Genomic Diversity of Listeria monocytogenes Isolated from Clinical and Non-Clinical Samples in Chile



 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Listeria monocytogenes Isolates Used in This Study
2.2. Genome Sequencing and Annotation
2.3. Lineage Determination and Phylogenetic Analysis
2.4. Subtyping
2.5. Screening of Virulence Genes and Stress-Related Elements
3. Results and Discussion
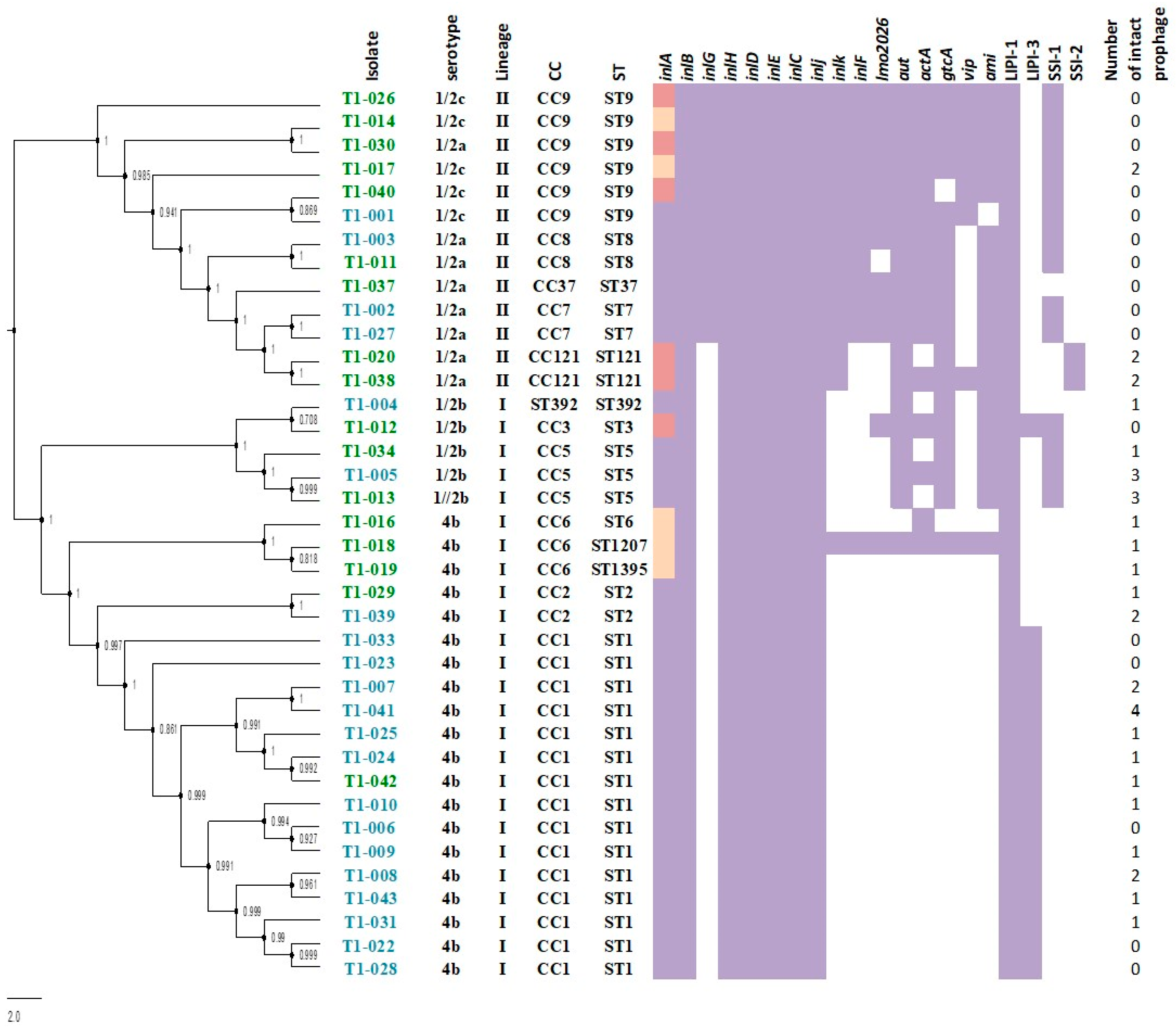
3.1. Listeria monocytogenes Isolates Are Mostly Represented by Clonal Complexes Distributed Worldwide and Involved in Human Infections and Outbreaks
3.2. Isolates of the Pulsed Field Gel Electrophoresis Type Causing the 2008–2009 Outbreaks Showed Genetic Relatedness to Other Worldwide Clinical Isolates
3.3. Clinical and Non-Clinical L. monocytogenes Isolates Showed Distinct Virulence and Stress Survival Genetic Elements
3.4. Presence of One Novel PMSC Mutation in the inlA Gene Along with Additional PMSC Reported in Other Countries in Isolates from Non-Clinical Samples
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Swaminathan, B.; Gerner-Smidt, P. The epidemiology of human listeriosis. Microbes Infect. 2007, 9, 1236–1243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mead, P.S.; Slutsker, L.; Dietz, V.; McCaig, L.F.; Bresee, J.S.; Shapiro, C.; Griffin, P.M.; Tauxe, R.V. Food-related illness and death in the United States. Emerg. Infect. Dis. 1999, 5, 607–625. [Google Scholar] [CrossRef] [PubMed]
- Farber, J.M.; Peterkin, P.I. Listeria monocytogenes, a food-borne pathogen. Microbiol. Rev. 1991, 55, 476–511. [Google Scholar] [CrossRef] [PubMed]
- McLauchlin, J.; Mitchell, R.T.; Smerdon, W.J.; Jewell, K. Listeria monocytogenes and listeriosis: A review of hazard characterisation for use in microbiological risk assessment of foods. Int. J. Food Microbiol. 2004, 92, 15–33. [Google Scholar] [CrossRef]
- Ragon, M.; Wirth, T.; Hollandt, F.; Lavenir, R.; Lecuit, M.; Le Monnier, A.; Brisse, S. A new perspective on Listeria monocytogenes evolution. PLoS Pathog. 2008, 4. [Google Scholar] [CrossRef] [PubMed]
- Chenal-Francisque, V.; Lopez, J.; Cantinelli, T.; Caro, V.; Tran, C.; Leclercq, A.; Lecuit, M.; Brisse, S. Worldwide distribution of major clones of Listeria monocytogenes. Emerg. Infect. Dis. 2011, 17, 1110–1112. [Google Scholar] [CrossRef] [PubMed]
