Possible Role of Envelope Components in the Extreme Copper Resistance of the Biomining Acidithiobacillus ferrooxidans



Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Preparation of Total Protein Extracts for iTRAQ Analysis
2.3. Protein Digestion and Tagging with iTRAQ-8-Plex® Reagent
2.4. Liquid Chromatography and Mass Spectrometry Analysis
2.5. Data Analysis and Statistics
2.6. Extraction of Total RNA from Acidithiobacillus ferrooxidans and Complementary DNA Synthesis
2.7. Primer Design, Real-Time PCR and Cloning of A. ferrooxidans Genes
2.8. Cloning A. ferrooxidans Genes in an Expression Vector
2.9. Lipopolysaccharide Extraction
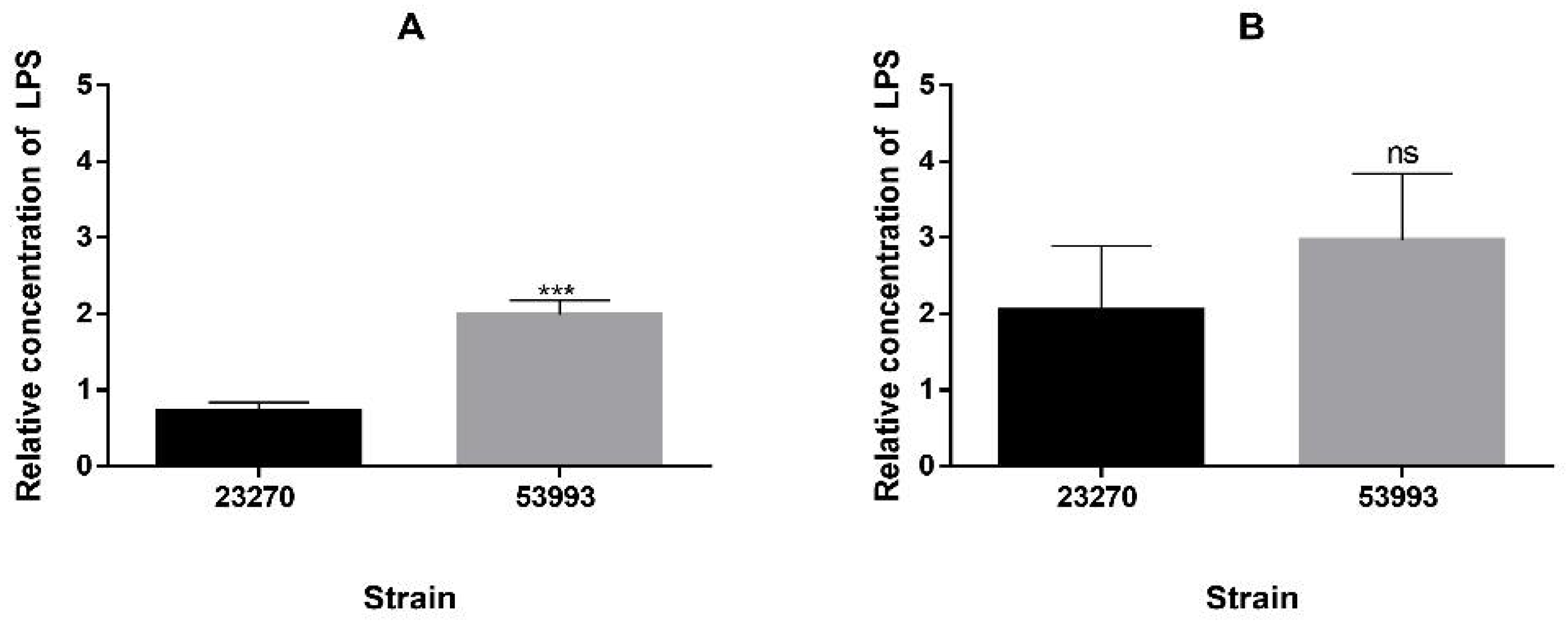
2.10. Lipopolysaccharide Quantification
3. Results and Discussion
3.1. Proteomic Analysis of the Copper Response of A. ferrooxidans ATCC 53993
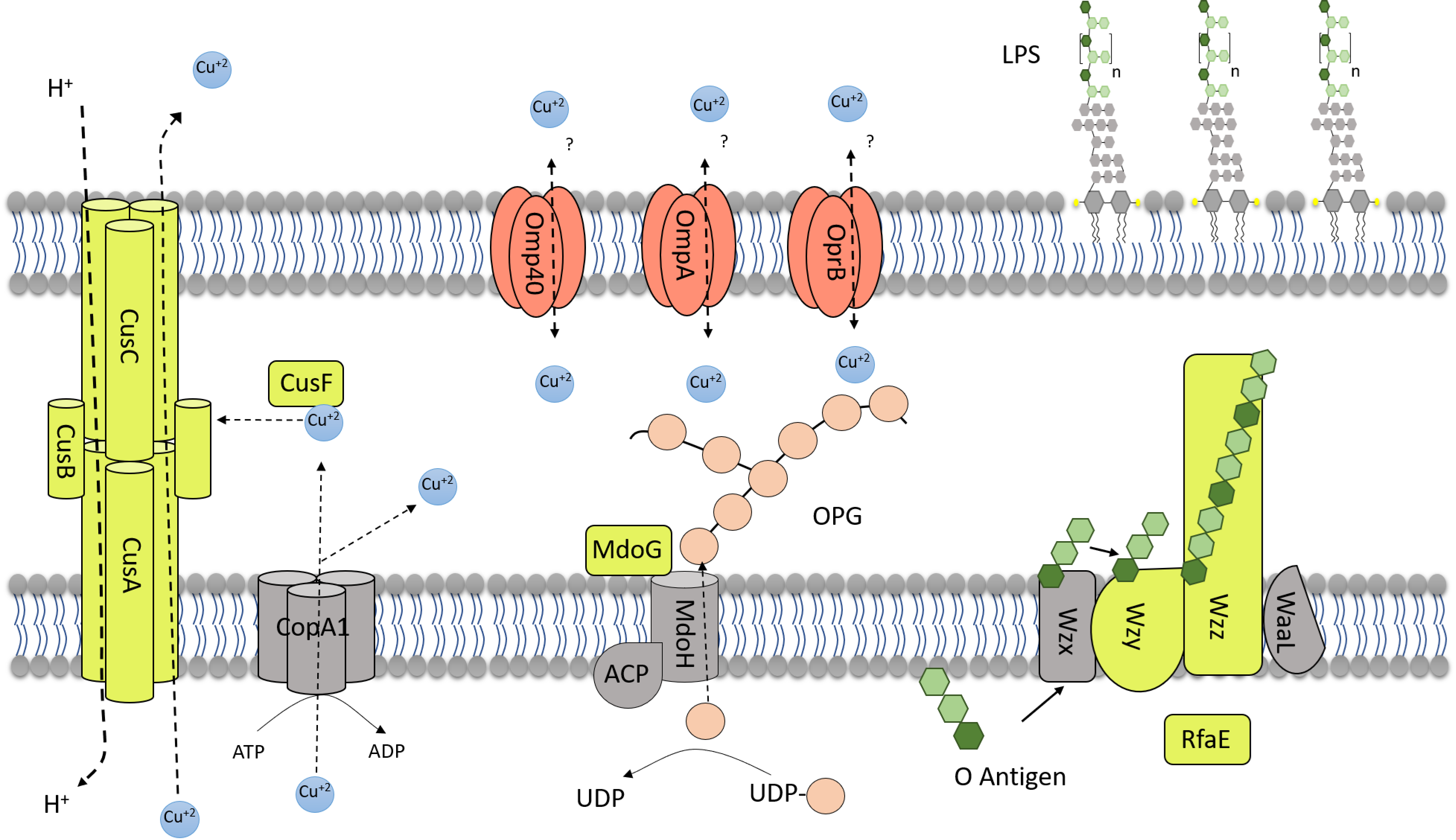
3.2. Overexpression of Resistance-Nodulation-Division Efflux Transporters and Possible Generation of Excess Acidity
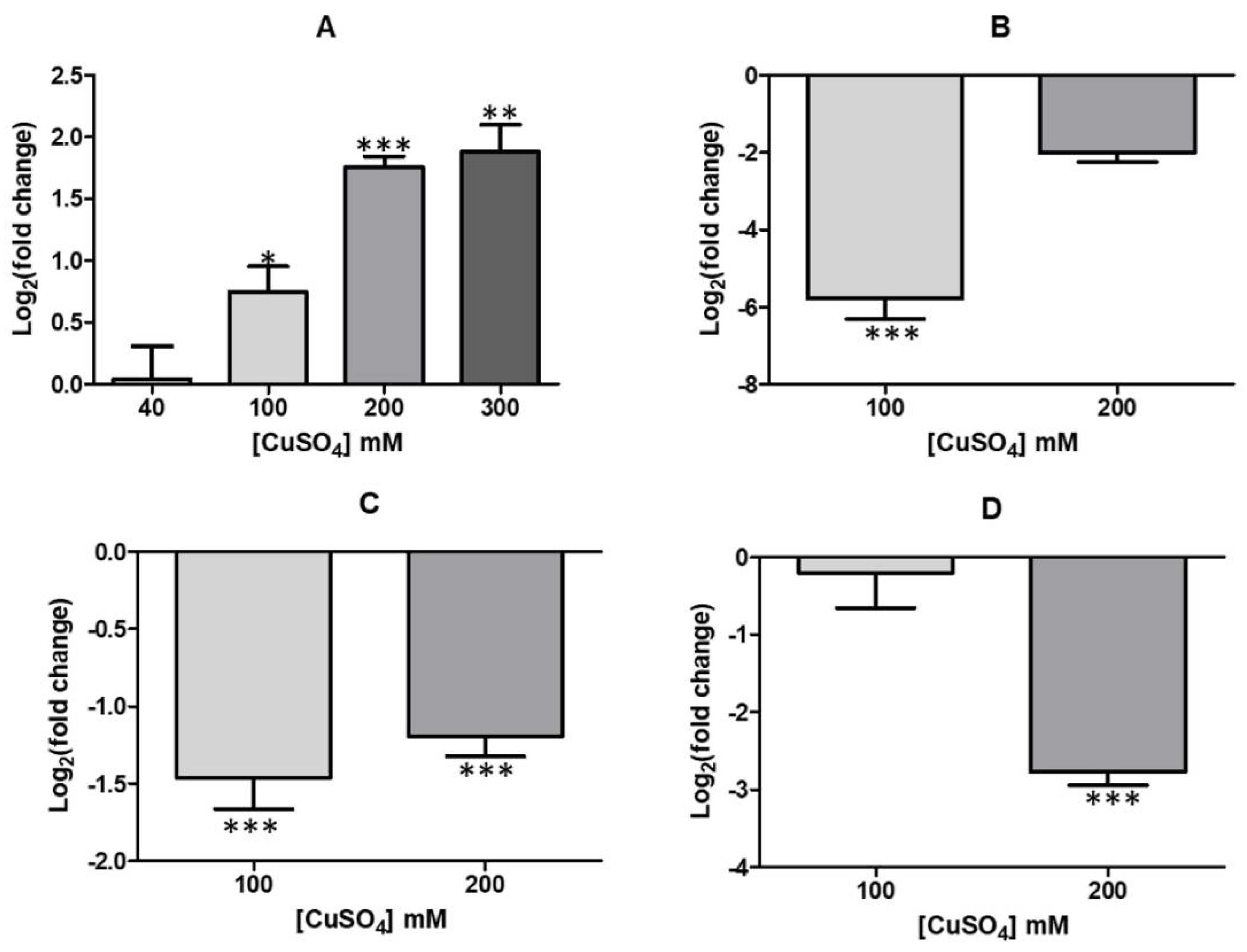
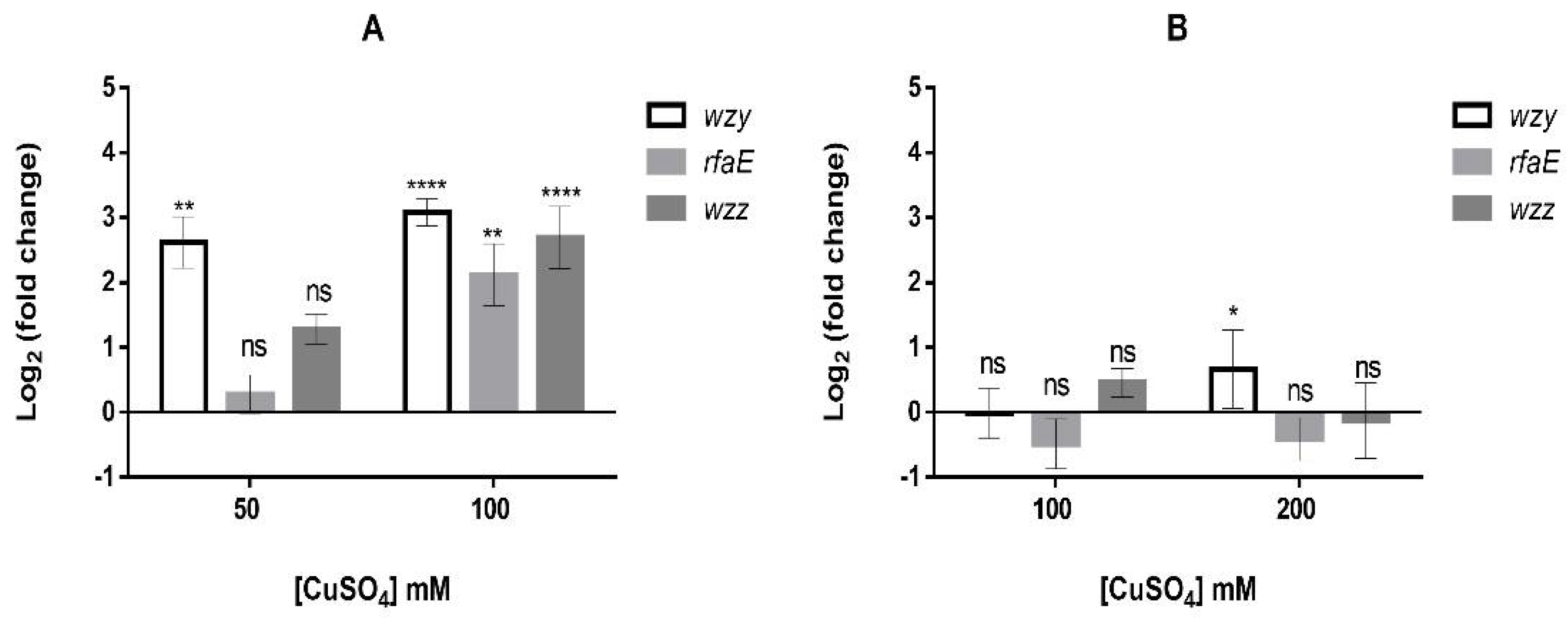
3.3. Changes of Several Additional Envelope Components Occur in Presence of High Copper Concentrations
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vera, M.; Schippers, A.; Sand, W. Progress in bioleaching: Fundamentals and mechanisms of bacterial metal sulfide oxidation—Part A. Appl. Microbiol. Biotechnol. 2013, 97, 7529–7541. [Google Scholar] [CrossRef] [PubMed]
- Rawlings, D.E. Characteristics and adaptability of iron- and sulfur-oxidizing microorganisms used for the recovery of metals from minerals and their concentrates. Microb. Cell Fact. 2005, 4, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valenzuela, L.; Chi, A.; Beard, S.; Orell, A.; Guiliani, N.; Shabanowitz, J.; Hunt, D.F.; Jerez, C.A. Genomics, metagenomics and proteomics in biomining microorganisms. Biotechnol. Adv. 2006, 24, 197–211. [Google Scholar] [CrossRef] [PubMed]
