Integrase-Controlled Excision of Metal-Resistance Genomic Islands in Acinetobacter baumannii


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Cultivation
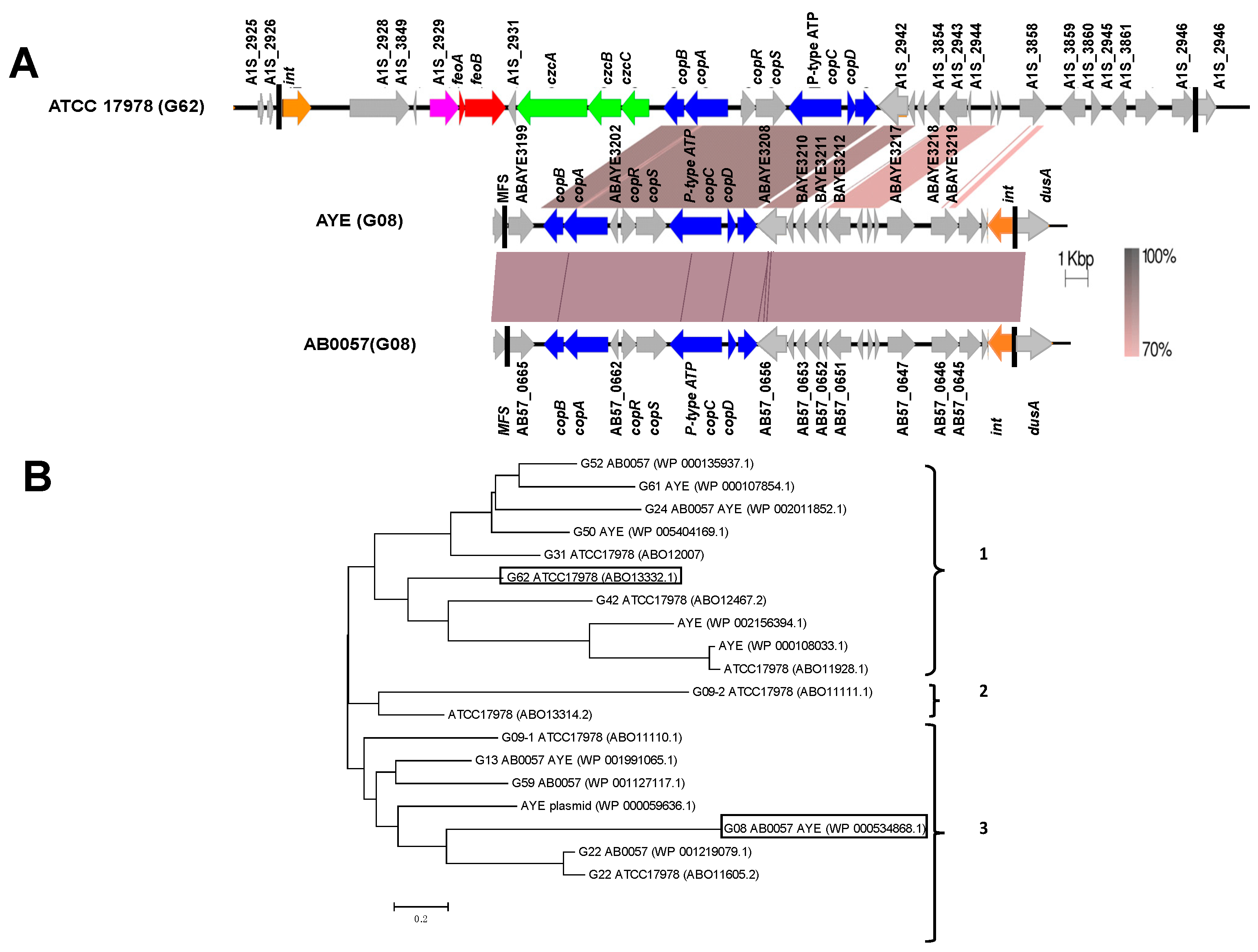
2.2. Genome Analysis
2.3. Colony Genotpying
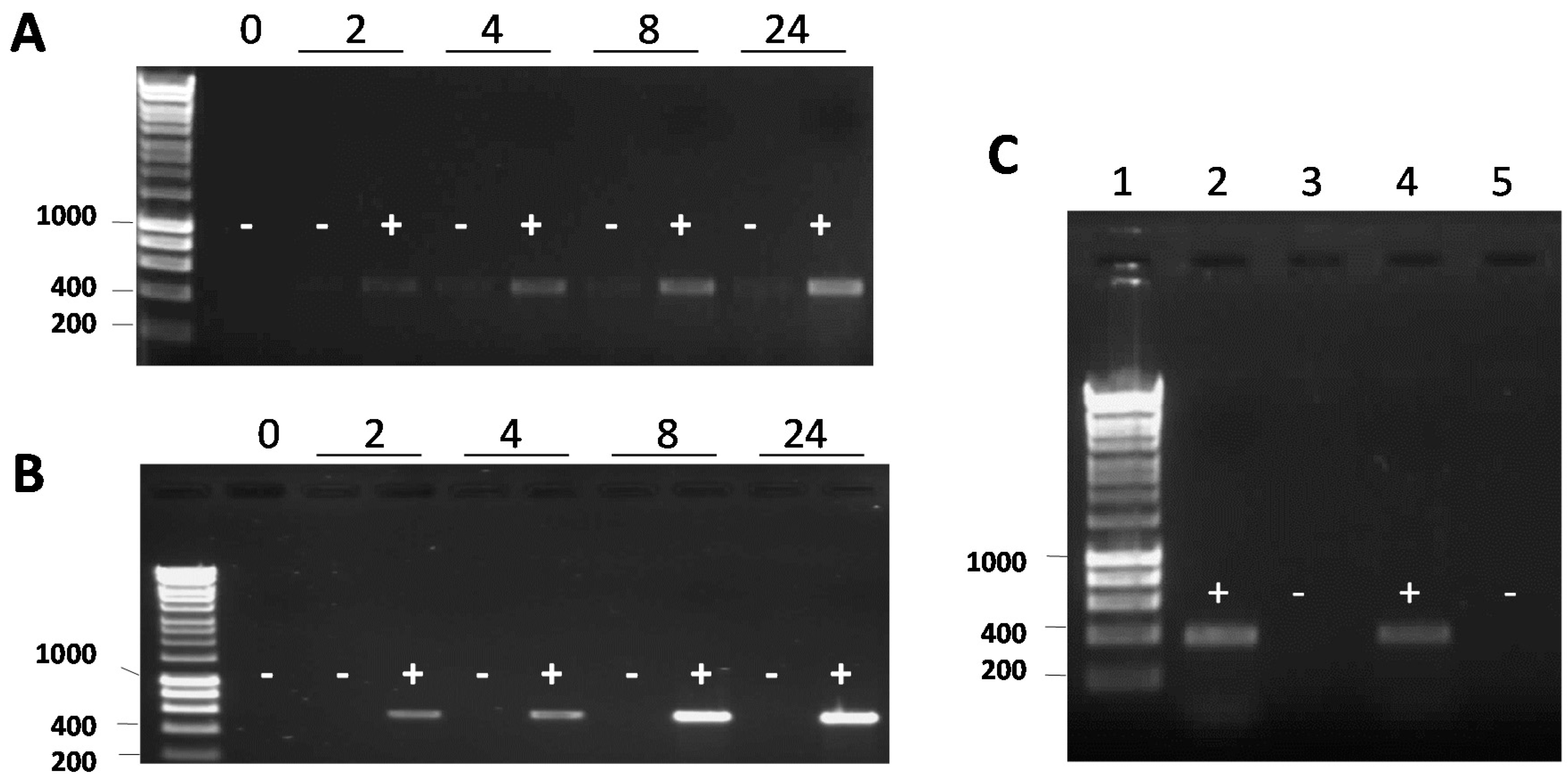
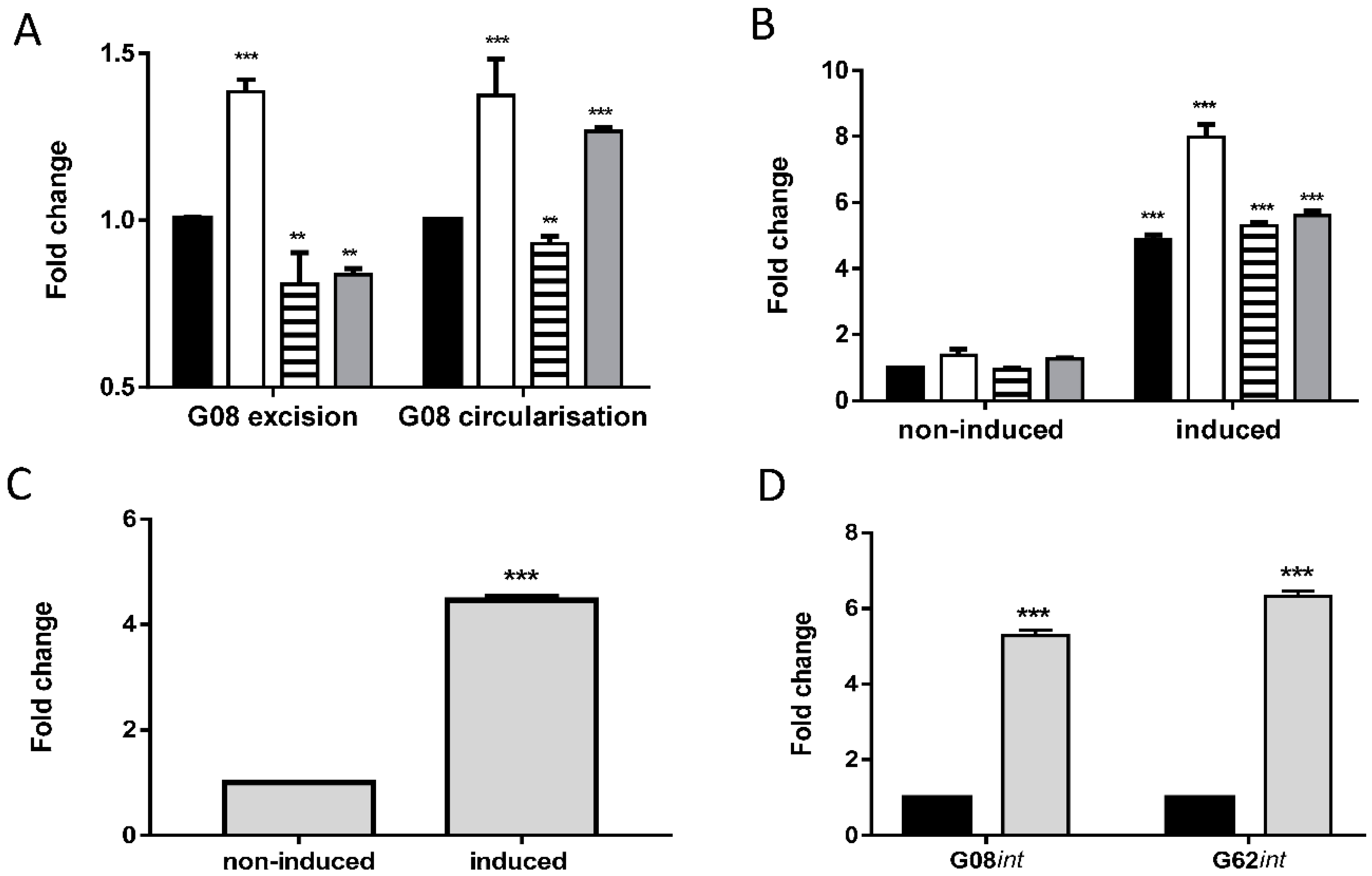
2.4. Real-Time PCR
2.5. RNA Extraction and Retrotranscription
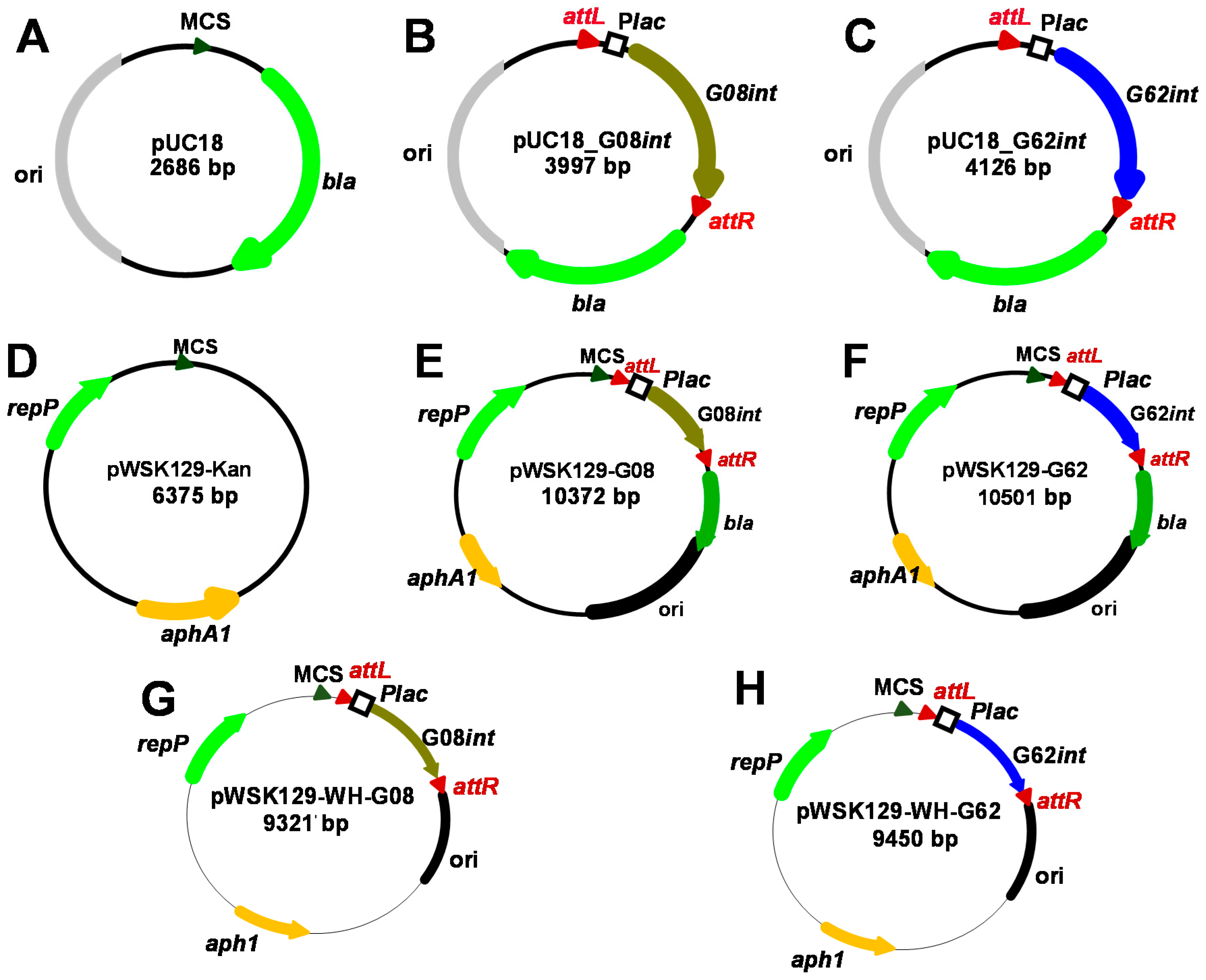
2.6. Construction of Plasmids with G08 and G62 Mini-GIs
2.7. Construction of Inducible Plasmids for A. baumannii
2.8. Suicide Vector-Based Allelic Exchange for Mutant Construction in A. baumannii
2.9. IPTG and Mitomycin C Induction
2.10. Metal Susceptibility Testing
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hacker, J.; Blum-Oehler, G.; Muhldorfer, I.; Tschape, H. Pathogenicity islands of virulent bacteria: Structure, function and impact on microbial evolution. Mol. Microbiol. 1997, 23, 1089–1097. [Google Scholar] [CrossRef] [PubMed]
- Juhas, M.; van der Meer, J.R.; Gaillard, M.; Harding, R.M.; Hood, D.W.; Crook, D.W. Genomic islands: Tools of bacterial horizontal gene transfer and evolution. FEMS Microbiol. Rev. 2009, 33, 376–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchrieser, C.; Brosch, R.; Bach, S.; Guiyoule, A.; Carniel, E. The high-pathogenicity island of Yersinia pseudotuberculosis can be inserted into any of the three chromosomal asn tRNA genes. Mol. Microbiol. 1998, 30, 965–978. [Google Scholar] [CrossRef] [PubMed]
- Gal-Mor, O.; Finlay, B.B. Pathogenicity islands: A molecular toolbox for bacterial virulence. Cell. Microbiol. 2006, 8, 1707–1719. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, W.W.L.; Ung, K.; Aeschliman, D.; Bryan, J.; Finlay, B.B.; Brinkman, F.S.L. Evidence of a large novel gene pool associated with prokaryotic genomic islands. PLoS Genet. 2005, 1, e62. [Google Scholar] [CrossRef] [PubMed]