- Cantinelli, T.; Chenal-Francisque, V.; Diancourt, L.; Frezal, L.; Leclercq, A.; Wirth, T.; Lecuit, M.; Brisse, S. Epidemic clones of Listeria monocytogenes are widespread and ancient clonal groups. J. Clin. Microbiol. 2013, 51, 3770–3779. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Boland, J.A.; Kuhn, M.; Berche, P.; Chakraborty, T.; Domi, G.; González-Zorn, B.; Wehland, J. Listeria pathogenesis and molecular virulence determinants Listeria pathogenesis and molecular virulence determinants. Clin. Microbiol. Rev. 2001, 14, 584–640. [Google Scholar] [CrossRef] [PubMed]
- Vazquez-Boland, J.A.; Dominguez-Bernal, G.; Gonzalez-Zorn, B.; Kreft, J.; Goebel, W. Pathogenicity islands and virulence evolution in Listeria. Microbes Infect. 2001, 3, 571–584. [Google Scholar] [CrossRef]
- Gaillard, J.L.; Jaubert, F.; Berche, P. The inlAB locus mediates the entry of Listeria monocytogenes into hepatocytes in vivo. J. Exp. Med. 1996, 183, 359–369. [Google Scholar] [CrossRef] [PubMed]
- Raffelsbauer, D.; Bubert, A.; Engelbrecht, F.; Scheinpflug, J.; Simm, A.; Hess, J.; Kaufmann, S.H.E.; Goebel, W. The gene cluster inlC2DE of Listeria monocytogenes contains additional new internalin genes and is important for virulence in mice. Mol. Genet. Genom. 1998, 260, 144–158. [Google Scholar] [CrossRef]
- Sabet, C.; Lecuit, M.; Cabanes, D.; Cossart, P. LPXTG protein InlJ, a newly identified internalin involved in Listeria monocytogenes virulence. Infect. Immun. 2005, 73, 6912–6922. [Google Scholar] [CrossRef] [PubMed]
- Neves, D.; Job, V.; Dortet, L.; Cossart, P.; Dessen, A. Structure of internalin InlK from the human pathogen Listeria monocytogenes. J. Mol. Biol. 2013, 425, 4520–4529. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Strain, E.A.; Allard, M.; Brown, E.W. Genome sequence of L. monocytogenes Strains J1816 and J1-220, associated with human outbreaks. J. Bacteriol. 2011, 193, 3424–3425. [Google Scholar] [CrossRef] [PubMed]
- Roche, S.M.; Grépinet, O.; Kerouanton, A.; Ragon, M.; Leclercq, A.; Témoin, S.; Schaeffer, B.; Skorski, G.; Mereghetti, L.; Le Monnier, A.; et al. Polyphasic characterization and genetic relatedness of low-virulence and virulent Listeria monocytogenes isolates. BMC Microbiol. 