Regulation of the X Chromosome in the Germline and Soma of Drosophila melanogaster Males


Abstract
:1. Introduction
2. Materials and Methods
2.1. High-Throughput Expression Data
2.2. Reporter Gene Mobilization
2.3. Mapping Reporter Gene Insertions
2.4. Measuring Reporter Gene Expression
2.5. Maintenance of Drosophila Stocks
3. Results
3.1. Expression of Native Genes on the X Chromosome and Autosomes
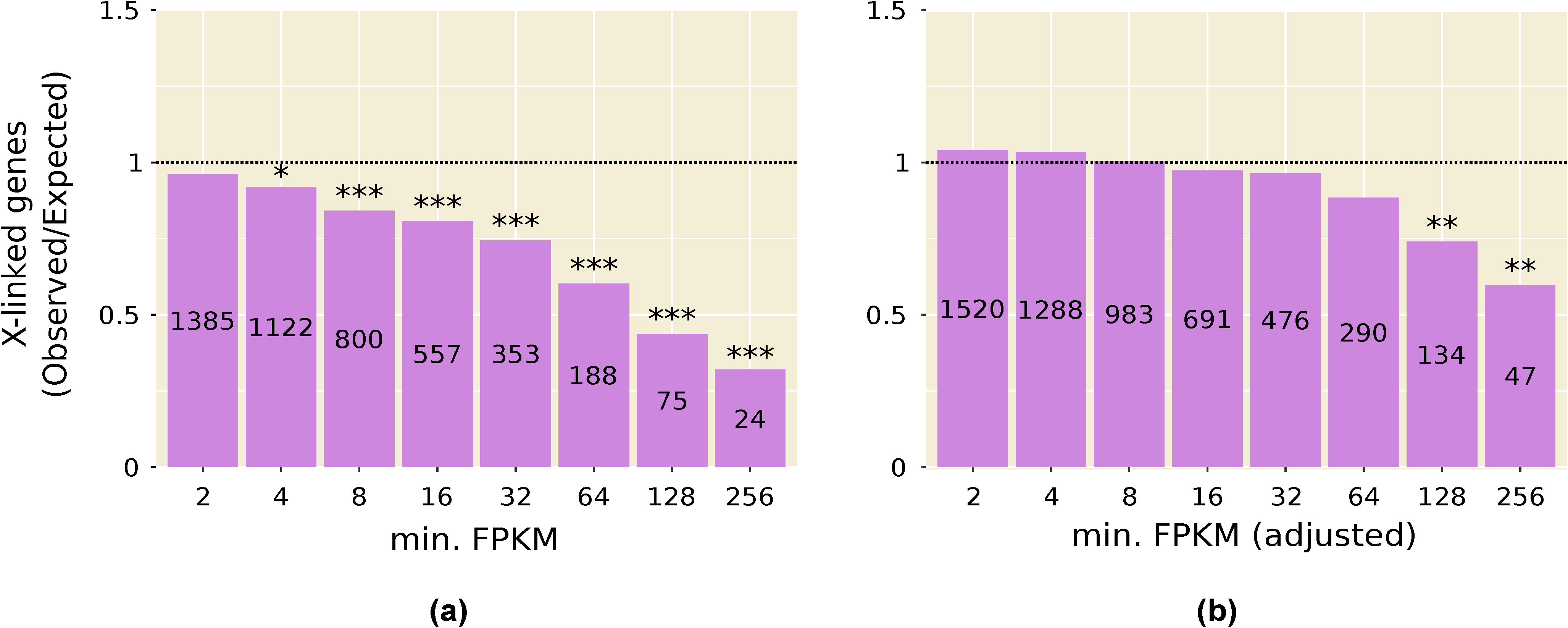
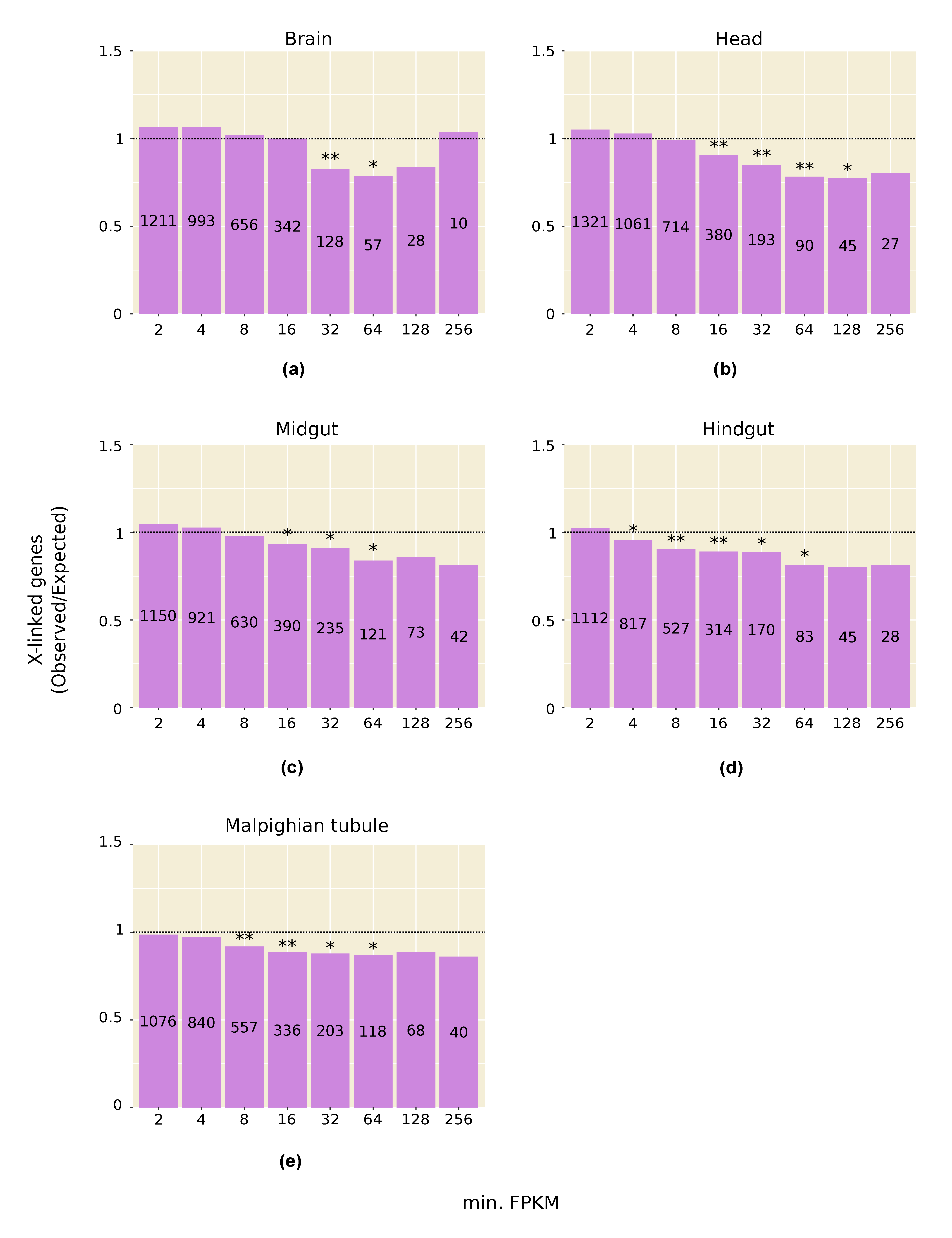
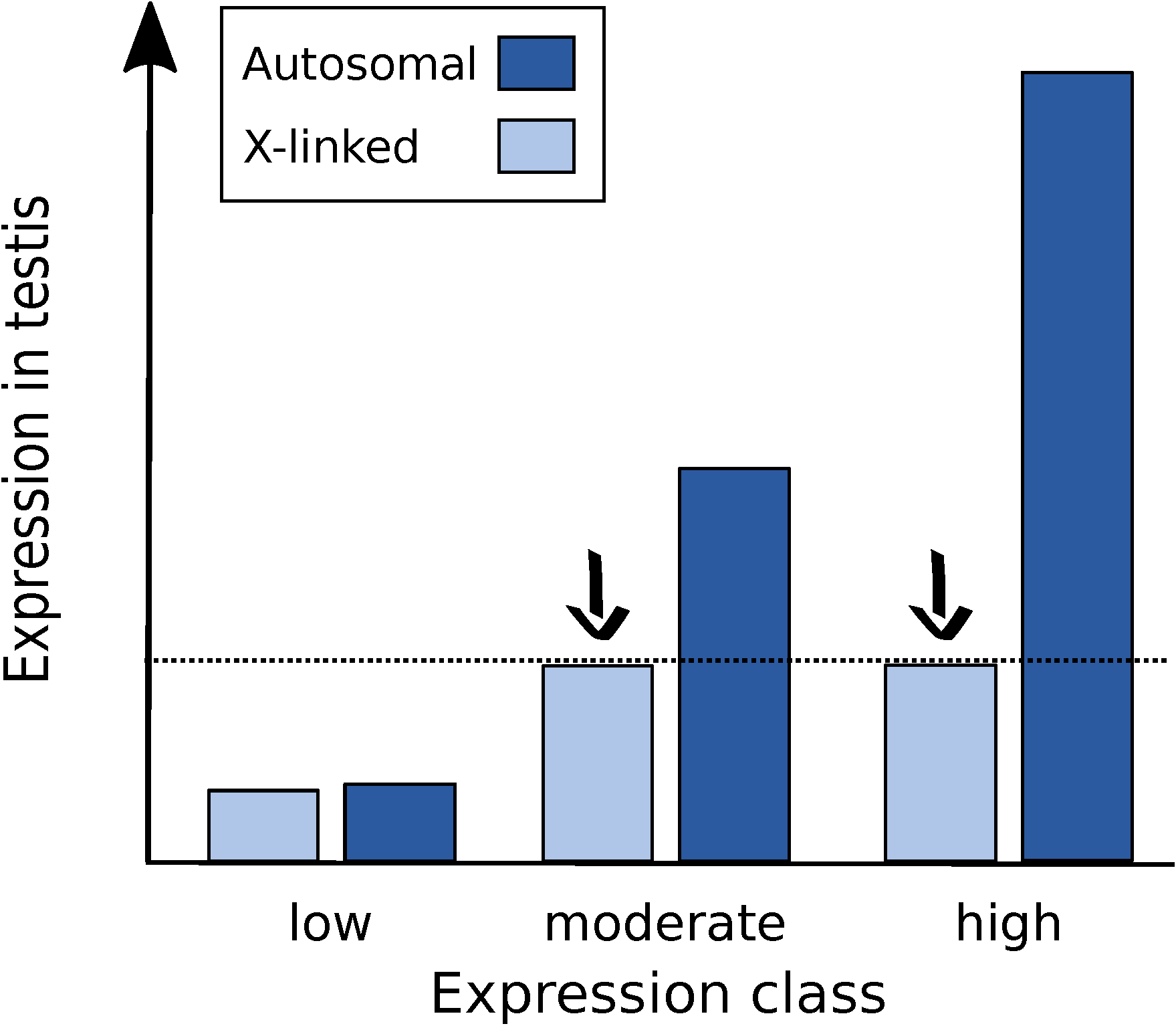
3.2. Highly Expressed Genes Are Underrepresented on the X Chromosome
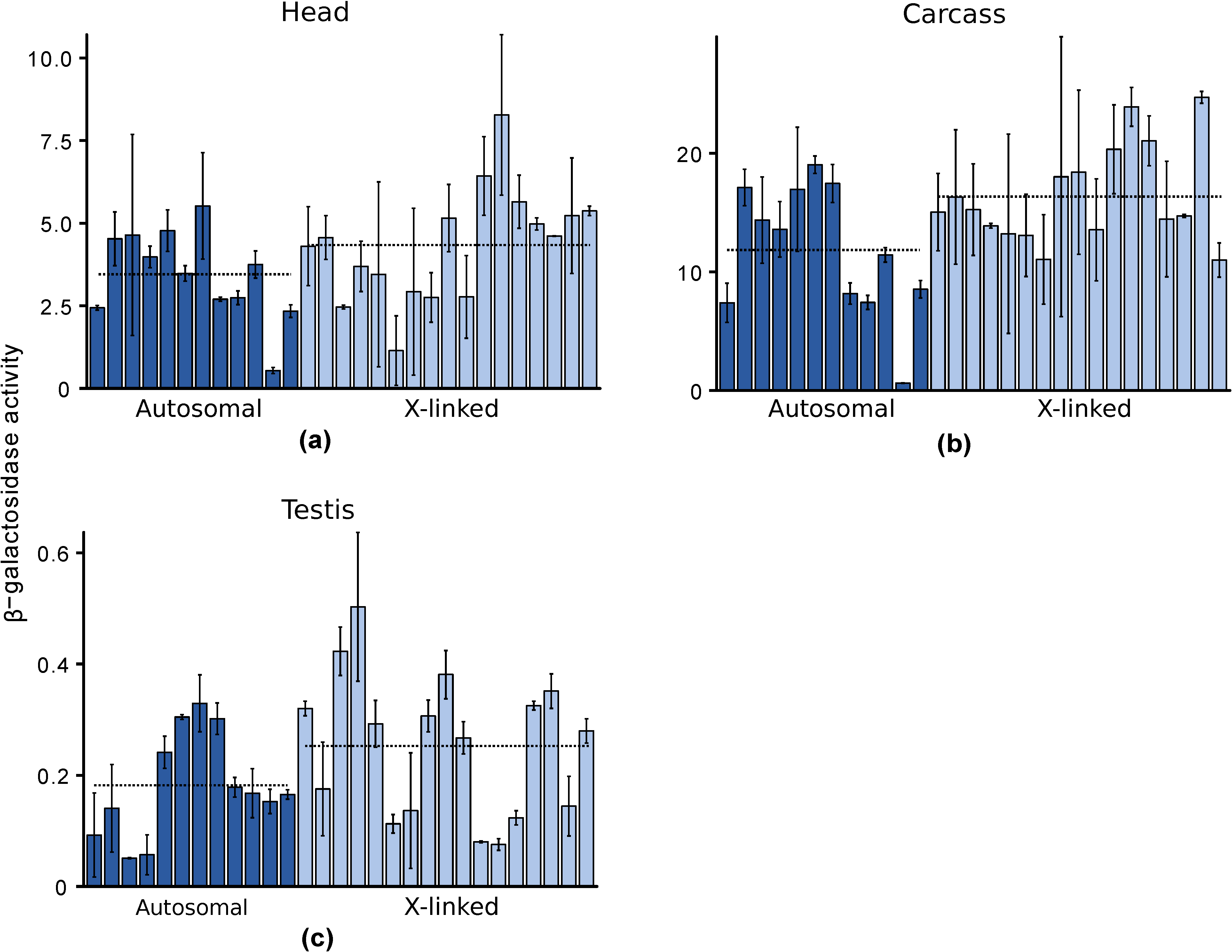
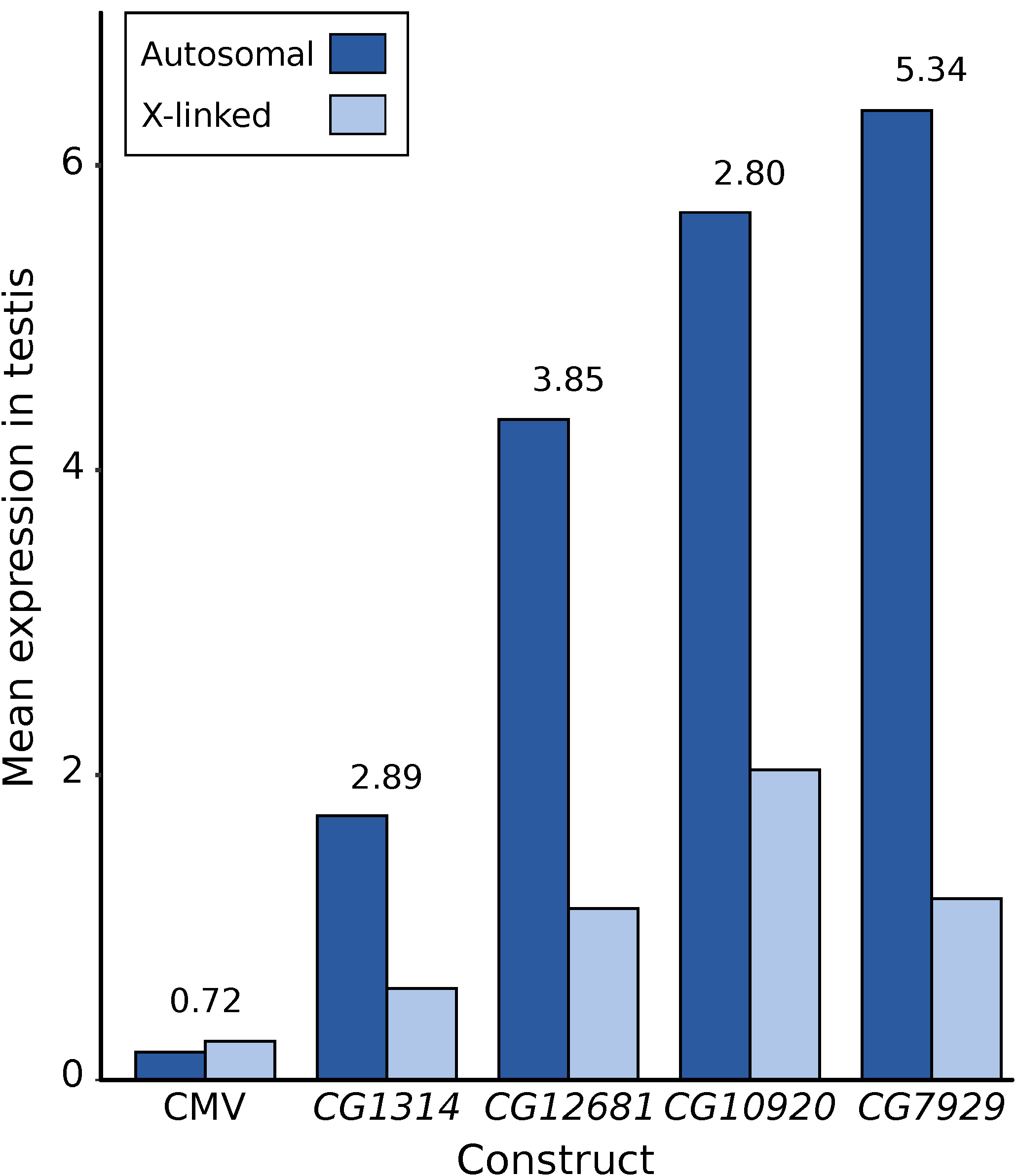
3.3. Expression of X-Linked and Autosomal Reporter Genes
4. Discussion
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Charlesworth, D.; Charlesworth, B.; Marais, G. Steps in the evolution of heteromorphic sex chromosomes. Heredity 2005, 95, 118–128. [Google Scholar] [CrossRef] [PubMed]
- Ellegren, H. Sex-chromosome evolution: Recent progress and the influence of male and female heterogamety. Nat. Rev. Genet. 2011, 12, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Bergero, R.; Charlesworth, D. The evolution of restricted recombination in sex chromosomes. Trends Ecol. Evol. 2009, 24, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Charlesworth, D. Evolution of recombination rates between sex chromosomes. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2017, 372, 20160456. [Google Scholar] [CrossRef] [PubMed]
- Charlesworth, B.; Charlesworth, D. The degeneration of Y chromosomes. Philos. Trans. R. Soc. B Biol. Sci. 2000, 355, 1563–1572. [Google Scholar] [CrossRef] [PubMed]
