Towards Long-Range RNA Structure Prediction in Eukaryotic Genes


Abstract
:1. Introduction
2. Instances of Long-Range RNA Structure
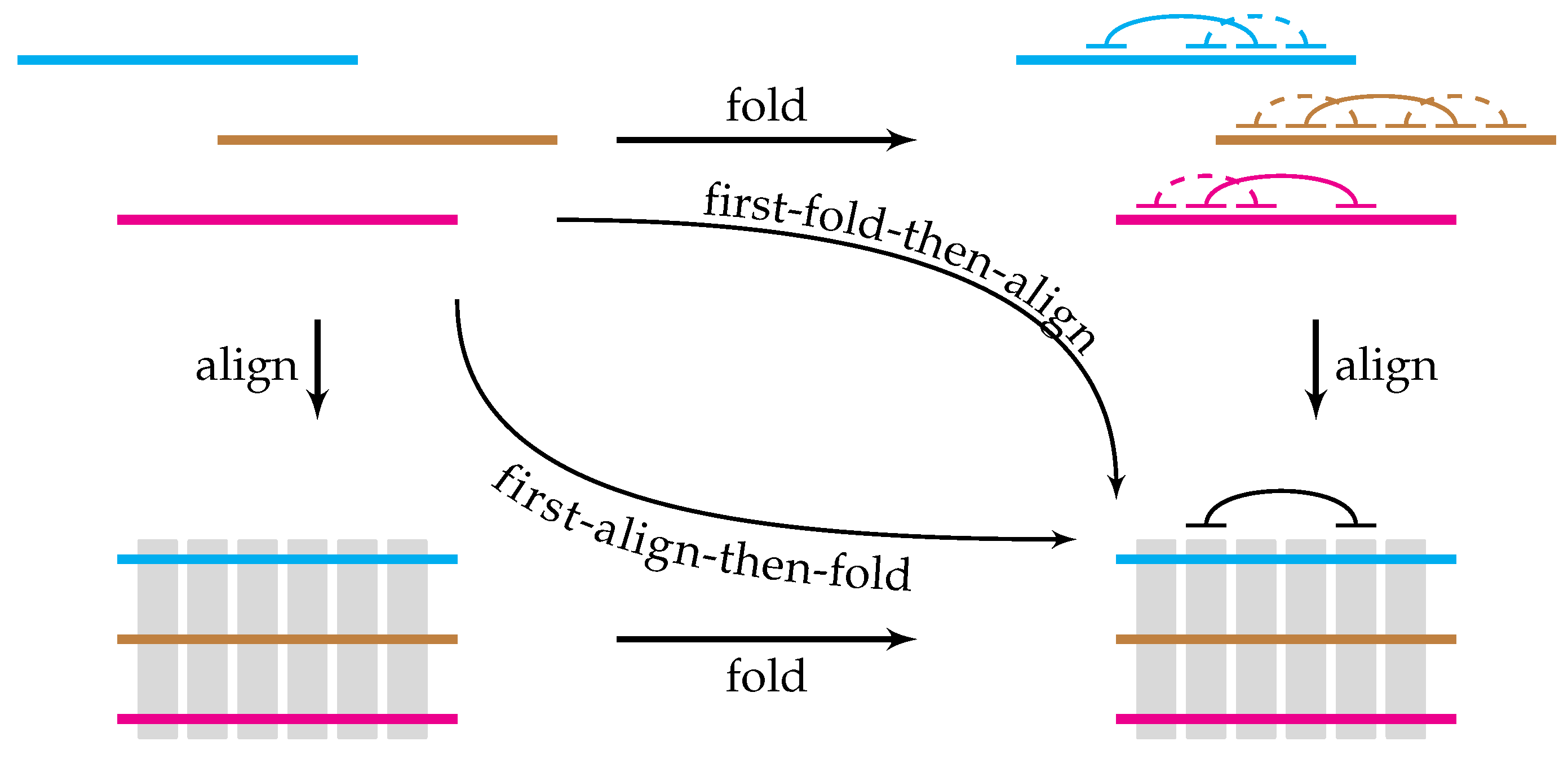
3. Predicting Long-Range RNA Structure
- most long-range RNA structures are well-conserved;
- the core of a long-range RNA structure is a long, nearly-perfect complementary match;
- elements of long-range RNA structures are located in syntenic regions.
