; Weigel, D. Population genomics of natural and experimental populations of guppies (Poecilia reticulata). Mol. Ecol. 2015, 24, 389–408. [Google Scholar] [CrossRef] [PubMed]
- Kirkpatrick, M.; Guerrero, R. Signatures of sex-antagonistic selection on recombining sex chromosomes. Genetics 2014, 197, 531–541. [Google Scholar] [CrossRef] [PubMed]
- Qiu, S.; Bergero, R.; Charlesworth, D. Testing for the footprint of sexually antagonistic polymorphisms in the pseudo-autosomal region of a plant sex chromosome pair. Genetics 2013, 194, 663–672. [Google Scholar] [CrossRef] [PubMed]
- Bohne, A.; Wilson, C.; Postlethwait, J.; Salzburger, W. Variations on a theme: Genomics of sex determination in the cichlid fish Astatotilapia burtoni. BMC Genom. 2016, 17, 883. [Google Scholar] [CrossRef] [PubMed]
- Meisel, R.P.; Gonzales, C.A.; Luu, H. The house fly Y Chromosome is young and minimally differentiated from its ancient X chromosome partner. Genome Res. 2017, 27, 1417–1426. [Google Scholar] [CrossRef] [PubMed]
- Meisel, R.P.; Scott, J.G.; Clark, A. Transcriptome differences between alternative sex determining genotypes in the house fly, Musca domestica. Genome Biol. Evol. 2015, 7, 2051–2061. [Google Scholar] [CrossRef] [PubMed]
- Hamm, R.L.; Meisel, R.P.; Scott, J.G. The evolving puzzle of autosomal versus Y-linked male determination in Musca domestica. G3 Genes Genomes Genet. 2015, 5, 371–384. [Google Scholar] [CrossRef] [PubMed]
- Brelsford, A.; Rodrigues, N.; Perrin, N. High-density linkage maps fail to detect any genetic component to sex determination in a Rana temporaria family. J. Evol. Biol. 2016, 29, 220–225. [Google Scholar] [CrossRef] [PubMed]
- Dufresnes, C.; Bertholet, Y.; Wassef, J.; Ghali, K.; Savary, R.; Pasteur, B.; Brelsford, A.; Rozenblut-Kościsty, B.; Ogielska, M.; Stöck, M.; et al. Sex-chromosome differentiation parallels postglacial range expansion in European tree frogs (Hyla arborea). Evolution 2014, 68, 3445–3456. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, R.; Kirkpatrick, M.; Perrin, N. Cryptic recombination in the ever-young sex chromosomes of Hylid frogs. J. Evol. Biol. 2012, 25, 1947–1954. [Google Scholar] [CrossRef] [PubMed]
- Brelsford, A.; Dufresnes, C.; Perrin, N. High-density sex-specific linkage maps of a European tree frog (Hyla arborea) identify the sex chromosome without information on offspring sex. Heredity 2015, 116, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, N.; Studer, T.; Dufresne, C.; Ma, R.; Veltsos, P.; Perrin, N. Dmrt1 polymorphism and sex chromosome differentiation in Rana temporaria. Mol. Ecol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Lenormand, T. The evolution of sex dimorphism in recombination. Genetics 2003, 163, 811–822. [Google Scholar] [PubMed]
- Hinch, A.G.; Altemose, N.; Noor, N.; Donnelly, P.; Myers, S.R. Recombination in the human pseudoautosomal region PAR1. PLoS Genet. 2014, 10, e1004503. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Males | Females | Male parent genotype | ||
