
Leaf-Scale Study of Biogenic Volatile Organic Compound Emissions from Willow (Salix spp.) Short Rotation Coppices Covering Two Growing Seasons


Abstract
:1. Introduction
2. Methods
2.1. Site Description
2.2. Experimental Setup
2.2.1. Branch Chamber Setup
2.2.2. Light Response Curves and Photosynthesis Rates
2.3. BVOC Sampling
2.4. Gas Chromatography–Mass Spectrometry and Data Processing
3. Results
3.1. BVOC Emissions from Salix
3.2. Seasonality of BVOC Emissions Measured with Branch Chambers
3.2.1. Isoprene
3.2.2. Monoterpenes
3.2.3. Sesquiterpenes
3.2.4. Other VOCs
3.3. Height Comparison with Branch Chamber Measurements in 2016
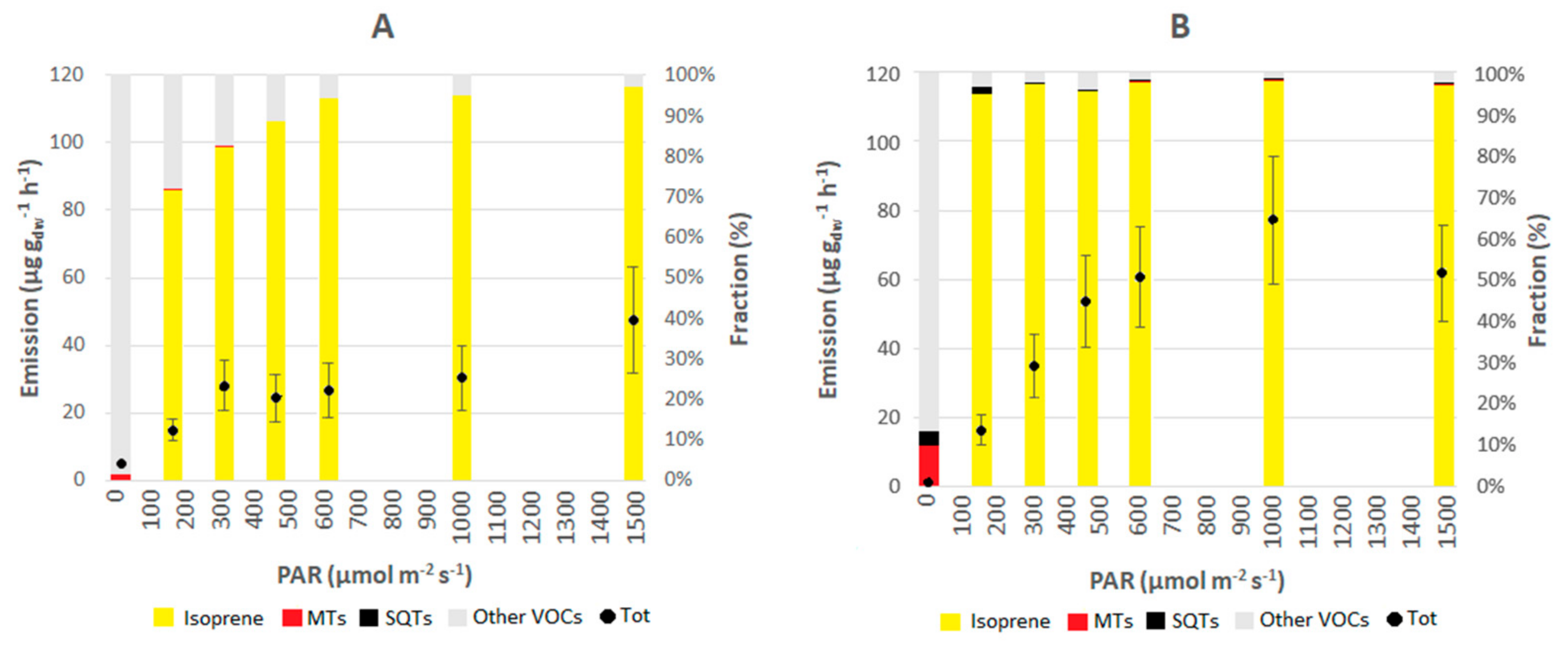
3.4. Light Response Curves with Leaf Chamber
4. Discussion
4.1. Isoprene Emissions, Net Assimilation, and Water Use Efficiency
4.2. Emissions of Monoterpenes, Sesquiterpenes, and Other Volatile Organic Compounds
5. Summary and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- A Policy Framework for Climate and Energy in the Period from 2020 up to 2030; European Commission: Brussels, Belgium, 2014.