4. Concluding Remarks
Funding
Acknowledgments
Conflicts of Interest
References
- Saldi, T.; Cortazar, M.A.; Sheridan, R.M.; Bentley, D.L. Coupling of RNA Polymerase II transcription elongation with pre-mRNA Splicing. J. Mol. Biol. 2016, 428, 2623–2635. [Google Scholar] [CrossRef] [PubMed]
- Kaida, D. The reciprocal regulation between splicing and 3’-end processing. Wiley Interdiscip. Rev. RNA 2016, 7, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Lepennetier, G.; Catania, F. Exploring the impact of cleavage and polyadenylation factors on pre-mRNA splicing across Eukaryotes. G3 Genes Genomes Genet. 2017, 7, 2107–2114. [Google Scholar] [CrossRef] [PubMed]
- Laurencikiene, J.; Kallman, A.M.; Fong, N.; Bentley, D.L.; Ohman, M. RNA editing and alternative splicing: The importance of co-transcriptional coordination. EMBO Rep. 2006, 7, 303–307. [Google Scholar] [CrossRef] [PubMed]
- Solomon, O.; Oren, S.; Safran, M.; Deshet-Unger, N.; Akiva, P.; Jacob-Hirsch, J.; Cesarkas, K.; Kabesa, R.; Amariglio, N.; Unger, R.; et al. Global regulation of alternative splicing by adenosine deaminase acting on RNA (ADAR). RNA 2013, 19, 591–604. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, R.; Barta, A.; Semrad, K. Strategies for RNA folding and assembly. Nat. Rev. Mol. Cell Biol. 2004, 5, 908–919. [Google Scholar] [CrossRef] [PubMed]
- Brion, P.; Westhof, E. Hierarchy and dynamics of RNA folding. Annu. Rev. Biophys. Biomol. Struct. 1997, 26, 113–137. [Google Scholar] [CrossRef] [PubMed]
- Herschlag, D. RNA chaperones and the RNA folding problem. J. Biol. Chem. 1995, 270, 20871–20874. [Google Scholar] [CrossRef] [PubMed]
- Van Nostrand, E.L.; Pratt, G.A.; Shishkin, A.A.; Gelboin-Burkhart, C.; Fang, M.Y.; Sundararaman, B.; Blue, S.M.; Nguyen, T.B.; Surka, C.; Elkins, K.; et al. Robust transcriptome-wide discovery of RNA-binding protein binding sites with enhanced CLIP (eCLIP). Nat. Methods 2016, 13, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Paulsen, M.T.; Veloso, A.; Prasad, J.; Bedi, K.; Ljungman, E.A.; Magnuson, B.; Wilson, T.E.; Ljungman, M. Use of Bru-Seq and BruChase-Seq for genome-wide assessment of the synthesis and stability of RNA. Methods 2014, 67, 45–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, S.; Higgs, P. Evidence for kinetic effects in the folding of large RNA molecules. J. Chem. Phys. 1996, 105, 7152. [Google Scholar] [CrossRef]
- Lai, D.; Proctor, J.R.; Meyer, I.M. On the importance of cotranscriptional RNA structure formation. RNA 2013, 19, 1461–1473. [Google Scholar] [CrossRef] [PubMed]
- Edlind, T.D.; Cooley, T.E.; Richards, S.H.; Ihler, G.M. Long range base-pairing in the leftward transcription unit of bacteriophage lambda. Characterization by electron microscopy and computer-aided sequence analysis. J. Mol. Biol. 1984, 179, 351–365. [Google Scholar] [CrossRef]
- Nicholson, B.L.; White, K.A. Functional long-range RNA-RNA interactions in positive-strand RNA viruses. Nat. Rev. Microbiol. 2014, 12, 493–504. [Google Scholar] [CrossRef] [PubMed]
- Archer, E.J.; Simpson, M.A.; Watts, N.J.; O’Kane, R.; Wang, B.; Erie, D.A.; McPherson, A.; Weeks, K.M. Long-range architecture in a viral RNA genome. Biochemistry 2013, 52, 3182–3190. [Google Scholar] [CrossRef] [PubMed]