|---|---|---|---|---|
| Sb | Non-Sb | Sb | Non-Sb | |
| 11 | 0 | 0 | 8 | X(−)/Y(Sb) |
| 7 | 0 | 0 | 6 | X(−)/Y(Sb) |
| 18 | 0 | 0 | 13 | X(−)/Y(Sb) |
| 14 | 0 | 0 | 16 | X(−)/Y(Sb) |
| 16 | 0 | 0 | 18 | X(−)/Y(Sb) |
| 11 | 0 | 0 | 7 | X(−)/Y(Sb) |
| 5 | 0 | 0 | 7 | X(−)/Y(Sb) |
| 9 | 0 | 0 | 5 | X(−)/Y(Sb) |
| 20 | 0 | 0 | 17 | X(−)/Y(Sb) |
| 40 | 0 | 0 | 35 | X(−)/Y(Sb) |
| 32 | 0 | 0 | 29 | X(−)/Y(Sb) |
| 28 | 0 | 0 | 18 | X(−)/Y(Sb) |
| 23 | 0 | 0 | 19 | X(−)/Y(Sb) |
| 7 | 0 | 0 | 14 | X(−)/Y(Sb) |
| 19 | 0 | 0 | 3 | X(−)/Y(Sb) |
| 31 | 0 | 0 | 25 | X(−)/Y(Sb) |
| 15 | 0 | 1 | 14 | X(−)/Y(Sb) |
| 11 | 0 | 0 | 13 | X(−)/Y(Sb) |
| 20 | 0 | 0 | 16 | X(−)/Y(Sb) |
| 30 | 0 | 0 | 19 | X(−)/Y(Sb) |
| 27 | 0 | 0 | 23 | X(−)/Y(Sb) |
| 20 | 0 | 0 | 10 | X(−)/Y(Sb) |
| 23 | 0 | 0 | 20 | X(−)/Y(Sb) |
| 11 | 0 | 0 | 9 | X(−)/Y(Sb) |
| 18 | 0 | 0 | 7 | X(−)/Y(Sb) |
| 12 | 0 | 0 | 12 | X(−)/Y(Sb) |
| 20 | 0 | 0 | 13 | X(−)/Y(Sb) |
| 21 | 0 | 0 | 10 | X(−)/Y(Sb) |
| 7 | 0 | 0 | 15 | X(−)/Y(Sb) |
| 8 | 0 | 0 | 5 | X(−)/Y(Sb) |
| 6 | 0 | 0 | 14 | X(−)/Y(Sb) |
| 14 | 0 | 0 | 6 | X(−)/Y(Sb) |
| 11 | 0 | 0 | 12 | X(−)/Y(Sb) |
| 565 | 0 | 1 | 458 | 1024 |
| Males | Females | Male parent genotype | ||
|---|---|---|---|---|
| Sb | Non-Sb | Sb | Non-Sb | |
| 6 | 0 | 1 | 10 | X(−)/Y(Sb) |
| 8 | 0 | 0 | 15 | X(−)/Y(Sb) |
| 12 | 0 | 0 | 20 | X(−)/Y(Sb) |
| 14 | 0 | 0 | 2 | X(−)/Y(Sb) |
| 13 | 0 | 0 | 14 | X(−)/Y(Sb) |
| 11 | 0 | 0 | 3 | X(−)/Y(Sb) |
| 12 | 0 | 0 | 6 | X(−)/Y(Sb) |
| 7 | 0 | 0 | 8 | X(−)/Y(Sb) |
| 0 | 15 | 4 | 0 | X(Sb)/Y(−) |
| 3 | 11 | 10 | 0 | X(Sb)/Y(−) |
| 1 | 10 | 14 | 0 | X(Sb)/Y(−) |
| 0 | 10 | 7 | 0 | X(Sb)/Y(−) |
| 17 | 0 | 21 | 0 | X(Sb)/Y(Sb) |
| 7 | 0 | 8 | 0 | X(Sb)/Y(Sb) |
| 9 | 0 | 17 | 0 | X(Sb)/Y(Sb) |
| 5 | 0 | 3 | 0 | X(Sb)/Y(Sb) |
| 6 | 0 | 5 | 0 | X(Sb)/Y(Sb) |
| 10 | 0 | 5 | 0 | X(Sb)/Y(Sb) |
| 14 | 0 | 13 | 0 | X(Sb)/Y(Sb) |
| 155 | 46 | 108 | 78 | 387 |
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Charlesworth, D. The Guppy Sex Chromosome System and the Sexually Antagonistic Polymorphism Hypothesis for Y Chromosome Recombination Suppression. Genes 2018, 9, 264. https://doi.org/10.3390/genes9050264
Charlesworth D. The Guppy Sex Chromosome System and the Sexually Antagonistic Polymorphism Hypothesis for Y Chromosome Recombination Suppression. Genes. 2018; 9(5):264. https://doi.org/10.3390/genes9050264
Chicago/Turabian StyleCharlesworth, Deborah. 2018. "The Guppy Sex Chromosome System and the Sexually Antagonistic Polymorphism Hypothesis for Y Chromosome Recombination Suppression" Genes 9, no. 5: 264. https://doi.org/10.3390/genes9050264
APA StyleCharlesworth, D. (2018). The Guppy Sex Chromosome System and the Sexually Antagonistic Polymorphism Hypothesis for Y Chromosome Recombination Suppression. Genes, 9(5), 264. https://doi.org/10.3390/genes9050264