- Jerez, C.A. The use of genomics, proteomics and other OMICS technologies for the global understanding of biomining microorganisms. Hydrometallurgy 2008, 94, 162–169. [Google Scholar] [CrossRef]
- Jerez, C.A. Bioleaching and biomining for the industrial recovery of metals. Compr. Biotechnol. 2011, 3, 717–729. [Google Scholar]
- Navarro, C.A.; von Bernath, D.; Jerez, C.A. Heavy metal resistance strategies of acidophilic bacteria and their acquisition: Importance for biomining and bioremediation. Biol. Res. 2013, 46, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Watling, H.R. The bioleaching of sulphide minerals with emphasis on copper sulphides—A review. Hydrometallurgy 2006, 84, 81–108. [Google Scholar] [CrossRef]
- Watkin, E.L.J.; Keeling, S.E.; Perrot, F.A.; Shiers, D.W.; Palmer, M.L.; Watling, H.R. Metals tolerance in moderately thermophilic isolates from a spent copper sulfide heap, closely related to Acidithiobacillus caldus, Acidimicrobium ferrooxidans and Sulfobacillus thermosulfidooxidans. J. Ind. Microbiol. Biotechnol. 2009, 36, 461–465. [Google Scholar] [CrossRef] [PubMed]
- Jerez, C.A. Biomining of metals: How to access and exploit natural resource sustainably. Microb. Biotechnol. 2017, 10, 1191–1193. [Google Scholar] [CrossRef] [PubMed]
- Franke, S.; Grass, G.; Rensing, C.; Nies, D.H. Molecular analysis of the copper-transporting efflux system CusCFBA of Escherichia coli. J. Bacteriol. 2003, 185, 3804–3812. [Google Scholar] [CrossRef] [PubMed]
- Padilla-Benavides, T.; Thompson, A.M.G.; McEvoy, M.M.; Argüello, J.M. Mechanism of ATPase-mediated Cu+ export and delivery to periplasmic chaperones: The interaction of Escherichia coli CopA and CusF. J. Biol. Chem. 2014, 289, 20492–20501. [Google Scholar] [CrossRef] [PubMed]