- Shin, M.K.; Kim, J.H.; Ryu, D.K.; Ryu, W.S. Circularization of an RNA template via long-range base pairing is critical for hepadnaviral reverse transcription. Virology 2008, 371, 362–373. [Google Scholar] [CrossRef] [PubMed]
- Fricke, M.; Dunnes, N.; Zayas, M.; Bartenschlager, R.; Niepmann, M.; Marz, M. Conserved RNA secondary structures and long-range interactions in hepatitis C viruses. RNA 2015, 21, 1219–1232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Borba, L.; Villordo, S.M.; Iglesias, N.G.; Filomatori, C.V.; Gebhard, L.G.; Gamarnik, A.V. Overlapping local and long-range RNA-RNA interactions modulate dengue virus genome cyclization and replication. J. Virol. 2015, 89, 3430–3437. [Google Scholar] [CrossRef] [PubMed]
- Ooms, M.; Abbink, T.E.; Pham, C.; Berkhout, B. Circularization of the HIV-1 RNA genome. Nucleic Acids Res. 2007, 35, 5253–5261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pervouchine, D.D.; Khrameeva, E.E.; Pichugina, M.Y.; Nikolaienko, O.V.; Gelfand, M.S.; Rubtsov, P.M.; Mironov, A.A. Evidence for widespread association of mammalian splicing and conserved long-range RNA structures. RNA 2012, 18, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Tajima, Y.; Iwakawa, H.O.; Kaido, M.; Mise, K.; Okuno, T. A long-distance RNA-RNA interaction plays an important role in programmed −1 ribosomal frameshifting in the translation of p88 replicase protein of Red clover necrotic mosaic virus. Virology 2011, 417, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Ruegsegger, U.; Leber, J.H.; Walter, P. Block of HAC1 mRNA translation by long-range base pairing is released by cytoplasmic splicing upon induction of the unfolded protein response. Cell 2001, 107, 103–114. [Google Scholar] [CrossRef]
- Watters, K.E.; Lucks, J.B. Mapping RNA structure in vitro with SHAPE chemistry and next-generation sequencing (SHAPE-Seq). Methods Mol. Biol. 2016, 1490, 135–162. [Google Scholar] [PubMed]
- Shen, C.K.; Hearst, J.E. A technique for relating long-range base pairing on single-stranded DNA and eukaryotic RNA processing. Anal. Biochem. 1979, 95, 108–116. [Google Scholar] [CrossRef]
- Ramani, V.; Qiu, R.; Shendure, J. High-throughput determination of RNA structure by proximity ligation. Nat. Biotechnol. 2015, 33, 980–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aw, J.G.; Shen, Y.; Wilm, A.; Sun, M.; Lim, X.N.; Boon, K.L.; Tapsin, S.; Chan, Y.S.; Tan, C.P.; Sim, A.Y.; et al. In vivo mapping of eukaryotic RNA interactomes reveals principles of higher-order organization and regulation. Mol. Cell 2016, 62, 603–617. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Zhang, Q.C.; Lee, B.; Flynn, R.A.; Smith, M.A.; Robinson, J.T.; Davidovich, C.; Gooding, A.R.; Goodrich, K.J.; Mattick, J.S.; et al. RNA duplex map in living cells reveals higher-order transcriptome structure. Cell 2016, 165, 1267–1279. [Google Scholar] [CrossRef] [PubMed]
- Sharma, E.; Sterne-Weiler, T.; O’Hanlon, D.; Blencowe, B.J. Global mapping of human RNA-RNA interactions. Mol. Cell 2016, 62, 618–626. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Dong, H.; Shi, Y.; Bian, L. Mutually exclusive alternative splicing of pre-mRNAs. Wiley Interdiscip. Rev. RNA 2018, 9, e1468. [Google Scholar] [CrossRef] [PubMed]