- Peleg, A.Y.; de Breij, A.; Adams, M.D.; Cerqueira, G.M.; Mocali, S.; Galardini, M.; Nibbering, P.H.; Earl, A.M.; Ward, D.V.; Paterson, D.L.; et al. The success of Acinetobacter species; genetic, metabolic and virulence attributes. PLoS ONE 2012, 7, e46984. [Google Scholar] [CrossRef] [PubMed]
- Camp, C.; Tatum, O.L. A Review of Acinetobacter baumannii as a highly successful pathogen in times of war. Lab. Med. 2010, 41, 649–657. [Google Scholar] [CrossRef]
- Bergogne-Bérézin, E.; Friedman, H.; Bendinelli, M. (Eds.) Acinetobacter Biology and Pathogenesis; Springer: New York, NY, USA, 2008; 220p. [Google Scholar]
- Amat, T.; Gutiérrez-Pizarraya, A.; Machuca, I.; Gracia-Ahufinger, I.; Pérez-Nadales, E.; Torre-Giménez, A.; Garnacho-Montero, J.; Cisneros, J.M.; Torre-Cisneros, J. The combined use of tigecycline with high-dose colistin might not be associated with higher survival in critically ill patients with bacteraemia due to carbapenem-resistant Acinetobacter baumannii. Clin. Microbiol. Infect. 2018, 24, 630–634. [Google Scholar] [CrossRef] [PubMed]
- Fournier, P.E.; Vallenet, D.; Barbe, V.; Audic, S.; Ogata, H.; Poirel, L.; Richet, H.; Robert, C.; Mangenot, S.; Abergel, C.; et al. Comparative genomics of multidrug resistance in Acinetobacter baumannii. PLoS Genet. 2006, 2, e7. [Google Scholar] [CrossRef] [PubMed]
- Kochar, M.; Crosatti, M.; Harrison, E.M.; Rieck, B.; Chan, J.; Constantinidou, C.; Pallen, M.; Ou, H.Y.; Rajakumar, K. Deletion of TnAbaR23rResults in both expected and unexpected antibiogram changes in a multidrug-resistant Acinetobacter baumannii strain. Antimicrob. Agents Chemother. 2012, 56, 1845–1853. