Golgi Reassembly and Stacking Protein (GRASP) Participates in Vesicle-Mediated RNA Export in Cryptococcus neoformans

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Material and Methods
2.1. Fungal Strains and Growth Conditions
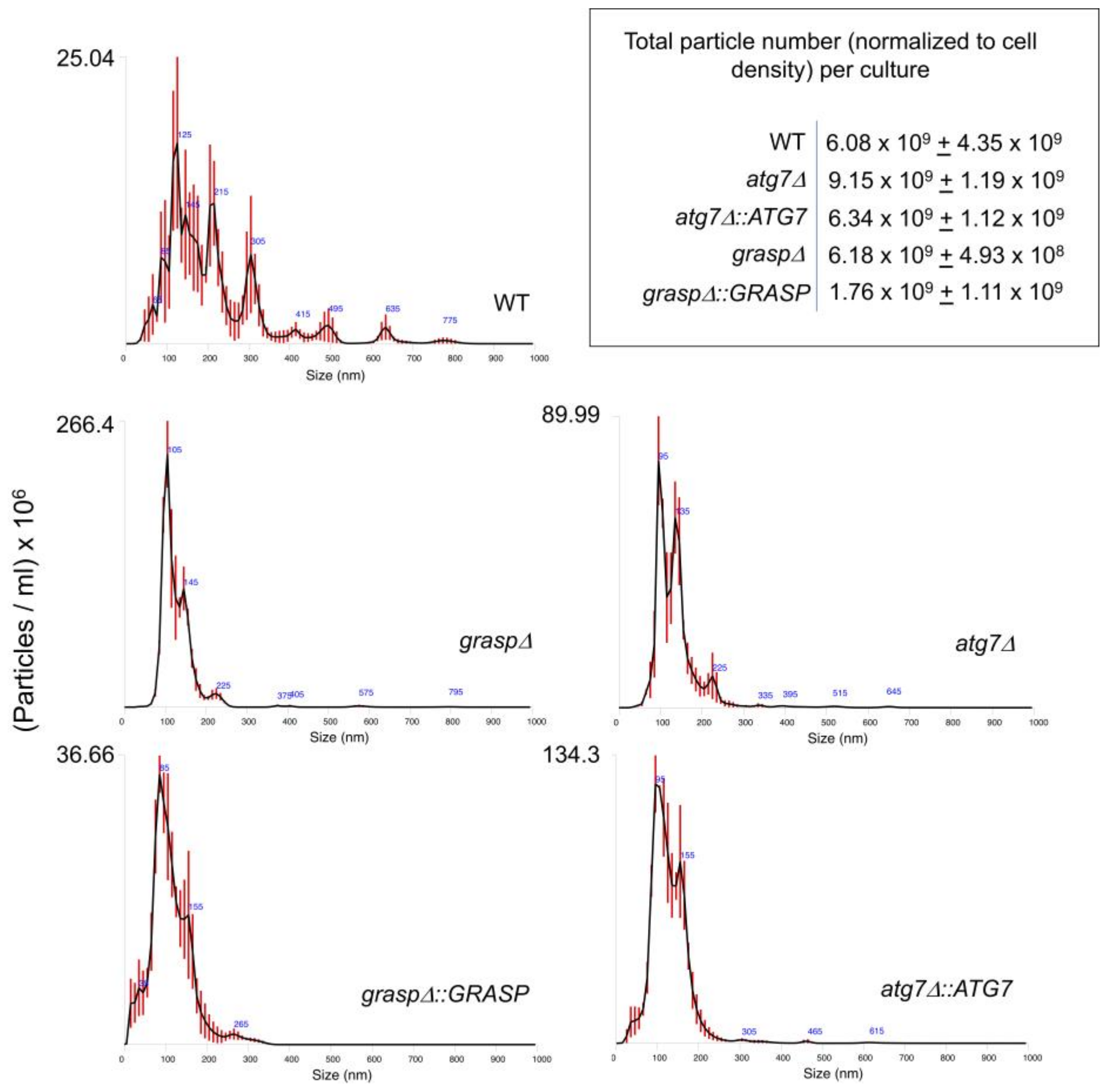
2.2. Extracellular Vesicle Isolation and Diameter Determination
2.3. Small RNA Isolation
2.4. RNA Sequencing
2.5. Cellular RNA Isolation and Quantitative PCR
2.6. In Silico Data Analysis
2.7. Data Access
3. Results
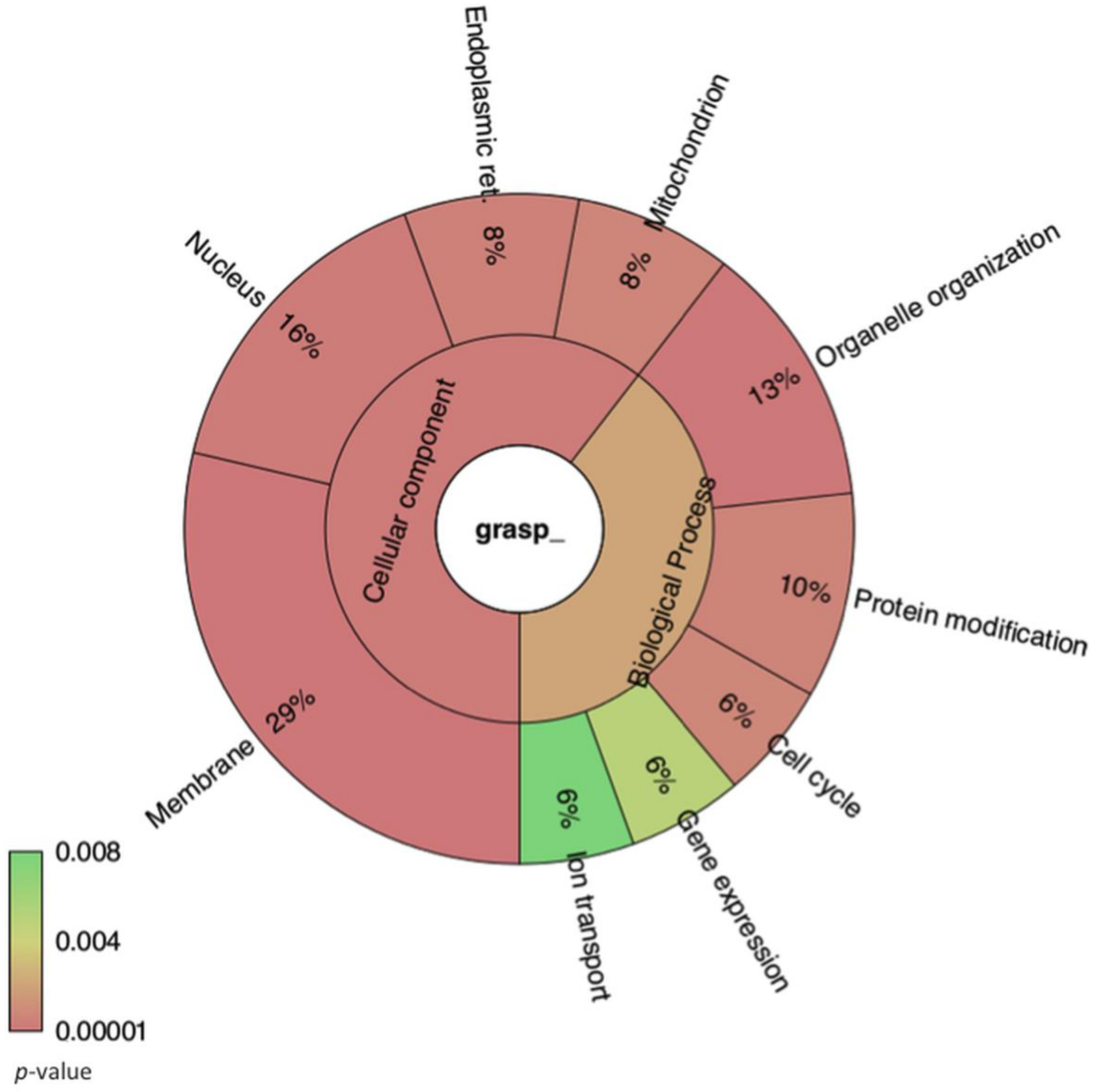
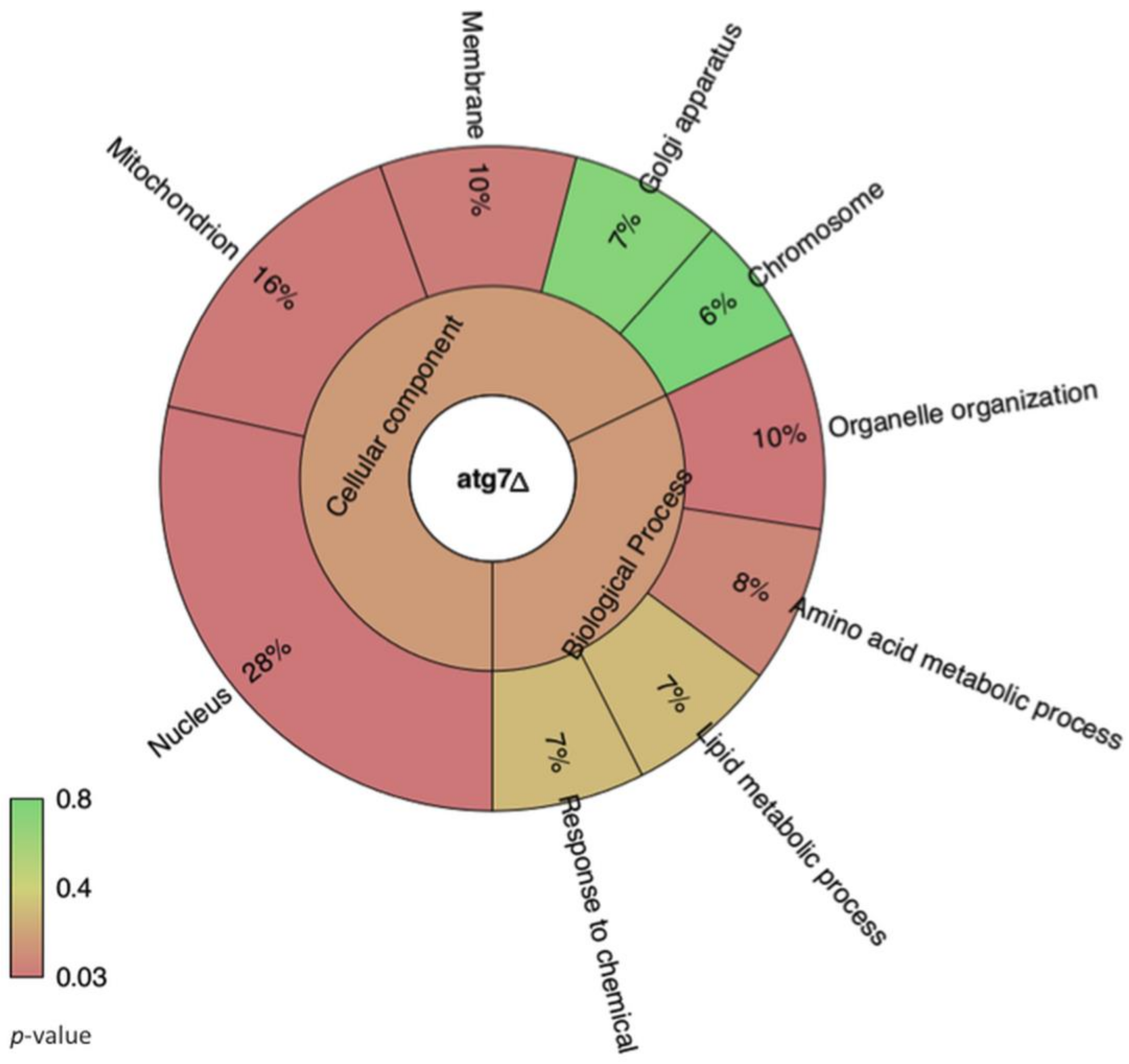
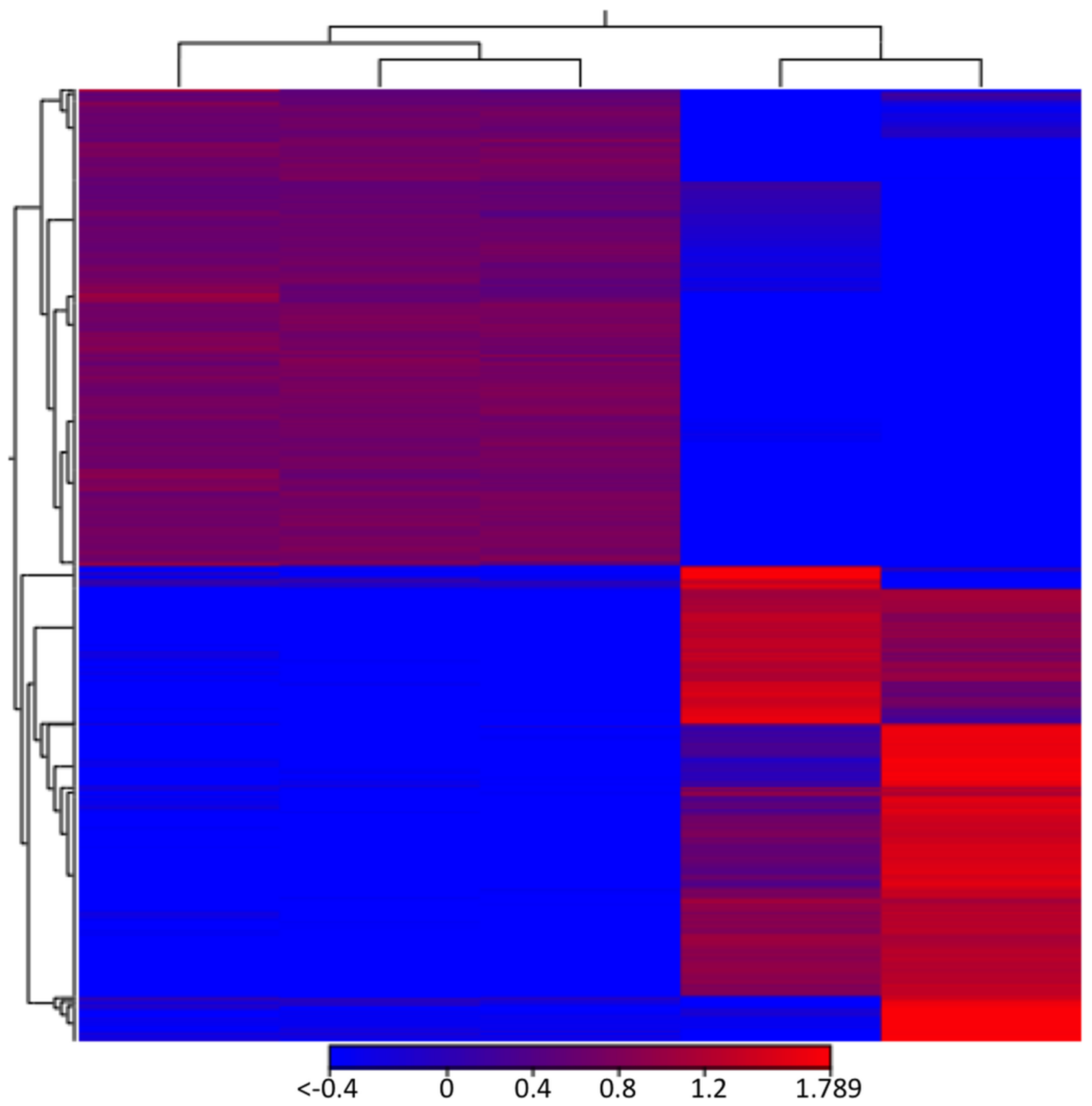
3.1. Lack of GRASP Results in Changes in the RNA Content of Cryptococcus neoformans Extracellular Vesicles
3.2. Comparison of Cellular RNA Versus Extracellular Vesicle RNA Composition
3.3. Intronic Reads
3.4. Non-Coding RNAs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Coakley, G.; Maizels, R.M.; Buck, A.H. Exosomes and other extracellular vesicles: The new communicators in parasite infections. Trends Parasitol. 2015, 31, 477–489. [Google Scholar] [CrossRef] [PubMed]
- Tkach, M.; Théry, C. Communication by extracellular vesicles: Where we are and where we need to go. Cell 2016, 164, 1226–1232. [Google Scholar] [CrossRef] [PubMed]
- Deatheragea, B.L.; Cooksona, B.T. Membrane vesicle release in bacteria, eukaryotes Eukaryotes and Archaea: A conserved yet underappreciated aspect of microbial life. Infect. Immun. 2012, 80, 1948–1957. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.L.; Nimrichter, L.; Oliveira, D.L.; Frases, S.; Miranda, K.; Zaragoza, O.; Alvarez, M.; Nakouzi, A.; Feldmesser, M.; Casadevall, A. Vesicular polysaccharide export in Cryptococcus neoformans is a eukaryotic solution to the problem of fungal trans-cell wall transport. Eukaryot. Cell 2007, 6, 48–59. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, P.C.; Nakayasu, E.S.; Rodrigues, M.L.; Frases, S.; Casadevall, A.; Zancope-Oliveira, R.M.; Almeida, I.C.; Nosanchuk, J.D. Vesicular transport in Histoplasma capsulatum: An effective mechanism for trans-cell wall transfer of proteins and lipids in ascomycetes. Cell Microbiol. 2008, 10, 1695–1710. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.L.; Nakayasu, E.S.; Oliveira, D.L.; Nimrichter, L.; Nosanchuk, J.D.; Almeida, I.C.; Casadevall, A. Extracellular vesicles produced by Cryptococcus neoformans contain protein components associated with virulence. Eukaryot. Cell 2008, 7, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Eisenman, H.C.; Frases, S.; Nicola, A.M.; Rodrigues, M.L.; Casadevall, A. Vesicle-associated melanization in Cryptococcus neoformans. Microbiology 2009, 155, 3860–3867. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, J.; Oliveira, D.L.; Joffe, L.S.; Hu, G.; Gazos-Lopes, F.; Fonseca, F.L.; Almeida, I.C.; Frases, S.; Kronstad, J.W.; Rodrigues, M.L. Role of the Apt1 protein in polysaccharide secretion by Cryptococcus neoformans. Eukaryot. Cell 2014, 13, 715–726. [Google Scholar] [CrossRef] [PubMed]
- Peres da Silva, R.; Puccia, R.; Rodrigues, M.L.; Oliveira, D.L.; Joffe, L.S.; César, G.V.; Nimrichter, L.; Goldenberg, S.; Alves, L.R. Extracellular vesicle-mediated export of fungal RNA. Sci. Rep. 2015, 5, 7763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, D.L.; Nakayasu, E.S.; Joffe, L.S.; Guimarães, A.J.; Sobreira, T.J.; Nosanchuk, J.D.; Cordero, R.J.; Frases, S.; Casadevall, A.; Almeida, I.C.; et al. Biogenesis of extracellular vesicles in yeast: Many questions with few answers. Commun. Integr. Biol. 2010, 3, 533–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, M.L.; Franzen, A.J.; Nimrichter, L.; Miranda, K. Vesicular mechanisms of traffic of fungal molecules to the extracellular space. Curr. Opin. Microbiol. 2013, 16, 414–420. [Google Scholar] [CrossRef] [PubMed]
- Barr, F.A.; Puype, M.; Vandekerckhove, J.; Warren, G. GRASP65, a protein involved in the stacking of Golgi cisternae. Cell 1997, 91, 253–262. [Google Scholar] [CrossRef]
- Shorter, J.; Watson, R.; Giannakou, M.E.; Clarke, M.; Warren, G.; Barr, F.A. GRASP55, a second mammalian GRASP protein involved in the stacking of Golgi cisternae in a cell-free system. EMBO J. 1999, 18, 4949–4960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabouille, C.; Malhotra, V.; Nickel, W. Diversity in unconventional protein secretion. J. Cell Sci. 2012, 125, 5251–5255. [Google Scholar] [CrossRef] [PubMed]
- Gee, H.Y.; Noh, S.H.; Tang, B.L.; Kim, K.H.; Lee, M.G. Rescue of ΔF508-CFTR trafficking via a GRASP-dependent unconventional secretion pathway. Cell 2011, 146, 746–760. [Google Scholar] [CrossRef] [PubMed]
- Grieve, A.G.; Rabouille, C. Extracellular cleavage of E-cadherin promotes epithelial cell extrusion. J. Cell Sci. 2014, 127, 3331–3346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinseth, M.A.; Anjard, C.; Fuller, D.; Guizzunti, G.; Loomis, W.F.; Malhotra, V. The Golgi-associated protein GRASP is required for unconventional protein secretion during development. Cell 2007, 130, 524–534. [Google Scholar] [CrossRef] [PubMed]