- Bachtrog, D. Sex chromosome evolution: Molecular aspects of Y-chromosome degeneration in Drosophila. Genome Res. 2005, 1393–1401. [Google Scholar] [CrossRef] [PubMed]
- Gramates, L.S.; Marygold, S.J.; Dos Santos, G.; Urbano, J.M.; Antonazzo, G.; Matthews, B.B.; Rey, A.J.; Tabone, C.J.; Crosby, M.A.; Emmert, D.B.; et al. FlyBase at 25: Looking to the future. Nucleic Acids Res. 2017, 45, D663–D671. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, A.B.; Lazzaro, B.P.; Clark, A.G. Y chromosomal fertility factors kl-2 and kl-3 of Drosophila melanogaster encode dynein heavy chain polypeptides. Proc. Natl. Acad. Sci. USA 2000, 97, 13239–13244. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, A.B.; Dobo, B.A.; Vibranovski, M.D.; Clark, A.G. Identification of five new genes on the Y chromosome of Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2001, 98, 13225–13230. [Google Scholar] [CrossRef] [PubMed]
- Vibranovski, M.D.; Zhang, Y.; Long, M. Out of the X chromosomal gene movement in the Drosophila genus. Genome Res. 2009, 19, 897–903. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, A.B.; Vicoso, B.; Russo, C.A.M.; Swenor, B.; Clark, A.G. Birth of a new gene on the Y chromosome of Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2015, 112, 12450–12455. [Google Scholar] [CrossRef] [PubMed]
- Kelley, R.L.; Kuroda, M.I. Equality for X chromosomes. Science 1995, 270, 1607–1610. [Google Scholar] [CrossRef] [PubMed]
- Straub, T.; Grimaud, C.; Gilfillan, G.D.; Mitterweger, A.; Becker, P.B. The chromosomal high-affinity binding sites for the Drosophila dosage compensation complex. PLoS Genet. 2008, 4, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuroda, M.I.; Hilfiker, A.; Lucchesi, J.C. Dosage compensation in Drosophila—A model for the coordinate regulation of transcription. Genetics 2016, 204, 435–450. [Google Scholar] [CrossRef] [PubMed]