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Yan, Z.; Zhang, Z.; Zhou, Q.; Zhou, J.; Wakeland, E.K.; Fang, X.; Xuan, Z.; Shen, D.; Li, Q.-Z. Complete genome analysis of three Acinetobacter baumannii clinical isolates in China for insight into the diversification of drug resistance elements. PLoS ONE 2013, 8, e66584. [Google Scholar] [CrossRef] [PubMed]
- Post, V.; Hall, R.M. AbaR5, a large multiple-antibiotic resistance region found in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2009, 53, 2667–2671. [Google Scholar] [CrossRef] [PubMed]
- Di Nocera, P.P.; Rocco, F.; Giannouli, M.; Triassi, M.; Zarrilli, R. Genome organization of epidemic Acinetobacter baumannii strains. BMC Microbiol. 2011, 11, 224. [Google Scholar] [CrossRef] [PubMed]
- Ou, H.Y.; Kuang, S.N.; He, X.; Molgora, B.M.; Ewing, P.J.; Deng, Z.; Osby, M.; Chen, W.; Xu, H.H. Complete genome sequence of hypervirulent and outbreak-associated Acinetobacter baumannii strain LAC-4: Epidemiology, resistance genetic determinants and potential virulence factors. Sci. Rep. 2015, 5, 8643. [Google Scholar] [CrossRef] [PubMed]
- Hassan, K.A.; Pederick, V.G.; Elbourne, L.D.; Paulsen, I.T.; Paton, J.C.; McDevitt, C.A.; Eijkelkamp, B.A. Zinc stress induces copper depletion in Acinetobacter baumannii. BMC Microbiol. 2017, 17, 59. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.G.; Gianoulis, T.A.; Pukatzki, S.; Mekalanos, J.J.; Ornston, L.N.; Gerstein, M.; Snyder, M. New insights into Acinetobacter baumannii pathogenesis revealed by high-density pyrosequencing and transposon mutagenesis. Genes Dev. 2007, 21, 601–614. [Google Scholar] [CrossRef] [PubMed]
- Eijkelkamp, B.; Stroeher, U.; Hassan, K.; Paulsen, I.; Brown, M. Comparative analysis of surface-exposed virulence factors of Acinetobacter baumannii. BMC Genom. 2014, 15, 1020. [Google Scholar] [CrossRef] [PubMed]