- Argüello, J.M.; Raimunda, D.; Padilla-Benavides, T. Mechanisms of copper homeostasis in bacteria. Front. Cell. Infect. Microbiol. 2013, 3, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Cárdenas, J.P.; Quatrini, R.; Holmes, D.S. Genomic and metagenomic challenges and opportunities for bioleaching: A mini-review. Res. Microbiol. 2016, 167, 529–538. [Google Scholar] [CrossRef] [PubMed]
- Orellana, L.H.; Jerez, C.A. A genomic island provides Acidithiobacillus ferrooxidans ATCC 53993 additional copper resistance: A possible competitive advantage. Appl. Microbiol. Biotechnol. 2011, 92, 761–767. [Google Scholar] [CrossRef] [PubMed]
- Remonsellez, F.; Orell, A.; Jerez, C.A. Copper tolerance of the thermoacidophilic archaeon Sulfolobus metallicus: Possible role of polyphosphate metabolism. Microbiology 2006, 152, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Dopson, M.; Holmes, D.S. Metal resistance in acidophilic microorganisms and its significance for biotechnologies. Appl. Microbiol. Biotechnol. 2014, 98, 8133–8144. [Google Scholar] [CrossRef] [PubMed]
- Orell, A.; Navarro, C.A.; Arancibia, R.; Mobarec, J.C.; Jerez, C.A. Life in blue: Copper resistance mechanisms of bacteria and Archaea used in industrial biomining of minerals. Biotechnol. Adv. 2010, 28, 839–848. [Google Scholar] [CrossRef] [PubMed]
- Orell, A.; Navarro, C.A.; Rivero, M.; Aguilar, J.S.; Jerez, C.A. Inorganic polyphosphates in extremophiles and their possible functions. Extremophiles 2012, 16, 573–583. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Bussenius, C.; Navarro, C.A.; Jerez, C.A. Microbial copper resistance: Importance in biohydromrtallurgy. Microb. Biotechnol. 2017, 10, 279–295. [Google Scholar] [CrossRef] [PubMed]