- IPCC. 2021: Summary for Policymakers. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge University Press: Cambridge, UK, 2021; In press. [Google Scholar]
- Bonde, I.; Kuylenstierna, J.; Bäckstrand, K.; Eckerberg, K.; Kåberger, T.; Löfgren, Å.; Rummukainen, M.; Sörlin, S. 2020-Report of the Swedish Climate Policy Council; Rapport nr 3; Swedish Climate Policy Council: Stockholm, Sweden, 2020; ISBN 978-91-984671-4-7. [Google Scholar]
- Don, A.; Osborne, B.; Hastings, A.; Skiba, U.; Carter, M.S.; Drewer, J.; Flessa, H.; Freibauer, A.; Hyvönen, N.; Jones, M.B.; et al. Land-use change to bioenergy production in Europe: Implications for the greenhouse gas balance and soil carbon. Glob. Chang. Biol. Bioenergy 2011, 4, 372–391. [Google Scholar] [CrossRef] [Green Version]
- Lindegaard, K.N.; Adams, P.W.; Holley, M.; Lamley, A.; Henriksson, A.; Larsson, S.; von Engelbrechten, H.G.; Esteban Lopez, G.; Pisarek, M. Short rotation plantations policy history in Europe: Lessons from the past and recommendations for the future. Food Energy Secur. 2016, 5, 125–152. [Google Scholar] [CrossRef]
- Kimming, M.; Sundberg, C.; Nordberg, Å.; Baky, A.; Bernesson, S.; Norén, O.; Hansson, P.A. Biomass from agriculture in small-scale combined heat andpower plants—A comparative life cycle assessment. Biomass Bioenerg. 2011, 35, 1572–1581. [Google Scholar] [CrossRef]
- Åhman, I.; Larsson, S. Resistensförädling i Salix för Energiproduktion, SLU; Institutionen för entomologi, Nr/avsnitt: 2; SLU: Alnarp, Sweden, 1999. [Google Scholar]
- Kägi, T.; Deimling, S.; Knuchel, R.F.; Gaillard, G.; Hölscher, T.; Müller-Sämann, K. Environmental impacts of annual and perennial energy crops compared to a reference foo crop rotation. In Proceedings of the Empowerment of the Rural Actors: A Renewal of Farming Systems Perspectives 8th European IFSA Symposium, Clermont-Ferrand, Clermont-Ferrand, France, 6–10 July 2008; pp. 675–681. [Google Scholar]
- Fredga, K.; Danell, K.; Frank, H.; Hedberg, D.; Kullander, S. Bioenergy—Opportunities and constraints. In Energy Committee Report; Kungliga Vetenskapsakademin: Stockholm, Sweden, 2008. [Google Scholar]
- Landberg, T.; Greger, M. Differences in uptake and tolerance to heavy metals in Salix from unpolluted and polluted areas. Appl. Geochem. 1996, 11, 175–180. [Google Scholar] [CrossRef]
- Meers, E.; Vandecasteele, B.; Ruttens, A.; Vangronsveld, J.; Tack, F. Potential of five willow species (Salix spp.) for phytoextraction of heavy metals. Environ. Exp. Bot. 2007, 60, 57–68. [Google Scholar] [CrossRef]
- SOU 2007:36. Bioenergi från Jordbruket–en Växande Resurs; Statens Offentliga Utredningar; Edita Sverige AB: Stockholm, Sweden, 2007; ISBN 978-91-38-22751-0. [Google Scholar]
- Kesselmeier, J.; Staudt, M. Biogenic volatile organic compounds (VOC): An overview on emission, physiology and ecology. J. Atmos. Chem. 1999, 33, 23–88. [Google Scholar] [CrossRef]
- Dicke, M.; Vet, L.E.M. Plant-carnivore carnivore interactions: Evolutionary and ecological consequences for plant, herbivore and carnivore. In Herbivores: Between Plants and Predators; Olff, H., Brown, V.K., Drent, R.H., Eds.; Blackwell Science: Oxford, UK, 1999; pp. 483–520. [Google Scholar]
- Llusià, J.; Peñuelas, J. Emission of volatile organic compounds by apple trees in response to spider mite attack and attraction of predatory mites. Exp. Appl. Acarol. 2001, 25, 65–77. [Google Scholar] [CrossRef] [PubMed]
- Dicke, M.; van Poecke, R.M.P.; de Boer, J.G. Inducible indirect defence of plants: From mechanisms to ecological functions. Basic Appl. Ecol. 2003, 4, 27–42. [Google Scholar] [CrossRef]
- Dudareva, N.; Pichersky, E. Metabolic engineering of plant volatiles. Curr. Opin. Biotechnol. 2008, 19, 1–9. [Google Scholar] [CrossRef]
- Laothawornkitkul, J.; Taylor, J.; Paul, N.; Hewitt, C. Biogenic volatile organic compounds in the Earth system. New Phytol. 2009, 183, 27–51. [Google Scholar] [CrossRef]
- Possell, M.; Loreto, F. The role of volatile organic compounds in plant resistance to abiotic stresses: Responses and mechanisms. In Biology, Controls and Models of Tree Volatile Organic Compound Emissions; Niinemets, Ü., Monson, R.K., Eds.; Tree Physiology; Springer: Berlin/Heidelberg, Germany, 2013; Volume 5, pp. 209–235. [Google Scholar]
- Folberth, G.A.; Hauglustaine, D.A.; Lathière, J.; and Brocheton, F. Interactive chemistry in the Laboratoire de Météorologie Dynamique general circulation model: Model description and impact analysis of biogenic hydrocarbons on tropospheric chemistry. Atmos. Chem. Phys. 2006, 6, 2273–2319. [Google Scholar] [CrossRef] [Green Version]
- Arneth, A.; Monson, R.K.; Schurgers, G.; Niinemets, Ü.; Palmer, P.I. Why are estimates of global terrestrial isoprene emissions so similar (and why is this not so for monoterpenes)? Atmos. Chem. Phys. 2008, 8, 4605–4620. [Google Scholar] [CrossRef] [Green Version]
- Calfapietra, C.; Fares, S.; Manes, F.; Morani, A.; Sgrigna, G.; Loreto, F. Role of biogenic volatile organic compounds (BVOC) emitted by urban trees on ozone concentration in cities: A review. Environ. Pollut. 2013, 183, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Chameides, W.L.; Lindsay, R.W.; Richardson, J. The Role of Biogenic Hydrocarbons in Urban Photochemical Smog: Atlanta as a Case Study. Science 1988, 241, 1473–1475. [Google Scholar] [CrossRef]
- Ryerson, T.B.; Trainer, M.; Holloway, J.S.; Parrish, D.D.; Huey, L.G.; Sueper, D.T.; Frost, G.J.; Donnelly, S.G.; Schauffler, S.; Atlas, E.L.; et al. Observations of Ozone Formation in Power Plant Plumes and Implications for Ozone Control Strategies. Science 2001, 292, 719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleinman, L.I.; Daum, P.H.; Imre, D.; Lee, Y.-N.; Nunnermacker, L.J.; Springston, S.R.; Weinstein-Lloyd, J.; Rudolph, J. Ozone production rate and hydrocarbon reactivity in 5 urban areas: A cause of high ozone concentration in Houston. Geophys. Res. Lett. 2002, 29, 105-1–105-4. [Google Scholar] [CrossRef] [Green Version]