- Meiklejohn, C.D.; Landeen, E.L.; Cook, J.M.; Kingan, S.B.; Presgraves, D.C. Sex chromosome-specific regulation in the Drosophila male germline but little evidence for chromosomal dosage compensation or meiotic inactivation. PLoS Biol. 2011, 9. [Google Scholar] [CrossRef] [PubMed]
- Meiklejohn, C.D.; Presgraves, D.C. Little evidence for demasculinization of the Drosophila X chromosome among genes expressed in the male germline. Genome Biol. Evol. 2012, 4, 1007–1016. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.; Parisi, M.; Sturgill, D.; Nuttall, R.; Doctolero, M.; Dudko, O.K.; Malley, J.D.; Eastman, P.S.; Oliver, B. Global analysis of X-chromosome dosage compensation. J. Biol. 2006, 5. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Hiatt, J.B.; Nguyen, D.K.; Ercan, S.; Sturgill, D.; Hillier, L.W.; Schlesinger, F.; Davis, C.A.; Reinke, V.J.; Gingeras, T.R.; et al. Evidence for compensatory upregulation of expressed X-linked genes in mammals, Caenorhabditis elegans and Drosophila melanogaster. Nat. Genet. 2011, 43, 1179–1185. [Google Scholar] [CrossRef] [PubMed]
- Hense, W.; Baines, J.F.; Parsch, J. X chromosome inactivation during Drosophila spermatogenesis. PLoS Biol. 2007, 5, 2288–2295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kemkemer, C.; Hense, W.; Parsch, J. Fine-scale analysis of X chromosome inactivation in the male germ line of Drosophila melanogaster. Mol. Biol. Evol. 2011, 28, 1561–1563. [Google Scholar] [CrossRef] [PubMed]