- Dopson, M.; Ossandon, F.J.; Lövgren, L.; Holmes, D. Metal resistance or tolerance? Acidophiles confront high metal loads via both abiotic and biotic mechanisms. Front. Microbiol. 2014, 5, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Almárcegui, R.J.; Navarro, C.A.; Paradela, A.; Albar, J.P.; von Bernath, D.; Jerez, C.A. New copper resistance determinants in the extremophile Acidithiobacillus ferrooxidans: A quantitative proteomic analysis. J. Proteome Res. 2014, 7, 946–960. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Bussenius, C.; Navarro, C.A.; Orellana, L.; Paradela, A.; Jerez, C.A. Global response of Acidithiobacillus ferrooxidans ATCC 53993 to high concentrations of copper: A quantitative proteomics approach. J. Proteom. 2016, 145, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Amaro, A.M.; Chamorro, D.; Seeger, M.; Arredondo, R. Effect of external pH perturbations on in vivo protein synthesis by the acidophilic bacterium Thiobacillus ferrooxidans. J. Bacteriol. 1991, 173, 910–915. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Fernández, A.; Paradela, A.; Navajas, R.; Albar, J.P. Generalized method for probability-based peptide and protein identification from tandem mass spectrometry data and sequencue database searching. Mol. Cell. Proteom. 2008, 7, 1748–1754. [Google Scholar] [CrossRef] [PubMed]
- López-Serra, P.; Marcilla, M.; Villanueva, A.; Ramos-Fernández, A.; Palau, A.; Leal, L.; Wahi, J.E.; Setien-Baranda, F.; Szczesna, K.; Moutinho, C.; et al. A DERL3-associated defect in the degradation of SLC2A1 mediates the Warburg effect. Nat. Commun. 2014, 5, 3608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vera, M.; Pagliai, F.; Guiliani, N.; Jerez, C.A. The chemolithoautotroph Acidithiobacillus ferrooxidans can survive under phosphate-limiting conditions by expressing a C-P lyase operon that allows it to grow on phosphonates. Appl. Environ. Microbiol. 