- Farrugia, D.N.; Elbourne, L.D.; Mabbutt, B.C.; Paulsen, I.T. A novel family of integrases associated with prophages and genomic islands integrated within the tRNA-dihydrouridine synthase A (dusA) gene. Nucleic Acids Res. 2015, 43, 4547–4557. [Google Scholar] [CrossRef] [PubMed]
- Croucher, N.J.; Coupland, P.G.; Stevenson, A.E.; Callendrello, A.; Bentley, S.D.; Hanage, W.P. Diversification of bacterial genome content through distinct mechanisms over different timescales. Nat. Commun. 2014, 5, 5471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A genome comparison visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nie, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Adams, M.D.; Goglin, K.; Molyneaux, N.; Hujer, K.M.; Lavender, H.; Jamison, J.J.; MacDonald, I.J.; Martin, K.M.; Russo, T.; Campagnari, A.A.; et al. Comparative genome sequence analysis of multidrug-resistant Acinetobacter baumannii. J. Bacteriol. 2008, 190, 8053–8064. [Google Scholar] [CrossRef] [PubMed]
- Diancourt, L.; Passet, V.; Nemec, A.; Dijkshoorn, L.; Brisse, S. The population structure of Acinetobacter baumannii: expanding multiresistant clones from an ancestral susceptible genetic pool. PLoS ONE 2010, 5, e10034. [Google Scholar] [CrossRef] [PubMed]
- van Vliet, A.H.; Kusters, J.G. Use of alignment-free phylogenetics for rapid genome sequence-based typing of Helicobacter pylori virulence markers and antibiotic susceptibility. J. Clin. Microbiol. 2015, 53, 2877–2888. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.F.; Kushner, S.R. Construction of versatile low-copy number vectors for cloning, sequencing and gene expression in Escherichia coli. Gene 1991, 100, 195–199. [Google Scholar] [CrossRef]