2012, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maury, M.; Tsai, Y.H.; Charlier, C.; Touchon, M.; Chenal-Francisque, V.; Leclercq, A.; Criscuolo, A.; Gaultier, C.; Roussel, S.; Brisabois, A.; et al. Uncovering Listeria monocytogenes hypervirulence by harnessing its biodiversity. Nat. Genet. 2016, 48, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Nightingale, K.K.; Windham, K.; Martin, K.E.; Yeung, M.; Wiedmann, M. Select Listeria monocytogenes subtypes commonly found in foods carry disctinct nonsense mutations in inlA. Appl. Environ. Microbiol. 2005, 71, 8764–8772. [Google Scholar] [CrossRef] [PubMed]
- Harter, E.; Wagner, E.M.; Zaiser, A.; Halecker, S.; Wagner, M.; Rychli, K. The novel stress survival islet 2 (SSI-2), predominantly present in Listeria monocytogenes strains of ST121, is involved in alkaline and oxidative stress response. Appl. Environ. Microbiol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Ryan, S.; Begley, M.; Hill, C.; Gahan, C.G.M. A five-gene stress survival islet (SSI-1) that contributes to the growth of Listeria monocytogenes in suboptimal conditions. J. Appl. Microbiol. 2010, 109, 984–995. [Google Scholar] [CrossRef] [PubMed]
- MINSAL, Departamento de Epidemiología. Informe Listeriosis Actualizado el 15 de Septiembre 2011. Available online: http://www.ispch.cl/sites/default/files/documento/2011/09/listeria2011.pdf (accessed on 31 July 2018).
- MINSAL, Departamento de Epidemiología; Informe año 2017. Situación Epidemiológica de Listeriosis en Chile. 2017, pp. 1–11. Available online: http://epi.minsal.cl/wp-content/uploads/2018/04/INFORME-ANUAL-LISTERIOSIS-2017_2018-03-09-RevIRO-SAF.pdf (accessed on 31 July 2018).
- Saludes, M.; Troncoso, M.; Figueroa, G. Presence of Listeria monocytogenes in Chilean food matrices. Food Control 2015, 50, 331–335. [Google Scholar] [CrossRef]
- Cordano, A.M.; Jacquet, C. Listeria monocytogenes isolated from vegetable salads sold at supermarkets in Santiago, Chile: Prevalence and strain characterization. Int. J. Food Microbiol. 2009, 132, 176–179. [Google Scholar] [CrossRef] [PubMed]
- Montero, D.; Bodero, M.; Riveros, G.; Lapierre, L.; Gaggero, A.; Vidal, R.M.; Vidal, M. Molecular epidemiology and genetic diversity of Listeria monocytogenes isolates from a wide variety of ready-to-eat foods and their relationship to clinical strains from listeriosis outbreaks in Chile. Front. Microbiol. 2015, 6, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 31 July 2018).