- Duran, J.M.; Anjard, C.; Stefan, C.; Loomis, W.F.; Malhotra, V. Unconventional secretion of Acb1 is mediated by autophagosomes. J. Cell Biol. 2010, 188, 527–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manjithaya, R.; Anjard, C.; Loomis, W.F.; Subramani, S. Unconventional secretion of Pichia pastoris Acb1 is dependent on GRASP protein, peroxisomal functions and autophagosome formation. J. Cell Biol. 2010, 188, 537–546. [Google Scholar] [CrossRef] [PubMed]
- Kmetzsch, L.; Joffe, L.S.; Staats, C.C.; de Oliveira, D.L.; Fonseca, F.L.; Cordero, R.J.; Casadevall, A.; Nimrichter, L.; Schrank, A.; Vainstein, M.H.; et al. Role for Golgi reassembly and stacking protein (GRASP) in polysaccharide secretion and fungal virulence. Mol. Microbiol. 2011, 81, 206–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glick, D.; Barth, S.; Macleod, K.F. Autophagy: Cellular and molecular mechanisms. J. Pathol. 2010, 221, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Nakatogawa, H.; Suzuki, K.; Kamada, Y.; Ohsumi, Y. Dynamics and diversity in autophagy mechanisms: Lessons from yeast. Nat. Rev. Mol. Cell Biol. 2009, 10, 458–467. [Google Scholar] [CrossRef] [PubMed]
- Lévêque, M.F.; Berry, L.; Cipriano, M.J.; Nguyen, H.M.; Striepen, B.; Besteiro, S. Autophagy-related protein ATG8 has a noncanonical function for apicoplast inheritance in Toxoplasma gondii. MBio 2015, 6, e01446-15. [Google Scholar] [CrossRef] [PubMed]
- DeSelm, C.J.; Miller, B.C.; Zou, W.; Beatty, W.L.; van Meel, E.; Takahata, Y.; Klumperman, J.; Tooze, S.A.; Teitelbaum, S.L.; Virgin, H.W. Autophagy proteins regulate the secretory component of osteoclastic bone resorption. Dev. Cell 2011, 21, 966–974. [Google Scholar] [CrossRef] [PubMed]
- Dreux, M.; Chisari, F.V. Impact of the autophagy machinery on hepatitis Hepatitis C virus infection. Viruses 2011, 3, 1342–1357. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Goldman, S.; Baerga, R.; Zhao, Y.; Komatsu, M.; Jin, S. Adipose-specific deletion of autophagy-related gene 7 (atg7) in mice reveals a role in adipogenesis. Proc. Natl. Acad. Sci. USA 2009, 106, 19860–19865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, I.H.; Kawai, Y.; Fergusson, M.M.; Rovira, I.I.; Bishop, A.J.; Motoyama, N.; Cao, L.; Finkel, T. Atg7 modulates p53 activity to regulate cell cycle and survival during metabolic stress. Science 2012, 336, 225–228. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, D.L.; Fonseca, F.L.; Zamith-Miranda, D.; Nimrichter, L.; Rodrigues, J.; Pereira, M.D.; Reuwsaat, J.C.; Schrank, A.; Staats, C.; Kmetzsch, L.; et al. The putative autophagy regulator Atg7 affects the physiology and pathogenic mechanisms of Cryptococcus neoformans. Future Microbiol. 2016, 11, 1405–1419. [Google Scholar] [CrossRef] [PubMed]