- van Aartsen, J.J.; Rajakumar, K. An optimized method for suicide vector-based allelic exchange in Klebsiella pneumoniae. J. Microbiol. Methods 2011, 86, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Ubeda, C.; Maiques, E.; Knecht, E.; Lasa, I.; Novick, R.P.; Penades, J.R. Antibiotic-induced SOS response promotes horizontal dissemination of pathogenicity island-encoded virulence factors in staphylococci. Mol. Microbiol. 2005, 56, 836–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barry, A.L.; Craig, W.A.; Nadler, H.; Reller, L.B.; Sanders, C.C.; Swenson, J.M. Methods for determining bactericidal activity of antimicrobial agents; approved guideline. NCCLS 1999, 19, M26-A. [Google Scholar]
- Yanisch-Perron, C.; Vierira, J.; Messing, J. Improved M13 phage cloning vectors and host strains: Nucleotide sequences of the M13mpI8 and pUC19 vectors. Gene 1985, 33, 103–119. [Google Scholar] [CrossRef]
- Chan, A.P.; Sutton, G.; DePew, J.; Krishnakumar, R.; Choi, Y.; Huang, X.Z.; Beck, E.; Harkins, D.M.; Kim, M.; Lesho, E.P.; et al. A novel method of consensus pan-chromosome assembly and large-scale comparative analysis reveal the highly flexible pan-genome of Acinetobacter baumannii. Genome Biol. 2015, 16, 143. [Google Scholar] [CrossRef] [PubMed]
- Nemergut, D.R.; Robeson, M.S.; Kysela, R.F.; Martin, A.P.; Schmidt, S.K.; Knight, R. Insights and inferences about integron evolution from genomic data. BMC Genom. 2008, 9, 261. [Google Scholar] [CrossRef] [PubMed]
- Gillings, M.R.; Holley, M.P.; Stokes, H.W.; Holmes, A.J. Integrons in Xanthomonas: A source of species genome diversity. Proc. Natl. Acad. Sci. USA 2005, 102, 4419–4424. [Google Scholar] [CrossRef] [PubMed]