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [PubMed]
- Tatusova, T.; Dicuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef] [PubMed]
- Treangen, T.J.; Ondov, B.D.; Koren, S.; Phillippy, A.M. The Harvest suite for rapid core-genome alignment and visualization of thousands of intraspecific microbial genomes. Genome Biol. 2014, 15. [Google Scholar] [CrossRef]
- Kaas, R.S.; Leekitcharoenphon, P.; Aarestrup, F.M.; Lund, O. Solving the problem of comparing whole bacterial genomes across different sequencing platforms. PLoS ONE 2014, 9, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doumith, M.; Buchrieser, C.; Glaser, P.; Jacquet, C.; Martin, P. Differentiation of the major Listeria monocytogenes serovars by multiplex PCR. J. Clin. Microbiol. 2004, 42, 3819–3822. [Google Scholar] [CrossRef] [PubMed]
- Larsen, M.V.; Cosentino, S.; Rasmussen, S.; Friis, C.; Hasman, H.; Marvig, R.L.; Jelsbak, L.; Sicheritz-Pontén, T.; Ussery, D.W.; Aarestrup, F.M.; et al. Multilocus sequence typing of total-genome-sequenced bacteria. J. Clin. Microbiol. 2012, 50, 1355–1361. [Google Scholar] [CrossRef] [PubMed]
- Arndt, D.; Grant, J.R.; Marcu, A.; Sajed, T.; Pon, A.; Liang, Y.; Wishart, D.S. PHASTER: A better, faster version of the PHAST phage search tool. Nucleic Acids Res. 2016, 44, W16–W21. [Google Scholar] [CrossRef] [PubMed]
- Carattoli, A.; Zankari, E.; García-Fernández, A.; Larsen, M.V.; Lund, O.; Villa, L.; Aarestrup, F.M.; Hasman, H. In silico detection and typing of plasmids using plasmidfinder and plasmid multilocus sequence typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [PubMed]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef] [PubMed]
- Madden, T. The BLAST Sequence Analysis Tool. In The NCBI Handbook National Library of Medicine (US); McEntyre, J., Ed.; National Center for Biotechnology Information: Bethesda, MD, USA, 2013. [Google Scholar]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7. [Google Scholar] [CrossRef] [PubMed]
- Joensen, K.G.; Scheutz, F.; Lund, O.; Hasman, H.; Kaas, R.S.; Nielsen, E.M.; Aarestrup, F.M. Real-time whole-genome sequencing for routine typing, surveillance, and outbreak detection of verotoxigenic Escherichia coli. J. Clin. Microbiol. 2014, 52, 1501–1510. [Google Scholar] [CrossRef] [PubMed]
- Cartwright, E.J.; Jackson, K.A.; Johnson, S.D.; Graves, L.M.; Silk, B.J.; Mahon, B.E. Listeriosis outbreaks and associated food vehicles, United States, 1998–2008. Emerg. Infect. Dis. 2013, 19, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Orsi, R.H.; den Bakker, H.C.; Wiedmann, M. Listeria monocytogenes lineages: Genomics, evolution, ecology, and phenotypic characteristics. Int. J. Med. Microbiol. 2011, 301, 79–96. [Google Scholar] [CrossRef] [PubMed]
- Graves, L.M.; Swaminathan, B. PulseNet standardized protocol for subtyping Listeria monocytogenes by macrorestriction and pulsed-field gel electrophoresis. Int. J. Food Microbiol. 2001, 65, 55–62. [Google Scholar] [CrossRef]
- Chen, J.; Chen, Q.; Jiang, J.; Hu, H.; Ye, J.; Fang, W. Serovar 4b complex predominates among Listeria monocytogenes isolates from imported aquatic products in China. Foodborne Pathog. Dis. 2010, 7, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Braga, V.; Vázquez, S.; Vico, V.; Pastorino, V.; Mota, M.I.; Legnani, M.; Schelotto, F.; Lancibidad, G.; Varela, G. Prevalence and serotype distribution of Listeria monocytogenes isolated from foods in Montevideo-Uruguay. Braz. J. Microbiol. 2017, 48, 689–694. [Google Scholar] [CrossRef] [PubMed]