- Bruns, C.; McCaffery, J.M.; Curwin, A.J.; Duran, J.M.; Malhotra, V. Biogenesis of a novel compartment for autophagosome-mediated unconventional protein secretion. J. Cell Biol. 2011, 195, 979–992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maas, S.L.N.; De Vrij, J.; Van Der Vlist, E.J.; Geragousian, B.; Van Bloois, L.; Mastrobattista, E.; Schiffelers, R.M.; Wauben, M.H.M.; Broekman, M.L.D.; Nolte-’t Hoen, E.N. Possibilities and limitations of current technologies for quantification of biological extracellular vesicles and synthetic mimics. J. Control. Release 2015, 200, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.L.; Oliveira, D.L.; Vargas, G.; Girard-Dias, W.; Franzen, A.J.; Frasés, S.; Miranda, K.; Nimrichter, L. Analysis of yeast extracellular vesicles. Methods Mol. Biol. 2016, 1459, 175–190. [Google Scholar] [CrossRef] [PubMed]
- Baggerly, K.A.; Deng, L.; Morris, J.S.; Aldaz, C.M. Differential expression in SAGE: Accounting for normal between-library variation. Bioinformatics 2003, 19, 1477–1483. [Google Scholar] [CrossRef] [PubMed]
- Semenza, J.C.; Hardwick, K.G.; Dean, N.; Pelham, H.R. ERD2, a yeast gene required for the receptor-mediated retrieval of luminal ER proteins from the secretory pathway. Cell 1990, 61, 1349–1357. [Google Scholar] [CrossRef]
- Von Mollard, G.; Stevens, T.H. The Saccharomyces cerevisiae v-SNARE Vti1p is required for multiple membrane transport pathways to the vacuole. Mol. Biol. Cell 1999, 10, 1719–1732. [Google Scholar] [CrossRef] [PubMed]
- Abels, E.R.; Breakefield, X.O. Introduction to extracellular vesicles: Biogenesis, RNA cargo selection, content, release and uptake. Cell. Mol. Neurobiol. 2016, 36, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Lev, S.; Saiardi, A.; Desmarini, D.; Sorrell, T.C.; Djordjevic, J.T. Identification of a major IP5 kinase in Cryptococcus neoformans confirms that PP-IP5/IP7, not IP6, is essential for virulence. Sci. Rep. 2016, 6, 23927. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Hilarion, S.; Paulet, D.; Lee, K.T.; Hon, C.C.; Lechat, P.; Mogensen, E.; Moyrand, F.; Proux, C.; Barboux, R.; Bussotti, G.; et al. Intron retention-dependent gene regulation in Cryptococcus neoformans. Sci. Rep. 2016, 6, 32252. [Google Scholar] [CrossRef] [PubMed]
- Keam, S.P.; Hutvagner, G. tRNA-Derived Fragments (tRFs): Emerging new roles for an ancient RNA in the regulation of gene expression. Life 2015, 5, 1638–1651. [Google Scholar] [CrossRef] [PubMed]