- Graveley, B.R. Mutually exclusive splicing of the insect Dscam pre-mRNA directed by competing intronic RNA secondary structures. Cell 2005, 123, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhan, L.; Zhang, W.; Sun, F.; Wang, W.; Tian, N.; Bi, J.; Wang, H.; Shi, D.; Jiang, Y.; et al. RNA secondary structure in mutually exclusive splicing. Nat. Struct. Mol. Biol. 2011, 18, 159–168. [Google Scholar] [CrossRef] [PubMed]
- May, G.E.; Olson, S.; McManus, C.J.; Graveley, B.R. Competing RNA secondary structures are required for mutually exclusive splicing of the Dscam exon 6 cluster. RNA 2011, 17, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Yue, Y.; Li, G.; Yang, Y.; Zhang, W.; Pan, H.; Chen, R.; Shi, F.; Jin, Y. Regulation of Dscam exon 17 alternative splicing by steric hindrance in combination with RNA secondary structures. RNA Biol. 2013, 10, 1822–1833. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, G.; Yang, Y.; Wang, W.; Zhang, W.; Pan, H.; Zhang, P.; Yue, Y.; Lin, H.; Liu, B.; et al. An RNA architectural locus control region involved in Dscam mutually exclusive splicing. Nat. Commun. 2012, 3, 1255. [Google Scholar] [CrossRef] [PubMed]
- Yue, Y.; Meng, Y.; Ma, H.; Hou, S.; Cao, G.; Hong, W.; Shi, Y.; Guo, P.; Liu, B.; Shi, F.; et al. A large family of Dscam genes with tandemly arrayed 5’ cassettes in Chelicerata. Nat. Commun. 2016, 7, 11252. [Google Scholar] [CrossRef] [PubMed]
- Yue, Y.; Yang, Y.; Dai, L.; Cao, G.; Chen, R.; Hong, W.; Liu, B.; Shi, Y.; Meng, Y.; Shi, F.; et al. Long-range RNA pairings contribute to mutually exclusive splicing. RNA 2016, 22, 96–110. [Google Scholar] [CrossRef] [PubMed]
- Yue, Y.; Hou, S.; Wang, X.; Zhan, L.; Cao, G.; Li, G.; Shi, Y.; Zhang, P.; Hong, W.; Lin, H.; et al. Role and convergent evolution of competing RNA secondary structures in mutually exclusive splicing. RNA Biol. 2017, 14, 1399–1410. [Google Scholar] [CrossRef] [PubMed]
- Raker, V.A.; Mironov, A.A.; Gelfand, M.S.; Pervouchine, D.D. Modulation of alternative splicing by long-range RNA structures in Drosophila. Nucleic Acids Res. 2009, 37, 4533–4544. [Google Scholar] [CrossRef] [PubMed]
- Pervouchine, D.D. IRBIS: A systematic search for conserved complementarity. RNA 2014, 20, 1519–1531. [Google Scholar] [CrossRef] [PubMed]
- Rubtsov, P.M. Role of pre-mRNA secondary structures in the regulation of alternative splicing. Mol. Biol. 2016, 50, 935–943. [Google Scholar] [CrossRef]
- Lovci, M.T.; Ghanem, D.; Marr, H.; Arnold, J.; Gee, S.; Parra, M.; Liang, T.Y.; Stark, T.J.; Gehman, L.T.; Hoon, S.; et al. Rbfox proteins regulate alternative mRNA splicing through evolutionarily conserved RNA bridges. Nat. Struct. Mol. Biol. 2013, 20, 1434–1442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, M.S.; Shay, J.W.; Wright, W.E. Regulation of human telomerase splicing by RNA:RNA pairing. Nat. Commun. 