- Fares, S.; Barta, C.; Ederli, L.; Ferranti, F.; Pasqualini, S.; Reale, L.; Brilli, F.; Tricoli, D.; Loreto, F. Impact of high ozone on isoprene emission and some anatomical and physiological parameters of developing Populus alba leaves directly or indirectly exposed to the pollutant. Physiol. Plant. 2006, 128, 456–465. [Google Scholar] [CrossRef]
- Wittig, V.E.; Ainsworth, E.A.; Naidu, S.L.; Karnoski, D.F.; Long, S.P. Quantifying the impact of current and future tropospheric ozone on tree biomass, growth, physiology and biochemistry: A quantitative meta-analysis. Glob. Chang. Biol. 2009, 15, 396–424. [Google Scholar] [CrossRef]
- Kurpius, M.R.; Goldstein, A.H. Gas-phase chemistry dominates O3 loss to a forest, implying a source of aerosols and hydroxyl radicals to the atmosphere. Geophys. Res. Lett. 2003, 30, 1371. [Google Scholar] [CrossRef]
- Agyei, T.; Juráň, S.; Kwakye, K.O.; Šigut, L.; Urban, O.; Marek, M.V. The impact of drought on total ozone flux in a mountain Norway spruce forest. J. For. Sci. 2020, 66, 280–287. [Google Scholar] [CrossRef]
- Ashmore, M.R. Assessing the future global impacts of ozone on vegetation. Plant Cell Environ. 2005, 28, 949–964. [Google Scholar] [CrossRef]
- Sitch, S.; Cox, P.M.; Collins, W.J.; Huntingford, C. Indirect radiative forcing of climate change through ozone effects on the land-carbon sink. Nature 2007, 448, 791–794. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, T.; Odum, J.R.; Bowman, F.; Collins, D.; Klockow, D.; Flagan, R.C.; Seinfeld, J.H. Formation of organic aerosols from the oxidation of biogenic hydrocarbons. J. Atmos. Chem. 1997, 26, 189–222. [Google Scholar] [CrossRef]
- O’Dowd, C.D.; Jimenez, J.L.; Bahreini, R.; Flagan, R.C.; Seinfeld, J.H.; Hämeri, K.; Pirjola, L.; Kulmala, M.; Jennings, S.G.; Hoffmann, T. Marine aerosol formation from biogenic iodine emissions. Nature 2002, 417, 497–501. [Google Scholar] [CrossRef]
- Claeys, M.; Graham, B.; Vas, G.; Wang, W.; Vermeylen, R.; Pashynska, V.; Cafmeyer, J.; Guyon, P.; Andreae, M.O.; Artaxo, P.; et al. Formation of secondary organic aerosols through photooxidation of isoprene. Science 2004, 303, 1173–1176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulmala, M.; Suni, T.; Lehtinen, K.E.J.; Dal Maso, M.; Boy, M.; Reissell, A.; Rannik, Û.; Aalto, P.; Keronen, P.; Hakola, H.; et al. A new feedback mechanism linking forests, aerosols, and climate. Atmos. Chem. Phys. 2004, 4, 557–562. [Google Scholar] [CrossRef] [Green Version]
- VanReken, T.M.; Greenberg, J.P.; Harley, P.C.; Guenther, A.B.; Smith, J.N. Direct measurement of particle formation and growth from the oxidation of biogenic emissions. Atmos. Chem. Phys. 2006, 6, 4403–4413. [Google Scholar] [CrossRef] [Green Version]
- Spracklen, D.V.; Bonn, B.; Carslaw, K.S. Boreal forests, aerosols and the impacts on clouds and climate. Philos. Trans. R. Soc. A 2008, 366, 4613–4626. [Google Scholar] [CrossRef]
- Paasonen, P.; Asmi, A.; Petäjä, T.; Kajos, M.K.; Äijälä, M.; Junninen, H.; Holst, T.; Abbatt, J.P.D.; Arneth, A.; Birmili, W.; et al. Warming-induced increase in aerosol number concentration likely to moderate climate change. Nat. Geosci. 2013, 6, 438–442. [Google Scholar] [CrossRef]
- Ehn, M.; Thornton, J.A.; Kleist, E.; Sipilä, M.; Junninen, H.; Pullinen, I.; Springer, M.; Rubach, F.; Tillmann, R.; Lee, B.; et al. A large source of low-volatility secondary organic aerosol. Nature 2014, 506, 476–479. [Google Scholar] [CrossRef]
- Boucher, O.; Randall, P.D.; Artaxo, C.; Bretherton, G.; Feingold, P.; Forster, V.-M.; Kerminen, Y.; Kondo, H.; Liao, U.; Lohmann, P.; et al. 2013: Clouds and Aerosols. In Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.-K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013. [Google Scholar]
- Vedel-Petersen, I.; Schollert, M.; Nymand, J.; Rinnan, R. Volatile organic compound emission profiles of four common arctic Plants. Atmos. Environ. 2015, 120, 117–126. [Google Scholar] [CrossRef]
- Karlsson, T.; Rinnan, R.; Holst, T. Variability of BVOC Emissions from Commercially Used Willow (Salix spp.) Varieties. Atmosphere 2020, 11, 356. [Google Scholar] [CrossRef] [Green Version]
- Owen, S.M.; Boissard, C.; Hewitt, C.N. Volatile organic compounds (VOCs) emitted from 40 Mediterranean plant species: VOC speciation and extrapolation to habitat scale. Atmos. Environ. 2001, 35, 5393–5409. [Google Scholar] [CrossRef]
- Olofsson, M.; Ekolausson, B.; Jensen, N.; Langer, S.; Ljungström, E. The flux of isoprene from a willow coppice plantation and the effect on local air quality. Atmos. Environ. 2005, 39, 2061–2070. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Copolovici, L.; Hüve, K. High within-canopy variation in isoprene emission potentials in temperate trees: Implications for predicting canopy-scale isoprene fluxes. J. Geophys. Res. 2010, 115, G04029. [Google Scholar] [CrossRef] [Green Version]
- Persson, Y.; Schurgers, G.; Ekberg, A.; Holst, T. Effects of intra-genotypic variation, variance with height and time of season on BVOC emissions. Meteorol. Z. 2016, 25, 377–388. [Google Scholar] [CrossRef]
- Alexandersson, H.; Eggertsson Karlström, C. Temperaturen och nederbörden i Sverige 1961–90: Referensnormaler—Utgåva 2. Meteorologi 99; Swedish Meteorological and Hydrological Institute: Norrköping, Sweden, 2001. (In Swedish) [Google Scholar]
- SGU, Geology Survey of Sweden. Available online: http://maps-test.sgu.se:8080/TestSguMapViewer2/kartvisare-lerhaltskarta-sv.html (accessed on 7 March 2019).