- Cambray, G.; Sanchez-Alberola, N.; Campoy, S.; Guerin, É.; Da Re, S.; González-Zorn, B.; Ploy, M.C.; Barbé, J.; Mazel, D.; Erill, I. Prevalence of SOS-mediated control of integron integrase expression as an adaptive trait of chromosomal and mobile integrons. Mob. DNA 2011, 2, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osborn, A.M.; Boltner, D. When phage, plasmids, and transposons collide: Genomic islands, and conjugative- and mobilizable-transposons as a mosaic continuum. Plasmid 2002, 48, 202–212. [Google Scholar] [CrossRef]
- Rose, A. TnAbaR1: A novel Tn7-related transposon in Acinetobacter baumannii that contributes to the accumulation and dissemination of large repertoires of resistance genes. Biosci. Horiz. 2010, 3, 40–48. [Google Scholar] [CrossRef]
- Doublet, B.; Boyd, D.; Mulvey, M.R.; Cloeckaert, A. The Salmonella genomic island 1 is an integrative mobilizable element. Mol. Microbiol. 2005, 55, 1911–1924. [Google Scholar] [CrossRef] [PubMed]
- Rameckers, J.; Hummel, S.; Herrmann, B. How many cycles does a PCR need? Determinations of cycle numbers depending on the number of targets and the reaction efficiency factor. Naturwissenschaften 1997, 84, 259–262. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, N.M.; Hackett, K.T.; Dillard, J.P. XerCD-mediated site-specific recombination leads to loss of the 57-kilobase gonococcal genetic island. J. Bacteriol. 2011, 193, 377–388. [Google Scholar] [CrossRef] [PubMed]
- Aranda, J.; Poza, M.; Shingu-Vazquez, M.; Cortes, P.; Boyce, J.D.; Adler, B.; Barbe, J.; Bou, G. Identification of a DNA-damage-inducible regulon in Acinetobacter baumannii. J. Bacteriol. 2013, 195, 5577–5582. [Google Scholar] [CrossRef] [PubMed]
- Starikova, I.; Harms, K.; Haugen, P.; Lunde, T.T.; Primicerio, R.; Samuelsen, O.; Nielsen, K.M.; Johnsen, P.J. A trade-off between the fitness cost of functional integrases and long-term stability of integrons. PLoS Pathog. 