- Vallim, D.C.; Barroso Hofer, C.; Lisbôa, R.D.C.; Victor Barbosa, A.; Alves Rusak, L.; Dos Reis, C.M.F.; Hofer, E. Twenty years of Listeria in Brazil: Occurrence of Listeria species and Listeria monocytogenes serovars in food samples in Brazil between 1990 and 2012. Biomed. Res. Int. 2015. [Google Scholar] [CrossRef] [PubMed]
- Leclercq, A.; Chenal-Francisque, V.; Dieye, H.; Cantinelli, T.; Drali, R.; Brisse, S.; Lecuit, M. Characterization of the novel Listeria monocytogenes PCR serogrouping profile IVb-v1. Int. J. Food Microbiol. 2011, 147, 74–77. [Google Scholar] [CrossRef] [PubMed]
- Burall, L.S.; Grim, C.J.; Datta, A.R. A clade of Listeria monocytogenes serotype 4b variant strains linked to recent listeriosis outbreaks associated with produce from a defined geographic region in the US. PLoS ONE 2017, 12. [Google Scholar] [CrossRef] [PubMed]
- Burall, L.S.; Grim, C.J.; Mammel, M.K.; Datta, A.R. A Comprehensive evaluation of the genetic relatedness of Listeria monocytogenes serotype 4b variant strains. Front. Public Health 2017, 5, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kwong, J.C.; Mercoulia, K.; Tomita, T.; Easton, M.; Li, H.Y.; Bulach, D.M.; Stinear, T.P.; Seemann, T.; Howden, B.P. Prospective whole-genome sequencing enhances national surveillance of Listeria monocytogenes. J. Clin. Microbiol. 2016, 54, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Cao, G.; Xu, X.; Allard, M.; Li, P.; Brown, E.; Yang, X.; Pan, H.; Meng, J. Evolution and diversity of Listeria monocytogenes from clinical and food samples in Shanghai, China. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Nightingale, K.K.; Boor, K.J.; Ho, A.; Wiedmann, M.; McGann, P. Distribution of internalin gene profiles of Listeria monocytogenes isolates from different sources associated with phylogenetic lineages. Foodborne Pathog. Dis. 2007, 4, 222–232. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Zhang, J.; Shi, W.; Yang, X.; Li, Y.; Pan, H.; Kuang, D.; Xu, X.; Shi, X.; Meng, J. Molecular characterization and antimicrobial susceptibility of Listeria monocytogenes isolated from foods and humans. Food Control 2016, 70, 96–102. [Google Scholar] [CrossRef]
- Clayton, E.M.; Hill, C.; Cotter, P.D.; Ross, R.P. Real-time PCR assay to differentiate listeriolysin S-positive and -negative strains of Listeria monocytogenes. Appl. Environ. Microbiol. 2011, 77, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Cotter, P.D.; Draper, L.A.; Lawton, E.M.; Daly, K.M.; Groeger, D.S.; Casey, P.G.; Ross, R.P.; Hill, C. Listeriolysin S, a novel peptide haemolysin associated with a subset of lineage I Listeria monocytogenes. PLoS Pathog. 2008, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, J.; Rump, L.; Zhang, Y.; Chen, Y.; Wang, X.; Meng, J. Molecular subtyping and virulence gene analysis of Listeria monocytogenes isolates from food. Food Microbiol. 2013, 35, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Clayton, E.M.; Daly, K.M.; Guinane, C.M.; Hill, C.; Cotter, P.D.; Ross, P.R. Atypical Listeria innocua strains possess an intact LIPI-3. BMC Microbiol. 2014, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanki, M.; Naruse, H.; Taguchi, M.; Kumeda, Y. Characterization of specific alleles in inlA and prfA of Listeria monocytogenes isolated from foods in Osaka, Japan and their ability to invade Caco-2 cells. Int. J. Food Microbiol. 2015, 211, 18–22. [Google Scholar] [CrossRef] [PubMed]