- Maute, R.L.; Schneider, C.; Sumazin, P.; Holmes, A.; Califano, A.; Basso, K.; Dalla-Favera, R. tRNA-derived microRNA modulates proliferation and the DNA damage response and is down-regulated in B cell lymphoma. Proc. Natl. Acad. Sci. USA 2013, 110, 1404–1409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gebetsberger, J.; Zywicki, M.; Künzi, A.; Polacek, N. tRNA-derived fragments target the ribosome and function as regulatory non-coding RNA in Haloferax volcanii. Archaea 2012, 2012, 260909. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Shibata, Y.; Malhotra, A.; Dutta, A. A novel class of small RNAs: tRNA-derived RNA fragments (tRFs). Genes Dev. 2009, 23, 2639–2649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haussecker, D.; Huang, Y.; Lau, A.; Parameswaran, P.; Fire, A.Z.; Kay, M.A. Human tRNA-derived small RNAs in the global regulation of RNA silencing. RNA 2010, 16, 673–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Couvillion, M.T.; Bounova, G.; Purdom, E.; Speed, T.P.; Collins, K. A Tetrahymena Piwi bound to mature tRNA 3′ fragments activates the exonuclease Xrn2 for RNA processing in the nucleus. Mol. Cell 2012, 48, 509–520. [Google Scholar] [CrossRef] [PubMed]
- Goodarzi, H.; Liu, X.; Nguyen, H.C.; Zhang, S.; Fish, L.; Tavazoie, S.F. Endogenous tRNA-derived fragments suppress breast cancer progression via YBX1 displacement. Cell 2015, 161, 790–802. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Silva, M.R.; Cabrera-Cabrera, F.; das Neves, R.F.; Souto-Padrón, T.; de Souza, W.; Cayota, A. Gene expression changes induced by Trypanosoma cruzi shed microvesicles in mammalian host cells: Relevance of tRNA-derived halves. BioMed Res. Int. 2014, 2014, 305239. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Silva, M.R.; das Neves, R.F.; Cabrera-Cabrera, F.; Sanguinetti, J.; Medeiros, L.C.; Robello, C.; Naya, H.; Fernandez-Calero, T.; Souto-Padron, T.; de Souza, W.; et al. Extracellular vesicles shed by Trypanosoma cruzi are linked to small RNA pathways, life cycle regulation and susceptibility to infection of mammalian cells. Parasitol. Res. 2014, 113, 285–304. [Google Scholar] [CrossRef] [PubMed]