- Haapanala, S.; Ekberg, A.; Hakola, A.; Tarvainen, V.; Rinne, J.; Hellén, H.; Arneth, A. Mountain birch—Potentially large source of sesquiterpenes into high latitude atmosphere. Biogeosciences 2009, 6, 2709–2718. [Google Scholar] [CrossRef] [Green Version]
- van Meeningen, Y.; Wang, M.; Karlsson, T.; Seifert, A.; Schurgers, G.; Rinnan, R.; Holst, T. Isoprenoid emission variation of Norway spruce across a European latitudinal transect. Atmos. Environ. 2017, 170, 45–57. [Google Scholar] [CrossRef]
- Wang, M.; Schurgers, G.; Arneth, A.; Ekberg, A.; Holst, T. Seasonal variation in biogenic volatile organic compound (BVOC) emissions from Norway spruce in a Swedish boreal forest. Boreal Environ. Res. 2017, 22, 353–367. [Google Scholar]
- van Meeningen, Y.; Schurgers, G.; Rinnan, R.; Holst, T. Isoprenoid emission response to changing light conditions of English oak, European beech and Norway spruce. Biogeosciences 2017, 14, 4045–4060. [Google Scholar] [CrossRef] [Green Version]
- Lindwall, F.; Svendsen, S.S.; Nielsen, C.S.; Michelsen, A.; Rinnan, R. Warming in-creases isoprene emissions from an arctic fen. Sci. Total Environ. 2016, 553, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Ortega, J.; Helmig, D. Approaches for quantifying reactive and low-volatility biogenic organic compound emissions by vegetation enclosure techniques—Part A. Chemosphere 2008, 72, 343–364. [Google Scholar] [CrossRef]
- Guenther, A.B.; Zimmerman, P.; Harley, P.C.; Monson, R.K.; Fall, R. Isoprene and monoterpene emission rate variability: Model evaluations and sensitivity analyses. J. Geophys. Res. 1993, 98, 12609–12617. [Google Scholar] [CrossRef] [Green Version]
- Helmig, D.; Ortega, J.; Duhl, T.; Tanner, D.; Guenther, A.; Harley, P.; Wiedinmyer, C.; Milford, J.; Sakulyanontvittaya, T. Sesquiterpene Emissions from Pine Trees—Identifications, Emission Rates and Flux Estimates for the Contiguous United States. Environ. Sci. Technol. 2007, 41, 1545–1553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niinemets, Ü.; Sun, Z.; Talts, E. Controls of the quantum yield and saturation light of isoprene emission in different-aged aspen leaves. Plant Cell Environ. 2015, 38, 2707–2720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Owen, S.M.; Hewitt, C.N. Extrapolating branch enclosure measurements to estimates of regional scale biogenic VOC fluxes in the north-western Mediterranean basin. J. Geophys. Res. 2000, 105, 11573–11583. [Google Scholar] [CrossRef]
- Morrison, E.C.; Drewer, J.; Heal, M.R. A comparison of isoprene and monoterpene emission rates from the perennial bioenergy crops short-rotation coppice willow and Miscanthus and the annual arable crops wheat and oilseed rape. GCB Bioenergy 2016, 8, 211–225. [Google Scholar] [CrossRef] [Green Version]
- Monson, R.K.; Jaeger, C.H.; Adams, W.; Driggers, E.; Silver, G.; Fall, R. Relationships among isoprene emission rate, photosynthesis rate, and isoprene synthase activity, as influenced by temperature. Plant Physiol. 1992, 92, 1175–1180. [Google Scholar] [CrossRef] [Green Version]
- Kuzma, J.; Fall, R. Leaf isoprene emission rate is dependent on leaf development and the level of isoprene synthase. Plant Physiol. 1993, 101, 435–440. [Google Scholar] [CrossRef] [Green Version]
- Monson, R.K.; Harley, P.C.; Litvak, M.E.; Wildermuth, M.; Guenther, A.B.; Zimmerman, P.R.; Fall, R. Environmental and developmental controls over the seasonal pattern of isoprene emission from aspen leaves. Oecologia 1994, 99, 260–270. [Google Scholar] [CrossRef]
- Schnitzler, J.-P.; Lehning, A.; Steinbrecher, R. Seasonal Pattern of Isoprene Synthase Activity in Quercus robur Leaves and its Significance or Modeling Isoprene Emission Rates. Bot. Acta 1997, 110, 240–243. [Google Scholar] [CrossRef]
- Toome, M.; Randjärv, P.; Copolovici, L.; Niinemets, Ü.; Heinsoo, K.; Luik, A.; Noe, S.M. Leaf rust induced volatile organic compounds signalling in willow during the infection. Planta 2010, 232, 235–243. [Google Scholar] [CrossRef]
- Jiang, Y.; Ye, J.; Veromann, L.-L.; Niinemets, Ü. Scaling of photosynthesis and constitutive and induced volatile emissions with severity of leaf infection by rust fungus (Melampsora larici-populina) in Populus balsamifera var. suaveolens. Tree Physiol. 2016, 36, 856–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharkey, T.D.; Loreto, F.; Delwiche, C.F. High carbon dioxide and sun/shade effects on isoprene emission from oak and aspen tree leaves. Plant Cell Environ. 1991, 14, 333–338. [Google Scholar] [CrossRef]