- Kovacevic, J.; Arguedas-Villa, C.; Wozniak, A.; Tasara, T.; Allen, K.J. Examination of food chain-derived Listeria monocytogenes strains of different serotypes reveals considerable diversity in inlA genotypes, mutability, and adaptation to cold temperatures. Appl. Environ. Microbiol. 2013, 79, 1915–1922. [Google Scholar] [CrossRef] [PubMed]
- Gorski, L.; Parker, C.T.; Liang, A.S.; Walker, S.; Romanolo, K.F. The majority of genotypes of the virulence gene inlA are intact among natural watershed isolates of Listeria monocytogenes from the central California Coast. PLoS ONE 2016, 11, e0167566. [Google Scholar] [CrossRef] [PubMed]
- Hingston, P.; Chen, J.; Dhillon, B.K.; Laing, C.; Bertelli, C.; Gannon, V.; Tasara, T.; Allen, K.; Brinkman, F.S.L.; Hansen, L.T.; et al. Genotypes associated with Listeria monocytogenes isolates displaying impaired or enhanced tolerances to cold, salt, acid, or desiccation stress. Front. Microbiol. 2017, 8, 369. [Google Scholar] [CrossRef] [PubMed]
- Olier, M.; Pierre, F.; Lemaître, J.P.; Divies, C.; Rousset, A.; Guzzo, J. Assessment of the pathogenic potential of two Listeria monocytogenes human faecal carriage isolates. Microbiology 2002, 148, 1855–1862. [Google Scholar] [CrossRef] [PubMed]
- Rousseaux, S.; Olier, M.; Piveteau, P.; Guzzo, J. Use of PCR-restriction fragment length polymorphism of inlA for rapid screening of Listeria monocytogenes strains deficient in the ability to invade Caco-2 cells. Appl. Environ. Microbiol. 2004, 70, 2180–2185. [Google Scholar] [CrossRef] [PubMed]
- Handa-Miya, S.; Kimura, B.; Takahashi, H.; Sato, M.; Ishikawa, T.; Igarashi, K.; Fujii, T. Nonsense-mutated inlA and prfA not widely distributed in Listeria monocytogenes isolates from ready-to-eat seafood products in Japan. Int. J. Food Microbiol. 2007, 117, 312–318. [Google Scholar] [CrossRef] [PubMed]
- Gelbíčová, T.; Koláčková, I.; Pantůček, R.; Karpíšková, R. A novel mutation leading to a premature stop codon in inlA of Listeria monocytogenes isolated from neonatal listeriosis. New Microbiol. 2015, 38, 293–296. [Google Scholar] [PubMed]
- Dramsi, S.; Trieu-Cuot, P.; Bierne, H. Sorting sortases: A nomenclature proposal for the various sortases of Gram-positive bacteria. Res. Microbiol. 2005, 156, 289–297. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Isolates | Origin 1 | Source | Isolation Date | Geographic Location 2 | Pulse Type 3 | Serogroup 4 | Sequence Type 4 | Clonal Complex 4 |
|---|---|---|---|---|---|---|---|---|
| T1-001 | Clinical | Amniotic fluid | 2009 | Santiago | 167 | IIc | ST9 | CC9 |
| T1-002 | Clinical | Blood | 2010 | Santiago | 3 | IIa | ST7 | CC7 |
| T1-003 | Clinical | Cerebrospinal fluid | 2011 | Los Lagos | 114 | IIa | ST8 | CC8 |
| T1-004 | Clinical | Cerebrospinal fluid | 2011 | Santiago | 260 | IIb | ST392 | - |
| T1-005 | Clinical | Cerebrospinal fluid | 2010 | Valparaiso | 19 | IIb | ST5 | CC5 |
| T1-006 | Clinical | Blood | 2010 | Aysén | 48 | IVb | ST1 | CC1 |
| T1-007 | Clinical | Blood | 2010 | Santiago | 197 | IVb | ST1 | CC1 |
| T1-008 | Clinical | Cerebrospinal fluid | 2011 | Araucanía | 235 | IVb | ST1 | CC1 |
| T1-009 | Clinical | Amniotic fluid | 2011 | Santiago | 252 | IVb | ST1 | CC1 |
| T1-010 | Clinical | Blood | 2011 | O’Higgins | 264 | IVb | ST1 | CC1 |
| T1-011 | Food | Ham | 2010 | Santiago | 167 | IIa | ST8 | CC8 |
| T1-012 | Food | Sausage | 2010 | Los Lagos | 147 | IIb | ST3 | CC3 |
| T1-013 | Food | Ice cream | 2010 | Santiago | 212 | IIb | ST5 | CC5 |
| T1-014 | Food | Sausage | 2010 | Los Lagos | 210 | IIc | ST9 | CC9 |