2014, 5, 3306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, N.N.; Lawler, M.N.; Ottesen, E.W.; Upreti, D.; Kaczynski, J.R.; Singh, R.N. An intronic structure enabled by a long-distance interaction serves as a novel target for splicing correction in spinal muscular atrophy. Nucleic Acids Res. 2013, 41, 8144–8165. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.N.; Lee, B.M.; Singh, R.N. Splicing regulation in spinal muscular atrophy by an RNA structure formed by long-distance interactions. Ann. N. Y. Acad. Sci. 2015, 1341, 176–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taube, J.R.; Sperle, K.; Banser, L.; Seeman, P.; Cavan, B.C.; Garbern, J.Y.; Hobson, G.M. PMD patient mutations reveal a long-distance intronic interaction that regulates PLP1/DM20 alternative splicing. Hum. Mol. Genet. 2014, 23, 5464–5478. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Schmidt, B.F.; Bruchez, M.P.; McManus, C.J. Structural analyses of NEAT1 lncRNAs suggest long-range RNA interactions that may contribute to paraspeckle architecture. Nucleic Acids Res. 2018, 46, 3742–3752. [Google Scholar] [CrossRef] [PubMed]
- Bernat, V.; Disney, M.D. RNA Structures as mediators of neurological diseases and as drug targets. Neuron 2015, 87, 28–46. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.N.; Howell, M.D.; Androphy, E.J.; Singh, R.N. How the discovery of ISS-N1 led to the first medical therapy for spinal muscular atrophy. Gene Ther. 2017, 24, 520–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umu, S.U.; Gardner, P.P. A comprehensive benchmark of RNA-RNA interaction prediction tools for all domains of life. Bioinformatics 2017, 33, 988–996. [Google Scholar] [CrossRef] [PubMed]
- Lai, D.; Meyer, I.M. A comprehensive comparison of general RNA-RNA interaction prediction methods. Nucleic Acids Res. 2016, 44, e61. [Google Scholar] [CrossRef] [PubMed]
- Wiebe, N.J.; Meyer, I.M. TRANSAT– method for detecting the conserved helices of functional RNA structures, including transient, pseudo-knotted and alternative structures. PLoS Comput. Biol. 2010, 6, e1000823. [Google Scholar] [CrossRef] [PubMed]
- Seemann, S.E.; Richter, A.S.; Gesell, T.; Backofen, R.; Gorodkin, J. PETcofold: Predicting conserved interactions and structures of two multiple alignments of RNA sequences. Bioinformatics 2011, 27, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Bindewald, E.; Shapiro, B.A. Computational detection of abundant long-range nucleotide covariation in Drosophila genomes. RNA 2013, 19, 1171–1182. [Google Scholar] [CrossRef] [PubMed]
- Fricke, M.; Marz, M. Prediction of conserved long-range RNA-RNA interactions in full viral genomes. Bioinformatics 2016, 32, 2928–2935. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, J.S.; Meyer, I.M.; Forsberg, R.; Simmonds, P.; Hein, J. A comparative method for finding and folding RNA secondary structures within protein-coding regions. Nucleic Acids Res. 2004, 32, 4925–4936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, J.S.; Forsberg, R.; Meyer, I.M.; Hein, J. An evolutionary model for protein-coding regions with conserved RNA structure. Mol. Biol. Evol. 2004, 21, 1913–1922. [Google Scholar] [CrossRef] [PubMed]