2008, 74, 1829–1835. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, S.; Jerez, C.A. Copper ions stimulate polyphosphate degradation and phosphate efflux in Acidithiobacillus ferrooxidans. Appl. Environ. Microbiol. 2004, 70, 5177–5182. [Google Scholar] [CrossRef] [PubMed]
- Navarro, C.A.; Orellana, L.H.; Mauriaca, C.; Jerez, C.A.; Navarro, C.A.; Orellana, L.H.; Mauriaca, C.; Jerez, C.A. Transcriptional and functional studies of Acidithiobacillus ferrooxidans genes related to survival in the presence of copper. Appl. Environ. Microbiol. 2009, 75, 6102–6109. [Google Scholar] [CrossRef] [PubMed]
- Rozen, S.; Skaletsky, H. Primer3 on the WWW for general users and for biologist programmers. Methods Mol. Biol. 2000, 132, 365–386. [Google Scholar] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Hitchcock, P.J.; Brown, T.M. Morphological heterogeneity among Salmonella lipopolysaccharide chemotypes in silver-stained polyacrylamide gels. J. Bacteriol. 1983, 154, 269–277. [Google Scholar] [PubMed]
- Quesenberry, M.S.; Lee, Y.C. A rapid formaldehyde assay using purpald reagent: Application under periodation conditions. Anal. Biochem. 1996, 234, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Tsai, C.M. Quantification of bacterial lipopolysaccharides by the purpald assay: Measuring formaldehyde generated from 2-keto-3-deoxyoctonate and heptose at the inner core by periodate oxidation. Anal. Biochem. 1999, 267, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Baker-Austin, C.; Dopson, M. Life in acid: pH homeostasis in acidophiles. Trends Microbiol. 2007, 15, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Quatrini, R.; Appia-Ayme, C.; Denis, Y.; Jedlicki, E.; Holmes, D.S.; Bonnefoy, V. Extending the models for iron and sulfur oxidation in the extreme acidophile Acidithiobacillus ferrooxidans. BMC Genom. 2009, 10, 394. [Google Scholar] [CrossRef] [PubMed]