| T1-016 | Food | Ham | 2010 | Santiago | 211 | IVb | ST1207 | CC6 |
| T1-017 | Food | Ham | 2010 | Santiago | 99 | IIc | ST9 | CC9 |
| T1-018 | Food | Ice cream | 2010 | Santiago | 156 | IVb | ST1395 7 | CC6 |
| T1-019 | Environment | Food plant | N/A | Bío-Bío | N/A | IVb | ST6 | CC6 |
| T1-020 | Environment | Food plant | N/A | Bío-Bío | N/A | IIa | ST121 | CC121 |
| T1-022 | Clinical | Blood | 2011 | Valparaiso | 256 | IVb | ST1 | CC1 |
| T1-023 | Clinical | Blood | 2008 | Santiago | 9 5 | IVb | ST1 | CC1 |
| T1-024 | Clinical | Blood | 2008 | O’Higgins | 9 5 | IVb | ST1 | CC1 |
| T1-025 | Clinical | Blood | 2010 | Santiago | 99 | IVb | ST1 | CC1 |
| T1-026 | Food | Sausage | 2010 | Los Lagos | 2 | IIc | ST9 | CC9 |
| T1-027 | Clinical | Blood | 2010 | Santiago | 46 | IIa | ST7 | CC7 |
| T1-028 | Clinical | Blood | 2008 | Bío-Bío | 9 5 | IVb | ST1 | CC1 |
| T1-029 | Food | Pork pate | 2010 | Araucanía | 126 | IVb | ST2 | CC2 |
| T1-030 | Food | Sausage | 2009 | Bío-Bío | 1 6 | IIa | ST9 | CC9 |
| T1-031 | Clinical | Human | 2010 | O’Higgins | 20 | IVb | ST1 | CC1 |
| T1-033 | Clinical | Blood | 2010 | Araucanía | 133 | IVb | ST1 | CC1 |
| T1-034 | Food | Ice cream | 2010 | Santiago | 64 | IIb | ST5 | CC5 |
| T1-037 | Clinical | Peritoneal fluid | 2011 | Bío-Bío | 137 | IIa | ST37 | CC37 |
| T1-038 | Food | Ham | 2010 | Santiago | 53 | IIa | ST121 | CC121 |
| T1-039 | Clinical | Cerebrospinal fluid | 2010 | Santiago | 209 | IVb | ST2 | CC2 |
| T1-040 | Food | Beef | 2009 | Bío-Bío | 1 6 | IIc | ST9 | CC9 |
| T1-041 | Clinical | Blood | 2011 | Bío-Bío | 245 | IVb | ST1 | CC1 |
| T1-042 | Food | Cheese | 2009 | Santiago | 9 5 | IVb | ST1 | CC1 |
| T1-043 | Clinical | Blood | 2010 | Santiago | 58 | IVb | ST1 | CC1 |
| Number of Isolates | InlA Length (aa) | Mutation Type (PMSC) | Nucleotide Mutation Position | Functional | Reference |
|---|---|---|---|---|---|
| 27 | 800 | - | - | yes | Glaser et al., 2001 |
| 5 | 797 | NA | NA | yes | Chen et al., 2011 |
| 1 | 684 | 11 | 2054 (G to A) | no | Rousseaux et al., 2002 |
| 1 | 527 | 13 | 1579 (Ato T) | no | Handa-Miya et al., 2007 |
| 2 | 491 | 6 | 1474 (Cto T) | no | Olier et al., 2002 |
| 1 | 325 | 19 | 976 (G to T) | no | Gelbicova et al., 2015 |
| 1 | 277 | Novel | 821 (deletion A) | no | In this study |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toledo, V.; Den Bakker, H.C.; Hormazábal, J.C.; González-Rocha, G.; Bello-Toledo, H.; Toro, M.; Moreno-Switt, A.I. Genomic Diversity of Listeria monocytogenes Isolated from Clinical and Non-Clinical Samples in Chile. Genes 2018, 9, 396. https://doi.org/10.3390/genes9080396
Toledo V, Den Bakker HC, Hormazábal JC, González-Rocha G, Bello-Toledo H, Toro M, Moreno-Switt AI. Genomic Diversity of Listeria monocytogenes Isolated from Clinical and Non-Clinical Samples in Chile. Genes. 2018; 9(8):396. https://doi.org/10.3390/genes9080396
Chicago/Turabian StyleToledo, Viviana, Henk C. Den Bakker, Juan Carlos Hormazábal, Gerardo González-Rocha, Helia Bello-Toledo, Magaly Toro, and Andrea I. Moreno-Switt. 2018. "Genomic Diversity of Listeria monocytogenes Isolated from Clinical and Non-Clinical Samples in Chile" Genes 9, no. 8: 396. https://doi.org/10.3390/genes9080396
APA StyleToledo, V., Den Bakker, H. C., Hormazábal, J. C., González-Rocha, G., Bello-Toledo, H., Toro, M., & Moreno-Switt, A. I. (2018). Genomic Diversity of Listeria monocytogenes Isolated from Clinical and Non-Clinical Samples in Chile. Genes, 9(8), 396. https://doi.org/10.3390/genes9080396