- Kemkemer, C.; Catalán, A.; Parsch, J. “Escaping” the X chromosome leads to increased gene expression in the male germline of Drosophila melanogaster. Heredity 2014, 112, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Landeen, E.L.; Muirhead, C.A.; Wright, L.; Meiklejohn, C.D.; Presgraves, D.C. Sex chromosome-wide transcriptional suppression and compensatory cis-regulatory evolution mediate gene expression in the Drosophila male germline. PLoS Biol. 2016, 14, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Vibranovski, M.D.; Lopes, H.F.; Karr, T.L.; Long, M. Stage-specific expression profiling of Drosophila spermatogenesis suggests that meiotic sex chromosome inactivation drives genomic relocation of testis-expressed genes. PLoS Genet. 2009, 5. [Google Scholar] [CrossRef] [PubMed]
- Vibranovski, M.D. Meiotic sex chromosome inactivation in Drosophila. J. Genom. 2014, 2, 104–117. [Google Scholar] [CrossRef] [PubMed]
- Leader, D.P.; Krause, S.A.; Pandit, A.; Davies, S.A.; Dow, J.A.T. FlyAtlas 2: A new version of the Drosophila melanogaster expression atlas with RNA-Seq, miRNA-Seq and sex-specific data. Nucleic Acids Res. 2018, 46, D809–D815. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; ISBN 3-900051-07-0. [Google Scholar]
- Parsch, J. Functional analysis of Drosophila melanogaster gene regulatory sequences by transgene coplacement. Genetics 2004, 168, 559–561. [Google Scholar] [CrossRef] [PubMed]
- Robertson, H.M.; Preston, C.R.; Phillis, R.W.; Johnson-Schlitz, D.M.; Benz, W.K.; Engels, W.R. A stable genomic source of P element transposase in Drosophila melanogaster. Genetics 1988, 118, 461–470. [Google Scholar] [PubMed]