- Nolte-’t Hoen, E.N.; Buermans, H.P.; Waasdorp, M.; Stoorvogel, W.; Wauben, M.H.; ’t Hoen, P.A. Deep sequencing of RNA from immune cell-derived vesicles uncovers the selective incorporation of small non-coding RNA biotypes with potential regulatory functions. Nucleic Acids Res. 2012, 40, 9272–9285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vojtech, L.; Woo, S.; Hughes, S.; Levy, C.; Ballweber, L.; Sauteraud, R.P.; Strobl, J.; Westerberg, K.; Gottardo, R.; Tewari, M.; et al. Exosomes in human semen carry a distinctive repertoire of small non-coding RNAs with potential regulatory functions. Nucleic Acids Res. 2014, 42, 7290–7304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huotari, J.; Helenius, A. Endosome maturation. EMBO J. 2011, 30, 3481–3500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanson, P.I.; Cashikar, A. Multivesicular body morphogenesis. Annu. Rev. Cell Dev. Biol. 2012, 28, 337–362. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, D.L.; Rizzo, J.; Joffe, L.S.; Godinho, R.M.; Rodrigues, M.L. Where do they come from and where do they go: Candidates for regulating extracellular vesicle formation in fungi. Int. J. Mol. Sci. 2013, 14, 9581–9603. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, V. Unconventional protein secretion: An evolving mechanism. EMBO J. 2013, 32, 1660–1664. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C. neoformans | ||||||
|---|---|---|---|---|---|---|
| WT | atg7Δ | graspΔ | ||||
| Uniquely Mapped | % of Total Mapped | Uniquely Mapped | % of Total Mapped | Uniquely Mapped | % of Total Mapped | |
| Exon | 5030 | 0.4 | 60,683 | 9.2 | 59,425 | 7.5 |
| Exon-exon | 10,664 | 0.6 | 1458 | 0.2 | 2350 | 0.3 |
| Total exon | 113,655 | 9.7 | 62,141 | 9.4 | 61,774 | 7.8 |
| Total intron | 1,003,971 | 90.3 | 568,003 | 84.9 | 758,109 | 86.9 |
| Total gene | 1,117,625 | 100 | 667,288 | 100.0 | 861,092 | 100.0 |
| Name | Product | EV vs. Cell-Log Fold Change | EV vs. Cell-FDR p-Value | SRR3199612 Cell 1 RPKM | SRR3199613 Cell 2-RPKM | SRR3199614Cell 3-RPKM | EV RNA 1-RPKM | EV RNA 2-RPKM |
|---|---|---|---|---|---|---|---|---|
| CNAG_03012 | quorum sensing-like molecule | −5.06 | 0.00% | 20,332.13 | 18,844.93 | 20,155.19 | 48.40 | 24.28 |
| CNAG_06207 | hypothetical protein | −6.93 | 0.00% | 16,304.37 | 14,037.19 | 13,815.62 | 8.35 | 6.02 |
| CNAG_04105 | hypothetical protein | −2.31 | 2.29% | 16,003.01 | 10,010.74 | 16,467.89 | 254.98 | 73.62 |
| CNAG_03143 | hypothetical protein | −2.09 | 3.87% | 13,070.91 | 8338.53 | 12,401.56 | 231.90 | 78.39 |
| CNAG_01735 | hypothetical protein | −3.60 | 0.03% | 9034.37 | 6373.02 | 7970.58 | 56.97 | 16.11 |
| CNAG_06075 | hypothetical protein | −2.98 | 0.39% | 6021.45 | 5119.83 | 6051.19 | 66.75 | 13.02 |
| CNAG_03007 | hypothetical protein | −6.58 | 0.00% | 5861.25 | 5356.39 | 4321.23 | 3.72 | 2.64 |