- Bohin, J.P. Osmoregulated periplasmic glucans in Proteobacteria. FEMS Microbiol. Lett. 2000, 186, 11–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chi, A.; Valenzuela, L.; Beard, S.; Mackey, A.J.; Shabanowitz, J.; Hunt, D.F.; Jerez, C.A. Periplasmic proteins of the extremophile Acidithiobacillus ferrooxidans. Mol. Cell. Proteom. 2007, 6, 2239–2251. [Google Scholar] [CrossRef] [PubMed]
- Bontemps-Gallo, S.; Bohin, J.P.; Lacroix, J.M. Osmoregulated periplasmic glucans. EcoSal Plus 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Weissborn, A.C.; Liu, Q.; Rumley, M.K.; Kennedy, E.P. UTP-D-glucose-1-phosphate uridylyltransferase of Escherichia coli: Isolation and DNA sequence of the galU gene and purification of the enzyme. J. Bacteriol. 1994, 176, 2611–2618. [Google Scholar] [CrossRef] [PubMed]
- Delcour, A.H.; Adler, J.; Kung, C.; Martinac, B. Membrane-derived oligosaccharides (MDO’s) promote closing of an E. coli porin channel. FEBS Lett. 1992, 304, 216–220. [Google Scholar] [CrossRef]
- Fiedler, W.; Rotering, H. Properties of Escherichia coli mutants lacking membrane-derived oligosaccharides. J. Biol. Chem. 1988, 263, 14684–14689. [Google Scholar] [PubMed]
- Speer, A.; Rowland, J.L.; Haeili, M.; Niederweis, M. Porins increase copper susceptibility of Mycobacterium tuberculosis. J. Bacteriol. 2013, 195, 5133–5140. [Google Scholar] [CrossRef] [PubMed]
- Gumulya, Y.; Boxall, N.J.; Khaleque, H.N.; Santala, V.; Carlson, R.P.; Kaksonen, A.H. In a quest for engineering acidophiles for biomining applications: Challenges and opportunities. Genes 2018, 9, 116. [Google Scholar] [CrossRef] [PubMed]