- Lerdau, M.; Throop, H.L. Sources of variability in isoprene emission and photosynthesis in two species of tropical wet forest trees. Biotropica 2000, 32, 670–676. [Google Scholar] [CrossRef]
- Monson, R.K.; Fall, R. Isoprene emission from aspen leaves. Plant Physiol. 1989, 90, 267–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guenther, A.B.; Monson, R.K.; Fall, R. Isoprene and monoterpene emission rate variability: Observations with eucalyptus and emission rate algorithm development. J. Geophys. Res. 1991, 96, 10799–10808. [Google Scholar] [CrossRef]
- Sharkey, T.D.; Monson, R.K. Isoprene research—60 years later, the biology is still enigmatic. Plant Cell Environ. 2017, 40, 1671–1678. [Google Scholar] [CrossRef]
- Simin, T.; Tang, J.; Holst, T.; Rinnan, R. Volatile organic compound emission in tundra shrubs—Dependence on species characteristics and the near-surface environment. Environ. Exp. Bot. 2021, 184, 104387. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Tenhunen, J.D.; Harley, P.C.; Steinbrecher, R. A model of isoprene emission based on energetic requirements for isoprene synthesis and leaf photosynthetic properties for Liquidambar and Quercus. Plant Cell Environ. 1999, 22, 1319–1335. [Google Scholar] [CrossRef]
- Kenzo, T.; Ichie, T.; Watanabe, Y.; Yoneda, R.; Ninomiya, I.; Koike, T. Changes in photosynthesis and leaf characteristics with tree height in five dipterocarp species in a tropical rain forest. Tree Physiol. 2006, 26, 865–873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosugi, Y.; Takanashi, S.; Yokoyama, N.; Philip, E.; Kamakura, M. Vertical variation in leaf gas exchange parameters for a Southeast Asian tropical rainforest in Peninsular Malaysia. J. Plant Res. 2012, 125, 735–748. [Google Scholar] [CrossRef] [PubMed]
- Juráň, S.; Pallozzi, E.; Guidolotti, G.; Fares, S.; Šigut, L.; Calfapietra, C.; Alivernini, A.; Savi, F.; Večeřová, K.; Křůmal, K.; et al. Fluxes of biogenic volatile organic compounds above temperate Norway spruce forest of the Czech Republic. Agric. For. Meteorol. 2017, 232, 500–513. [Google Scholar] [CrossRef]
- Guidolotti, G.; Calfapietra, C.; Loreto, F. The relationship between isoprene emission, CO2 assimilation and water use efficiency across a range of poplar genotypes. Physiol. Plant. 2011, 142, 297–304. [Google Scholar] [CrossRef]
- Tingey, D.T.; Manning, M.; Grothaus, L.C.; Burns, W.F. The influence of light and temperature on isoprene emission rates from live oak. Physiol. Plant. 1979, 47, 112–118. [Google Scholar] [CrossRef]
- Llusià, J.; Peñuelas, J. Seasonal patterns of terpene content and emission from seven Mediterranean woody species in field conditions. Am. J. Bot. 2000, 87, 133–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kesselmeier, J.; Ciccioli, P.; Kuhn, U.; Stefani, P.; Biesenthal, T.; Rottenberger, S.; Wolf, A.; Vitullo, M.; Valentini, R.; Nobre, A.; et al. Volatile organic compound emissions in relation to plant carbon fixation and the terrestrial carbon budget. Glob. Biogeochem. Cycles 2002, 16, 1126. [Google Scholar] [CrossRef]
- Bracho-Nunez, A.; Knothe, N.M.; Welter, S.; Staudt, M.; Costa, W.R.; Liberato, M.A.R.; Piedade, M.T.F.; Kesselmeier, J. Leaf level emissions of volatile organic compounds (VOC) from some Amazonian and Mediterranean plants. Biogeosciences 2013, 10, 5855–5873. [Google Scholar] [CrossRef] [Green Version]
- Loreto, F.; Sharkey, T.D. A gas-exchange study of photosynthesis and isoprene emission in Quercus rubra L. Planta 1990, 182, 523–531. [Google Scholar] [CrossRef]
- Larcher, W. Carbon Utilization and Dry Matter Production. In Physiological Plant Ecology: Ecophysiology and Stress Physiology of Functional Groups, 4th ed.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 2003; pp. 69–184. [Google Scholar]
- Winer, A.M.; Arey, J.; Atkinson, R.; Aschmann, S.M.; Long, W.D.; Morrison, C.L.; Olszyk, D.M. Emission rates of organics from vegetation in California’s central valley. Atmos. Environ. 1992, 26, 2647–2659. [Google Scholar] [CrossRef]