- Eddy, S.R.; Durbin, R. RNA sequence analysis using covariance models. Nucleic Acids Res. 1994, 22, 2079–2088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, E.I.; Rodionov, D.A. Computational analysis of riboswitch-based regulation. Biochim. Biophys. Acta 2014, 1839, 900–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sankoff, D. Simultaneous solution of the RNA folding, alignment and protosequence problems. SIAM J. Appl. Math. 1985, 45, 810–825. [Google Scholar] [CrossRef]
- Havgaard, J.H.; Torarinsson, E.; Gorodkin, J. Fast pairwise structural RNA alignments by pruning of the dynamical programming matrix. PLoS Comput. Biol. 2007, 3, 1896–1908. [Google Scholar] [CrossRef] [PubMed]
- Will, S.; Yu, M.; Berger, B. Structure-based whole-genome realignment reveals many novel noncoding RNAs. Genome Res. 2013, 23, 1018–1027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatje, K.; Kollmar, M. Expansion of the mutually exclusive spliced exome in Drosophila. Nat. Commun. 2013, 4, 2460. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Sato, K.; Hamada, M.; Watanabe, Y.; Asai, K.; Akutsu, T. RactIP: Fast and accurate prediction of RNA-RNA interaction using integer programming. Bioinformatics 2010, 26, i460–i466. [Google Scholar] [CrossRef] [PubMed]
- Meyer, I.M.; Miklos, I. SimulFold: Simultaneously inferring RNA structures including pseudoknots, alignments, and trees using a Bayesian MCMC framework. PLoS Comput. Biol. 2007, 3, e149. [Google Scholar] [CrossRef] [PubMed]
- Touzet, H.; Perriquet, O. CARNAC: Folding families of related RNAs. Nucleic Acids Res. 2004, 32, W142–W145. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Species | Gene | Function | Length * | Spread * | References |
|---|---|---|---|---|---|
| Drosophila | Dscam | Exon 4 cluster | 13 | 4500 | [31] |
| Dscam | Exon 6 cluster | 16 | 11,000 | [30,32,34] | |
| Dscam | Exon 9 cluster | 16 | 14,000 | [31] | |
| Dscam | Exon 17 cluster | 15 | 1000 | [33] | |
| Mhc | Exon 7 cluster | 14 | 2500 | [31] | |
| Mhc | Exon 9 cluster | 14 | 1600 | [31] | |
| Mhc | Exon 11 cluster | 15 | 2600 | [31] | |
| Nmnat | Exon 5 and polyA site | 14 | 400 | [38] | |
| Atrophin | Exon 10 | 16 | 350 | [38] | |
| srp | Exon 4 cluster | 21 | 450 | [36] | |
| 14-3-3ζ | Exon 5 cluster | 22 | 1200 | [31] | |
| Human | SF1 | Exon 10 | 17 | 100 | [20] |
| ENAH | Exon 11a | 18 | 1800 | [41] | |
| DST | Exons 47-52 | 15 | 10,000 | [39] | |
| SMN2 | Exon 7 | 8 + 7 + 8 | 280 | [43,44] | |
| PLP1 | Exon 3 | 10 + 5 | 600 | [45] | |
| TERT | Exons 7 and 8 | Repeat | 6500 | [42] | |
| NEAT1 | Paraspeckle formation | N/A | 10,000 | [46] |
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pervouchine, D.D. Towards Long-Range RNA Structure Prediction in Eukaryotic Genes. Genes 2018, 9, 302. https://doi.org/10.3390/genes9060302
Pervouchine DD. Towards Long-Range RNA Structure Prediction in Eukaryotic Genes. Genes. 2018; 9(6):302. https://doi.org/10.3390/genes9060302
Chicago/Turabian StylePervouchine, Dmitri D. 2018. "Towards Long-Range RNA Structure Prediction in Eukaryotic Genes" Genes 9, no. 6: 302. https://doi.org/10.3390/genes9060302
APA StylePervouchine, D. D. (2018). Towards Long-Range RNA Structure Prediction in Eukaryotic Genes. Genes, 9(6), 302. https://doi.org/10.3390/genes9060302