- Pisani, F.; Italiano, F.; De Leo, F.; Gallerani, R.; Rinalducci, S.; Zolla, L.; Agostiano, A.; Ceci, L.R.; Trotta, M. Soluble proteome investigation of cobalt effect on the carotenoidless mutant of Rhodobacter sphaeroides. J. Appl. Microbiol. 2009, 106, 338–349. [Google Scholar] [CrossRef] [PubMed]
- Lerouge, I.; Vanderleyden, J. O-antigen structural variation: Mechanisms and possible roles in animal/plant-microbe interactions. FEMS Microbiol. Rev. 2001, 26, 17–47. [Google Scholar] [CrossRef]
- Snyder, D.S.; Brahamsha, B.; Azadi, P.; Palenik, B.; Acteriol, J.B. Structure of compositionally simple lipopolysaccharide from marine Synechococcus. J. Bacteriol. 2009, 191, 5499–5509. [Google Scholar] [CrossRef] [PubMed]
- Langley, T.; Beveridge, T.J. Effect of O-side-chain-lipopolysaccharide chemistry on metal binding. Appl. Environ. Microbiol. 1999, 65, 489–498. [Google Scholar] [PubMed]
- Vilinska, A.; Rao, K.H. Surface characterization of Acidithiobacillus ferrooxidans adapted to high copper and zinc ions concentration. Geomicrobiol. J. 2011, 28, 221–228. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Function/Similarity | ORF | Name | q Value (FDR) | Coverage (%) | Peptide Number | Log2 Fold Change (Cu 200/0 mM) |
|---|---|---|---|---|---|---|
| Outer membrane efflux protein | Lferr_1619 | CusC1 | 0.001 | 45 | 9 | 1.258 |
| Efflux transporter, RND family, MFP subunit | Lferr_1618 | CusB1 | 0.001 | 63.3 | 15 | 0.859 |
| Uncharacterized protein | Lferr_2057 | CusF2 | 0.001 | 60 | 3 | 1.92 |
| Uncharacterized protein | Lferr_0174 | CusF3 | 0.001 | 60 | 3 | 1.63 |
| Heavy metal efflux pump, CzcA family | Lferr_0172 | CusA3 | 0.001 | 39.1 | 10 | 1.019 |
| Outer membrane efflux protein | Lferr_2062 | CusC2 | 0 | 45 | 12 | 1.084 |