- König, G.; Brunda, M.; Puxbaum, H.; Hewitt, C.N.; Duckham, S.C.; Rudolph, J. Relative contribution of oxygenated hydrocarbons to the total biogenic VOC emissions of selected mid-European agricultural and natural plant species. Atmos. Environ. 1995, 29, 861–874. [Google Scholar] [CrossRef]
- Karl, M.; Guenther, A.; Köble, R.; Leip, A.; Seufert, G. A new European plantspecific emission inventory of biogenic volatile organic compounds for use in atmospheric transport models. Biogeosciences 2009, 6, 1059–1087. [Google Scholar] [CrossRef]
- Roberts, J.M.; Williams, J.; Baumann, K.; Buhr, M.P.; Goldan, P.D.; Holloway, J.; Hubler, G.; Kuster, W.C.; McKeen, S.A.; Ryerson, T.B.; et al. Measurements of PAN, PPN, MPAN made during the 1994 and 1995 Nashville Intensives of the Southern Oxidant Study: Implications for regional ozone production from biogenic hydrocarbons. J. Geophys. Res. 1998, 103, 22473–22490. [Google Scholar] [CrossRef]
- Wang, K.-Y.; Shallcross, D.E. Modelling terrestrial biogenic isoprene fluxes and their potential impact on global chemical species using a coupled LSM-CTM model. Atmos. Environ. 2000, 34, 2909–2925. [Google Scholar] [CrossRef]
- Watson, L.A.; Wang, K.-Y.; Paul, H.; Shallcross, D.E. The potential impact of biogenic emissions of isoprene on urban chemistry in the United Kingdom. Atmos. Sci. Lett. 2006, 7, 96–100. [Google Scholar] [CrossRef]
- Hakola, H.; Rinne, J.; Laurila, T. The VOC emission rates of boreal deciduous trees. In Biogenic VOC Emissions and Photochemistry in the Boreal Regions of Europe-Biphorep; European Commission: Brussels, Belgium, 1999. [Google Scholar]
- Staudt, M.; Seufert, G. Light-dependent emission of monoterpenes by Holm oak (Quercus ibex L.). Naturwissenschaften 1995, 82, 89–92. [Google Scholar] [CrossRef]
- Kesselmeier, J.; Schäfer, L.; Ciccioli, P.; Brancaleoni, E.; Cecinato, A.; Frattoni, M.; Foster, P.; Jacob, V.; Denis, J.; Fugit, J.-L.; et al. Emission of monoterpenes and isoprene from a Mediterranean oak species Quercus ilex L. measured within the BEMA (Biogenic Emissions in the Mediterranean Area) project. Atmos. Environ. 1996, 30, 1841–1850. [Google Scholar] [CrossRef]
- Peñuelas, J.; Llusià, J. Short-term responses of terpene emission rates to experimental changes of PFD in Pinus halepensis and Quercus ilex in summer field conditions. Environ. Exp. Bot. 1999, 42, 61–68. [Google Scholar] [CrossRef]
- Loreto, F.; Forster, A.; Durr, M.; Csiky, O.; Seufert, G. On the monoterpene emission under heat stress and on the increased thermotolerance of leaves of Quercus ilex L. fumigated with selected monoterpenes. Plant Cell Environ. 1998, 21, 101–107. [Google Scholar] [CrossRef]
- Paré, P.W.; Tumlinson, J.H. Plant volatiles as a defense against insect herbivores. Plant Physiol. 1999, 121, 325–332. [Google Scholar] [CrossRef] [Green Version]
- Delfine, S.; Csiky, O.; Seufert, G.; Loreto, F. Fumigation with exogenous monoterpenes of a non-isoprenoid-emitting oak (Quercus suber): Monoterpene acquisition, translocation, and effect on the photosynthetic properties at high temperatures. New Phytol. 2000, 146, 27–36. [Google Scholar] [CrossRef]
- Peñuelas, J.; Llusià, J. Linking photorespiration, monoterpenes and thermotolerance in Quercus. New Phytol. 2002, 155, 227–237. [Google Scholar] [CrossRef] [Green Version]
- Copolovici, L.; Kännaste, A.; Pazouki, L.; Niinemets, Ü. Emissions of green leaf volatiles and terpenoids from Solanum lycopersicum are quantitatively related to the severity of cold and heat shock treatments. J. Plant Physiol. 2012, 169, 664–672. [Google Scholar] [CrossRef]
- Palmer-Young, E.C.; Veit, D.; Gershenzon, J.; Schuman, M.C. The Sesquiterpenes (E)-ß-Farnesene and (E)-α-Bergamotene Quench Ozone but Fail to Protect the Wild Tobacco Nicotiana attenuata from Ozone, UVB, and Drought Stresses. PLoS ONE 2015, 10, e0127296. [Google Scholar]
- Pazouki, L.; Kanagendran, A.; Li, S.; Kännaste, A.; Memari, H.R.; Bichele, R.; Niinemets, Ü. Mono- and sesquiterpene release from tomato (Solanum lycopersicum) leaves upon mild and severe heat stress and through recovery: From gene expression to emission responses. Environ. Exp. Bot. 2016, 132, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Tiiva, P.; Häikiö, E.; Kasurinen, A. Impact of warming, moderate nitrogen addition and bark herbivory on BVOC emissions and growth of Scots pine (Pinus sylvestris L.) seedlings. Tree Physiol. 2018, 38, 1461–1475. [Google Scholar] [CrossRef] [PubMed]