- Bellen, H.J.; Levis, R.W.; Liao, G.; He, Y.; Carlson, J.W.; Tsang, G.; Evans-Holm, M.; Hiesinger, P.R.; Schulze, K.L.; Rubin, G.M.; et al. The BDGP gene disruption project: Single transposon insertions associated with 40% of Drosophila genes. Genetics 2004, 167, 761–781. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Stenberg, P.; Lundberg, L.E.; Johansson, A.M.; Rydén, P.; Svensson, M.J.; Larsson, J. Buffering of segmental and chromosomal aneuploidies in Drosophila melanogaster. PLoS Genet. 2009, 5. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Malone, J.H.; Powell, S.K.; Periwal, V.; Spana, E.; MacAlpine, D.M.; Oliver, B. Expression in aneuploid Drosophila S2 cells. PLoS Biol. 2010, 8. [Google Scholar] [CrossRef] [PubMed]
- Vicoso, B.; Charlesworth, B. The deficit of male-biased genes on the D. melanogaster X chromosome is expression-dependent: A consequence of dosage compensation? J. Mol. Evol. 2009, 68, 576–583. [Google Scholar] [CrossRef] [PubMed]
- Hurst, L.D.; Ghanbarian, A.T.; Forrest, A.R.R.; Huminiecki, L.; Rehli, M.; Kenneth Baillie, J.; de Hoon, M.J.L.; Haberle, V.; Lassmann, T.; Kulakovskiy, I.V.; et al. The constrained maximal expression level owing to haploidy shapes gene content on the mammalian X chromosome. PLoS Biol. 2015, 13, 1–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Argyridou, E.; Huylmans, A.K.; Königer, A.; Parsch, J. X-linkage is not a general inhibitor of tissue-specific gene expression in Drosophila melanogaster. Heredity 2017, 119, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Schoenmakers, S.; Wassenaar, E.; Hoogerbrugge, J.W.; Laven, J.S.E.; Grootegoed, J.A.; Baarends, W.M. Female meiotic sex chromosome inactivation in chicken. PLoS Genet. 2009, 5. [Google Scholar] [CrossRef] [PubMed]