| CNAG_06298 | hypothetical protein | −7.02 | 0.00% | 5319.83 | 5499.72 | 6635.36 | 2.65 | 3.96 |
| CNAG_06101 | ADP, ATP carrier protein | −3.04 | 0.11% | 4475.82 | 5415.80 | 4535.39 | 44.21 | 32.06 |
| CNAG_07466 | U3 small nucleolar RNA-associated protein 7, U3 small nucleolar RNA-associated protein 7, variant 1, U3 small nucleolar RNA-associated protein 7, variant 2 | 10.05 | 0.00% | 392.25 | 765.29 | 487.61 | 39,204.58 | 43,212.28 |
| CNAG_01093 | hypothetical protein | 8.04 | 0.00% | 45.01 | 41.14 | 33.25 | 831.32 | 480.72 |
| CNAG_06651 | amidohydrolase | 12.70 | 0.00% | 3.80 | 4.64 | 3.38 | 777.33 | 4354.12 |
| CNAG_00311 | 3-hydroxyisobutyryl-CoA hydrolase | 6.97 | 0.00% | 62.61 | 78.16 | 54.48 | 650.07 | 373.07 |
| CNAG_02129 | hypothetical protein | 2.12 | 3.76% | 423.90 | 380.09 | 587.72 | 178.91 | 56.55 |
| CNAG_05774 | hypothetical protein, hypothetical protein, variant | 4.48 | 0.00% | 87.07 | 87.32 | 82.46 | 146.53 | 104.07 |
| CNAG_05651 | hypothetical protein | 7.83 | 0.00% | 5.28 | 8.41 | 7.53 | 138.73 | 51.57 |
| CNAG_07515 | hypothetical protein | 4.79 | 0.00% | 57.55 | 52.26 | 77.35 | 118.53 | 127.65 |
| CNAG_04124 | hypothetical protein | 7.66 | 0.00% | 6.60 | 6.26 | 5.59 | 113.90 | 21.83 |
| CNAG_07028 | 26S proteasome regulatory subunit N11 | 4.03 | 0.00% | 103.45 | 112.54 | 86.34 | 112.59 | 118.66 |
| ID | Data obtained from Gonzalez-Hilarion et al., 2016 [38] | RPKM | Unique Exon Reads | Unique Intron Reads | RPKM | Unique Exon Reads | Unique Intron Reads | RPKM | Unique Exon Reads | Unique Intron Reads | Product | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| intron | Type | RPKM | Exons | WT | ΔAtg7 | ΔGRASP | ||||||||
| CNAG_03602 | Ic2-554 | in5UTR | 6.66 | 5 | 23.5 | 4 | 228 | 40.59 | 3 | 87.5 | 28.57 | 1.5 | 81 | U3 small nucleolar RNA-associated protein 5 |
| Ic2-555 | in5UTR | 204.80 | ||||||||||||
| CNAG_03645 | Ic2-787 | inCDS | 8.01 | 8 | 6.3 | 1.5 | 73.5 | 53.43 | 5.5 | 9.5 | 58.98 | 6.5 | 14.5 | NET1-associated nuclear protein 1 (U3 small nucleolar RNA-associated protein 17) |
| Ic2-788 | inCDS | 12.93 | ||||||||||||
| CNAG_04068 | Ic2-3155 | inCDS | 3.18 | 4 | 71.2 | 3.5 | 10 | 270.93 | 6 | 11 | 522.07 | 12 | 4.5 | large subunit ribosomal protein L28e |
| CNAG_07982 | Ic4-247 | inCDS | 6.96 | 5 | 1061.8 | 60.5 | 100.5 | 81.42 | 5 | 3 | 60.25 | 3.5 | 7 | hypothetical protein |
| Ic4-248 | inCDS | 17.47 | ||||||||||||
| CNAG_00930 | Ic4-351 | inCDS | 2.61 | 7 | 50.1 | 5.5 | 328 | 81.47 | 4 | 303.5 | 106.30 | 6 | 397 | argininosuccinate synthase |
| Ic4-349 | in5UTR | 1066.15 | ||||||||||||