- Hildebrand, D.F. Lipoxygenases. Physiol. Plant. 1989, 76, 249–253. [Google Scholar] [CrossRef]
- Andersen, R.A.; Hamilton-Kemp, T.R.; Hildebrand, D.F.; McCracken, C.T.; Collins, R.W.; Fleming, P.D. Structure-Antifungal Activity Relationships among Volatile C6 and C9 Aliphatic Aldehydes, Ketones, and Alcohols. J. Agric. Food Chem. 1994, 42, 1563–1568. [Google Scholar] [CrossRef]
- Wildt, J.; Kobel, K.; Schuh-Thomas, G.; Heiden, A. Emissions of Oxygenated Volatile Organic Compounds from Plants Part II: Emissions of Saturated Aldehydes. J. Atmos. Chem. 2003, 45, 173–196. [Google Scholar] [CrossRef]
- Misztal, P.K.; Hewitt, C.N.; Wildt, J.; Blande, J.D.; Eller, A.; Fares, S.; Gentner, D.; Gilman, J.; Graus, M.; Greenberg, J.; et al. Atmospheric benzenoid emissions from plants rival those from fossil fuels. Sci. Rep. 2015, 5, 12064. [Google Scholar] [CrossRef] [Green Version]
- Arimura, G.; Huber, D.P.; Bohlmann, J. Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnal emissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides): cDNA cloning, functional characterization, and patterns of gene expression of (–)-germacrene D synthase, PtdTPS1. Plant J. 2004, 37, 603–616. [Google Scholar] [PubMed]
- Kleist, E.; Mentel, T.F.; Andres, S.; Bohne, A.; Folkers, A.; Kiendler-Scharr, A.; Rudich, Y.; Springer, M.; Tillmann, R.; Wildt, J. Irreversible impacts of heat on the emissions of monoterpenes, sesquiterpenes, phenolic BVOC and green leaf volatiles from several tree species. Biogeosciences 2012, 9, 5111–5123. [Google Scholar] [CrossRef] [Green Version]
- Hakola, H.; Rinne, J.; Laurila, T. The hydrocarbon emission rates of tea-leafed willow (Salix phylicifolia), silver birch (Betula pendula) and European aspen (Populus tremula). Atmos. Environ. 1998, 32, 1825–1833. [Google Scholar] [CrossRef]
- Copeland, N.; Cape, J.N.; Heal, M.R. Volatile organic compound emissions from Miscanthus and short rotation coppice willow bioenergy crops. Atmos. Environ. 2012, 60, 327–335. [Google Scholar] [CrossRef] [Green Version]
- Peräkylä, O.; Vogt, M.; Tikkanen, O.P.; Laurila, T.; Kajos, M.K.; Rantala, P.A.; Patokoski, J.; Aalto, J.; Yli-Juuti, T.; Ehn, M.; et al. Monoterpenes oxidation capacity and rate over a boreal forest: Temporal variation and connection to growth of newly formed particles. Boreal Environ. Res. 2014, 19, 293–310. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| P1 | P2 | |
|---|---|---|
| Size | 6 ha | 5 ha |
| Formerly used for | cereals | Salix |
| Established | 1994 | 2003 |
| Canopy height 1 | 4.5 m | 7 m |
| LAI 2 | 5.5 | 4.9 |
| Soil type 3 | silty clay loam | clay loam |
| Clay content 3 | 36% | 34% |
| Silt content 3 | 46% | 45% |
| Organic content 3 | 0% | 0% |
| Treatments 4 | Fertilized (ca 2 × 100 kg N ha−1) | Fertilized (ca 1 × 100 kg N ha−1) |
| Varieties | unknown S. viminalis | Tora |
| Last harvest | 2013 | 2013 |
| Yield 5 | ca 200 m3 ha−1 | ca 200 m3 ha−1 |
| Branch Chamber | ||||||
| Campaign No. | Time | Relative Canopy Height (z/H) | Height Group | Samples | Trees | Plantation |
| 1 | 2015: 16–17 July | 1.00 | zH | 8 | 1 | P1 |
| 2 | 2015: 28–31 July | 1.00 | zH | 30 | 1 | P1 |
| 3 | 2015: 4–7 Aug. | 1.00 | zH | 28 | 1 | P1 |
| 4 | 2015: 18–21 Aug. | 1.00 | zH | 31 | 1 | P1 |
| 5 | 2015: 1–4 Sep. | 0.85 | zH | 31 | 1 | P1 |
| 6 | 2015: 8 and 10–11 Sep. | 0.35; 0.80 | zM; zH | 4; 10 | 2 | P1* & P2 |
| Tot: 142 | Tot: 7 | |||||
| Branch Chamber | ||||||
| Campaign No. | Time | Relative Canopy Height (z/H) | Height Group | Samples | Trees | Plantation |
| 1 | 2016: 4–5 May | 0.40 | zM | 14 | 1 | P2 |
| 2 | 2016: 24 May | 0.75 | zH | 6 | 1 | P2 |
| 3 | 2016: 6–10 June | 0.30; 0.75 | zM; zH | 39; 10 | 2 | P2 |
| 4 | 2016: 19–23 June | 0.30; 0.75 | zM; zH | 39; 40 | 2 | P2 |
| 5 | 2016: 11–15 July | 0.30; 0.80 | zM; zH | 36; 36 | 2 | P2 |
| 6 | 2016: 26–29 July | 0.70; 0.90 | zM; zH | 32; 30 | 2 | P2 |
| 7 | 2016: 15–17 Aug. | 0.55; 0.90 | zM; zH | 20; 12 | 2 | P2 |
| 8 | 2016: 6–9 Sep. | 0.65; 0.90 | zM; zH | 30; 28 | 2 | P2 |
| 9 | 2016: 10–13 Oct. | 0.50; 0.95 | zM; zH | 29; 26 | 2 | P2 |