- Cabrero, J.; Teruel, M.; Carmona, F.D.; Jiménez, R.; Camacho, J.P.M. Histone H3 lysine 9 acetylation pattern suggests that X and B chromosomes are silenced during entire male meiosis in a grasshopper. Cytogenet. Genome Res. 2007, 119, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Bean, C.J.; Schaner, C.E.; Kelly, W.G. Meiotic pairing and imprinted X chromatin assembly in Caenorhabditis elegans. Nat. Genet. 2004, 36, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Da Cruz, I.; Rodríguez-Casuriaga, R.; Santiñaque, F.F.; Farías, J.; Curti, G.; Capoano, C.A.; Folle, G.A.; Benavente, R.; Sotelo-Silveira, J.R.; Geisinger, A. Transcriptome analysis of highly purified mouse spermatogenic cell populations: Gene expression signatures switch from meiotic- to postmeiotic-related processes at pachytene stage. BMC Genom. 2016, 17, 294. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.M.A. Meiotic sex chromosome inactivation. Development 2007, 134, 1823–1831. [Google Scholar] [CrossRef] [PubMed]
- Bell, G. The Masterpiece of Nature: The Evolution and Genetics of Sexuality; University of California Press: Berkeley, CA, USA, 1982; ISBN 9780856647536. [Google Scholar]
- Hassold, T.; Hunt, P. To err (meiotically) is human: The genesis of human aneuploidy. Nat. Rev. Genet. 2001, 2, 280–291. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.I.; Xu, E.Y. Sexual antagonism and X inactivation—The SAXI hypothesis. Trends Genet. 2003, 19, 243–247. [Google Scholar] [CrossRef]
- Tao, Y.; Hartl, D.L.; Laurie, C.C. Sex-ratio segregation distortion associated with reproductive isolation in Drosophila. Proc. Natl. Acad. Sci. USA 2001, 98, 13183–13188. [Google Scholar] [CrossRef] [PubMed]