2012, 8, e1003043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domingues, S.; da Silva, G.J.; Nielsen, K.M. Integrons: Vehicles and pathways for horizontal dissemination in bacteria. Mob. Genet. Elements 2012, 2, 211–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaneechoutte, M.; Young, D.M.; Ornston, L.N.; De Baere, T.; Nemec, A.; Van Der Reijden, T.; Carr, E.; Tjernberg, I.; Dijkshoorn, L. Naturally transformable Acinetobacter sp. strain ADP1 belongs to the newly described species Acinetobacter baylyi. Appl. Environ. Microbiol. 2006, 72, 932–936. [Google Scholar] [CrossRef] [PubMed]
- Elbourne, L.D.; Tetu, S.G.; Hassan, K.A.; Paulsen, I.T. TransportDB 2.0: A database for exploring membrane transporters in sequenced genomes from all domains of life. Nucleic Acids Res. 2017, 45, D320–D324. [Google Scholar] [CrossRef] [PubMed]
- Legatzki, A.; Grass, G.; Anton, A.; Rensing, C.; Nies, D.H. Interplay of the Czc system and two P-type ATPases in conferring metal resistance to Ralstonia metallidurans. J. Bacteriol. 2003, 185, 4354–4361. [Google Scholar] [CrossRef] [PubMed]
- Nies, D.H. Efflux-mediated heavy metal resistance in prokaryotes. FEMS Microbiol. Rev. 2003, 27, 313–339. [Google Scholar] [CrossRef] [Green Version]
- Purves, J.; Thomas, J.; Riboldi, G.P.; Zapotoczna, M.; Tarrant, E.; Andrew, P.W.; London, A.; Planet, P.J.; Geoghegan, J.A.; Waldron, K.J.; et al. A horizontally gene transferred copper resistance locus confers hyper-resistance to antibacterial copper toxicity and enables survival of community acquired methicillin resistant Staphylococcus aureus USA300 in macrophages. Environ. Microbiol. 2018, 40, 1576–1589. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | ST | Relevant Characteristics | Reference |
|---|---|---|---|
| A424 | 1 | Clinical isolate from Croatia | [11] |
| A424 ΔG08 | 1 | G08::aacC1 | This study |