| Heavy metal efflux pump, CzcA | Lferr_1617 | CusA1 | 0.002 | 35 | 9 | 0.855 |
| Efflux transporter, RND family, MFP subunit | Lferr_2061 | CusB2 | 0.003 | 68.3 | 6 | 0.968 |
| Heavy metal efflux pump, CzcA family | Lferr_2060 | CusA2 | 0.003 | 38.3 | 9 | 0.846 |
| Periplasmic glucan biosynthesis protein MdoG | Lferr_1075 | MdoG | 0.009 | 48.1 | 15 | 0.415 |
| Carbohydrate-selective porin OprB | Lferr_1898 | OprB | 0.005 | 36.46 | 10 | −0.635 |
| O-antigen polymerase | Lferr_0408 | Wzy | 0.026 | 3.19 | 1 | 1.795 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oetiker, N.; Norambuena, R.; Martínez-Bussenius, C.; Navarro, C.A.; Amaya, F.; Álvarez, S.A.; Paradela, A.; Jerez, C.A. Possible Role of Envelope Components in the Extreme Copper Resistance of the Biomining Acidithiobacillus ferrooxidans. Genes 2018, 9, 347. https://doi.org/10.3390/genes9070347
Oetiker N, Norambuena R, Martínez-Bussenius C, Navarro CA, Amaya F, Álvarez SA, Paradela A, Jerez CA. Possible Role of Envelope Components in the Extreme Copper Resistance of the Biomining Acidithiobacillus ferrooxidans. Genes. 2018; 9(7):347. https://doi.org/10.3390/genes9070347
Chicago/Turabian StyleOetiker, Nia, Rodrigo Norambuena, Cristóbal Martínez-Bussenius, Claudio A. Navarro, Fernando Amaya, Sergio A. Álvarez, Alberto Paradela, and Carlos A. Jerez. 2018. "Possible Role of Envelope Components in the Extreme Copper Resistance of the Biomining Acidithiobacillus ferrooxidans" Genes 9, no. 7: 347. https://doi.org/10.3390/genes9070347
APA StyleOetiker, N., Norambuena, R., Martínez-Bussenius, C., Navarro, C. A., Amaya, F., Álvarez, S. A., Paradela, A., & Jerez, C. A. (2018). Possible Role of Envelope Components in the Extreme Copper Resistance of the Biomining Acidithiobacillus ferrooxidans. Genes, 9(7), 347. https://doi.org/10.3390/genes9070347