| Ic4-350 | in5UTR | 81.59 | ||||||||||||
| CNAG_07884 | Ic8-1359 | inCDS | 10.84 | 3 | 7.4 | 0.5 | 66 | 18.73 | 0.5 | 21.5 | 85.27 | 2.5 | 21 | hypothetical protein |
| CNAG_07813 | Ic12-778 | inCDS | 8.46 | 5 | 75.4 | 6.5 | 231.5 | 14.71 | 0.5 | 141.5 | 37.66 | 1 | 208.5 | hypothetical protein |
| Ic12-776 | in5UTR | 7.38 | ||||||||||||
| CNAG_06167 | Ic13-990 | in5UTR | 26.36 | 5 | 131.1 | 11 | 770 | 103.14 | 6.5 | 152.5 | 146.05 | 10.5 | 295 | metal homeostatis protein bsd2 |
| CNAG_01820 | Ic3-1947 | in3UTR | 28.14 | 12 | 342.4 | 47 | 180.5 | 236.94 | 17 | 15.5 | 344.85 | 27.5 | 15 | pyruvate kinase, pyruvate kinase, variant |
| CNAG_06033 | Ic13-230 | inCDS | 10.93 | 7 | 99.4 | 15 | 22 | 47.14 | 3.5 | 5 | 33.92 | 3.5 | 63.5 | pfkB family carbohydrate kinase superfamily |
| CNAG_03730 | Ic2-1335 | in5UTR | 35.87 | 4 | 41.7 | 2 | 479.5 | 102.24 | 2 | 23.5 | 0.00 | 0 | 175.5 | DNA-directed RNA polymerase II subunit RPB11 |
| CNAG_06401 | Ic14-772 | in5UTR | 14.61 | 11 | 6.1 | 0.5 | 23 | 52.25 | 4.5 | 162.5 | 28.12 | 2.5 | 27.5 | hypothetical protein |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peres da Silva, R.; Martins, S.D.T.; Rizzo, J.; Dos Reis, F.C.G.; Joffe, L.S.; Vainstein, M.; Kmetzsch, L.; Oliveira, D.L.; Puccia, R.; Goldenberg, S.; et al. Golgi Reassembly and Stacking Protein (GRASP) Participates in Vesicle-Mediated RNA Export in Cryptococcus neoformans. Genes 2018, 9, 400. https://doi.org/10.3390/genes9080400
Peres da Silva R, Martins SDT, Rizzo J, Dos Reis FCG, Joffe LS, Vainstein M, Kmetzsch L, Oliveira DL, Puccia R, Goldenberg S, et al. Golgi Reassembly and Stacking Protein (GRASP) Participates in Vesicle-Mediated RNA Export in Cryptococcus neoformans. Genes. 2018; 9(8):400. https://doi.org/10.3390/genes9080400
Chicago/Turabian StylePeres da Silva, Roberta, Sharon De Toledo Martins, Juliana Rizzo, Flavia C. G. Dos Reis, Luna S. Joffe, Marilene Vainstein, Livia Kmetzsch, Débora L. Oliveira, Rosana Puccia, Samuel Goldenberg, and et al. 2018. "Golgi Reassembly and Stacking Protein (GRASP) Participates in Vesicle-Mediated RNA Export in Cryptococcus neoformans" Genes 9, no. 8: 400. https://doi.org/10.3390/genes9080400
APA StylePeres da Silva, R., Martins, S. D. T., Rizzo, J., Dos Reis, F. C. G., Joffe, L. S., Vainstein, M., Kmetzsch, L., Oliveira, D. L., Puccia, R., Goldenberg, S., Rodrigues, M. L., & Alves, L. R. (2018). Golgi Reassembly and Stacking Protein (GRASP) Participates in Vesicle-Mediated RNA Export in Cryptococcus neoformans. Genes, 9(8), 400. https://doi.org/10.3390/genes9080400