- Montchamp-Moreau, C. Sex-ratio meiotic drive in Drosophila simulans: Cellular mechanism, candidate genes and evolution. Biochem. Soc. Trans. 2006, 34, 562–565. [Google Scholar] [CrossRef] [PubMed]
- Phadnis, N.; Allen Orr, H. A single gene causes both male sterility and segre. Science 2009, 323, 376–379. [Google Scholar] [CrossRef] [PubMed]
- Paczolt, K.A.; Reinhardt, J.A.; Wilkinson, G.S. Contrasting patterns of X-chromosome divergence underlie multiple sex-ratio polymorphisms in stalk-eyed flies. J. Evol. Biol. 2017, 30, 1772–1784. [Google Scholar] [CrossRef] [PubMed]
- Hurst, L.D.; Pomiankowski, A. The eyes have it. Nature 1998, 391, 223–224. [Google Scholar] [CrossRef] [PubMed]
- Huylmans, A.K.; Parsch, J. Variation in the X: Autosome distribution of male-biased genes among Drosophila melanogaster tissues and its relationship with dosage compensation. Genome Biol. Evol. 2015, 7, 1960–1971. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tissue | X Genes | X Median | A Genes | A Median | X/A | p |
|---|---|---|---|---|---|---|
| Testis | 1581 | 8.16 | 8657 | 12.46 | 0.65 | <2.2−16 |
| Brain | 1355 | 7.73 | 6871 | 8.16 | 0.95 | 0.22 |
| Head | 1471 | 7.64 | 7595 | 8.35 | 0.91 | 0.007 |
| Midgut | 1268 | 7.88 | 6649 | 8.94 | 0.88 | 0.10 |
| Hindgut | 1249 | 6.27 | 6661 | 7.58 | 0.83 | 0.0006 |
| Tubule | 1258 | 6.76 | 6962 | 7.43 | 0.91 | 0.08 |
| Tissue | X Genes | X Median | A Genes | A Median | X/A | p |
|---|---|---|---|---|---|---|
| Ovaries | 1206 | 10.78 | 5723 | 10.52 | 1.02 | 0.56 |
| Brain | 1356 | 7.82 | 6904 | 7.98 | 0.98 | 0.90 |
| Head | 1474 | 7.99 | 7580 | 8.50 | 0.94 | 0.05 |
| Midgut | 1194 | 6.82 | 6065 | 8.31 | 0.82 | 0.004 |
| Hindgut | 1289 | 7.74 | 6687 | 8.91 | 0.87 | 0.01 |