| A424 pWSK129-WHG08 | 1 | Complemented ΔG08 strain | This study |
| AYE | 1 | Epidemic MDR type strain, France | [10] |
| AB0057 | 1 | MDR type strain | [23] |
| ATCC 17978 | 437 | Reference strain | [17] |
| ATCC 17978 ΔG62 | 437 | G62::aacC1 | This study |
| ATCC 17978 pWSK129-WHG62 | 437 | Complemented ΔG62 strain | This study |
| KR3831 | 1 | Clinical isolate from SQUH, Oman | This study |
| ATCC 17978 | ATCC 17978 ΔG62 | A424 | A424 ΔG08 | |
|---|---|---|---|---|
| ZnSO4 | 4 * | 2 | 0.5 | 0.5 |
| CuSO4 | 8 | 8 | 8 | 8 |
| CdCl2 | 4 | 2 | 2 | 2 |
| MnSO4 | 10 | 10 | 1 | 0.5 |
| FeSO4 | 4 | 4 | 4 | 4 |
| CoBr2 | 4 | 2 | 4 | 4 |
| NiSO4 | 10 | 5 | 10 | 10 |
| As2O3 | 4 | 4 | 4 | 4 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
AL-Jabri, Z.; Zamudio, R.; Horvath-Papp, E.; Ralph, J.D.; AL-Muharrami, Z.; Rajakumar, K.; Oggioni, M.R. Integrase-Controlled Excision of Metal-Resistance Genomic Islands in Acinetobacter baumannii. Genes 2018, 9, 366. https://doi.org/10.3390/genes9070366
AL-Jabri Z, Zamudio R, Horvath-Papp E, Ralph JD, AL-Muharrami Z, Rajakumar K, Oggioni MR. Integrase-Controlled Excision of Metal-Resistance Genomic Islands in Acinetobacter baumannii. Genes. 2018; 9(7):366. https://doi.org/10.3390/genes9070366
Chicago/Turabian StyleAL-Jabri, Zaaima, Roxana Zamudio, Eva Horvath-Papp, Joseph D. Ralph, Zakariya AL-Muharrami, Kumar Rajakumar, and Marco R. Oggioni. 2018. "Integrase-Controlled Excision of Metal-Resistance Genomic Islands in Acinetobacter baumannii" Genes 9, no. 7: 366. https://doi.org/10.3390/genes9070366
APA StyleAL-Jabri, Z., Zamudio, R., Horvath-Papp, E., Ralph, J. D., AL-Muharrami, Z., Rajakumar, K., & Oggioni, M. R. (2018). Integrase-Controlled Excision of Metal-Resistance Genomic Islands in Acinetobacter baumannii. Genes, 9(7), 366. https://doi.org/10.3390/genes9070366