| Tubule | 1226 | 7.82 | 6332 | 8.84 | 0.88 | 0.12 |
| Tissue | X Genes | X Median | A Genes | A Median | X/A | p |
|---|---|---|---|---|---|---|
| Testis | 1460 | 7.17 | 7245 | 8.99 | 0.80 | 2.4−05 |
| Brain | 1317 | 7.47 | 6594 | 7.72 | 0.97 | 0.59 |
| Head | 1408 | 7.21 | 7127 | 7.60 | 0.95 | 0.09 |
| Midgut | 1171 | 7.10 | 6026 | 7.44 | 0.95 | 0.47 |
| Hindgut | 1188 | 5.89 | 6233 | 6.87 | 0.86 | 0.05 |
| Tubule | 1164 | 5.97 | 6391 | 6.44 | 0.93 | 0.13 |
| Tissue | X Genes | X Median | A Genes | A Median | X/A Median | p |
|---|---|---|---|---|---|---|
| Ovaries | 1158 | 10.34 | 5485 | 10.09 | 1.02 | 0.47 |
| Brain | 1313 | 7.54 | 6638 | 7.57 | 1.00 | 0.78 |
| Head | 1398 | 7.44 | 7113 | 7.74 | 0.96 | 0.15 |
| Midgut | 1108 | 6.35 | 5521 | 7.14 | 0.89 | 0.03 |
| Hindgut | 1213 | 7.20 | 6196 | 7.90 | 0.91 | 0.06 |
| Tubule | 1121 | 6.88 | 5750 | 7.44 | 0.92 | 0.17 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Argyridou, E.; Parsch, J. Regulation of the X Chromosome in the Germline and Soma of Drosophila melanogaster Males. Genes 2018, 9, 242. https://doi.org/10.3390/genes9050242
Argyridou E, Parsch J. Regulation of the X Chromosome in the Germline and Soma of Drosophila melanogaster Males. Genes. 2018; 9(5):242. https://doi.org/10.3390/genes9050242
Chicago/Turabian StyleArgyridou, Eliza, and John Parsch. 2018. "Regulation of the X Chromosome in the Germline and Soma of Drosophila melanogaster Males" Genes 9, no. 5: 242. https://doi.org/10.3390/genes9050242
APA StyleArgyridou, E., & Parsch, J. (2018). Regulation of the X Chromosome in the Germline and Soma of Drosophila melanogaster Males. Genes, 9(5), 242. https://doi.org/10.3390/genes9050242