| Tot: 427 | Tot: 16 | |||||
| Leaf Chamber | ||||||
| Campaign No. | Time | Relative Canopy Height (z/H) | Height Group | Samples | Trees | Plantation |
| 5 | 2016: 11–15 July | 0.15; 0.15; 0.40 | zM; zM; zM | 7; 7; 4 | 3 | P2 |
| 6 | 2016: 26–29 July | 0.15; 0.60; 0.80; 0.80 | zM; zL; zH; zH | 7; 7; 7; 7 | 3 | P2 |
| 7 | 2016: 15–17 Aug. | 0.20; 0.65; 0.90 | zM; zL; zH | 6; 7; 7 | 3 | P2 |
| 8 | 2016: 6–9 Sep. | 0.65; 0.20; 0.95; 0.95 | zL; zM; zH; zH | 7; 7; 7; 7 | 3 | P2 |
| Tot: 94 | Tot: 12 | |||||
| BVOC | E (µg gdw−1 h−1) | ES (µg gdw−1 h−1) |
|---|---|---|
| isoprene | 23.5 (28.1) | 45.2 (42.9) |
| Total MTs | 0.163 (0.117) | 0.301 (0.201) |
| ocimene | 0.137 (0.321) | 0.255 (0.540) |
| limonene * | 0.008 (0.042) | 0.014 (0.081) |
| p-cymene | 0.006 (0.042) | 0.011 (0.069) |
| linalool | 0.006 (0.017) | 0.010 (0.035) |
| α-pinene | 0.003 (0.021) | 0.005 (0.034) |
| 3-carene * | 0.002 (0.019) | 0.004 (0.040) |
| Total SQTs | 0.035 (0.062) | 0.103 (0.249) |
| caryophellene † | 0.011 (0.031) | 0.024 (0.080) |
| humulene | 0.010 (0.090) | 0.040 (0.409) |
| α-farnesene † | 0.009 (0.062) | 0.017 (0.125) |
| Total other VOCs | 0.751 (0.159) | − |
| cyclopentyl acetylene § | 0.083 (0.205) | − |
| benzaldehyde (benzenoid) | 0.065 (0.163) | − |
| hexanal (aldehyde) | 0.064 (0.320) | − |
| nonanal (aldehyde) | 0.061 (0. 147) | − |
| 2-ethylhexanoic acid § (carboxylic acid) | 0.059 (0.252) | − |
| pentanal § (aldehyde) | 0.037 (0.226) | − |
| decanal § (aldehyde) | 0.036 (0.148) | − |
| octanal (aldehyde) | 0.030 (0.105) | − |
| 2-methylbutane § (alkane) | 0.027 (0.105) | − |
| 2-pentanone § (ketone) | 0.026 (0.208) | − |
| 2016 zH. | May | June | July | Aug | Sep | Oct |
|---|---|---|---|---|---|---|
| isoprene | 44.8 (99.1%) | 32.6 (95.8%) | 37.5 (98.3%) | 11.1 (97.8%) | 18.2 (98.1%) | 0.2 (87.3%) |
| MTs (tot) | 0.127 (0.3%) | 1.240 (3.6%) | 0.371 (1.0%) | 0.071 (0.6%) | 0.275 (1.5%) | 0 |
| α-pinene | 0 | <0.001 (<0.1%) | 0.001 (0.3%) | 0.001 (1.4%) | 0.001 (0.4%) | 0 |
| myrcene | 0 | <0.001 (<0.1%) | 0 | 0 | 0 | 0 |
| 3-carene | 0.008 (6.3%) | 0 | <0.001 (0.1%) | 0.001 (1.4%) | 0 | 0 |
| ocimene | 0.004 (3.2%) | 1.200 (96.8%) | 0.358 (96.4%) | 0.038 (53.6%) | 0.257 (93.4%) | 0 |
| limonene | 0.004 (3.2%) | <0.001 (<0.1%) | 0.003 (0.8%) | 0 | 0 | 0 |
| linalool | 0.104 (81.7%) | 0.033 (2.7%) | 0.007 (1.9%) | 0.028 (39.4%) | 0.016 (5.8%) | 0 |
| p-cymene | 0.003 (2.4%) | <0.001 (<0.1%) | <0.001 (0.2%) | 0.001 (1.4%) | 0 | 0 |
| allo-ocimene | 0 | <0.001 (<0.1%) | 0 | 0 | 0 | 0 |
| eucalyptol | 0.004 (3.2%) | 0.004 (0.3%) | 0.001 (0.3%) | 0.002 (2.8%) | 0.001 (0.4%) | 0 |
| SQTs (tot) | 0 | 0.010 (<0.1%) | 0.043 (0.1%) | 0 | 0.007 (<0.1%) | 0 |
| α-farnesene | 0 | 0 | 0.002 (4.7%) | 0 | 0.007 (100%) | 0 |
| humulene | 0 | <0.001 (10.0%) | 0.005 (11.6%) | 0 | 0 | 0 |
| caryophyllene | 0 | 0.009 (90.0%) | 0.036 (83.7%) | 0 | 0 | 0 |
| other VOCs (tot) | 0.262 (0.6%) | 0.172 (0.5%) | 0.228 (0.6%) | 0.176 (1.6%) | 0.067 (0.4%) | 0.029 (12.7%) |
| BVOCs (tot) | 45.189 (100%) | 34.021 (100%) | 38.140 (100%) | 11.347 (100%) | 18.549 (100%) | 0.229 (100%) |
| TC | 26.9 (3.5) | 24.5 (6.0) | 25.5 (4.8) | 31.1 (6.4) | 25.7 (4.4) | 8.8 (2.9) |
| PARC | 521 (149) | 727 (494) | 629 (365) | 833 (445) | 313 (235) | 96 (126) |
| 2016 zM | May | June | July | Aug | Sep | Oct |
|---|---|---|---|---|---|---|
| isoprene | 23.5 (95.3%) | 48.7 (99.8%) | 34.0 (97.4%) | 60.5 (98.9%) | 9.6 (96.0%) | 2.8 (93.6%) |
| MTs (tot) | 0.788 (3.2%) | 0.005 (<0.1%) | 0.147 (0.4%) | 0.062 (0.1%) | 0.012 (0.1%) | 0.042 (1.4%) |
| α-pinene | 0 | <0.001 (15.9%) | 0.004 (2.7%) | 0 | 0 | 0.007 (16.7%) |
| myrcene | 0 | 0 | 0 | 0 | 0 | 0 |
| 3-carene | 0.002 (0.3%) | <0.001 (6.2%) | 0.005 (3.4%) | 0 | 0.003 (25.0%) | 0.014 (33.3%) |
| ocimene | 0.701 (89.0%) | 0.003 (55.9%) | 0.121 (82.3%) | 0.057 (91.9%) | 0 | 0 |
| limonene | 0 | 0 | 0.001 (0.7%) | 0 | 0.003 (25.0%) | 0.021 (50.0%) |
| linalool | 0.071 (9.0%) | 0 | <0.001 (<0.1%) | 0.005 (8.1%) | 0 | 0 |
| p-cymene | 0.001 (0.1%) | 0.001 (22.0%) | 0.006 (4.1%) | 0 | 0.004 (33.3%) | 0 |
| allo-ocimene | 0 | 0 | 0 | 0 | 0 | 0 |
| eucalyptol | 0.013 (1.6%) | 0 | 0.010 (6.8%) | 0 | 0.002 (16.7%) | 0 |
| SQTs (tot) | 0 | 0 | 0 | 0.086 (0.1%) | 0 | 0 |
| α-farnesene | 0 | 0 | 0 | 0.021 (24.4%) | 0 | 0 |
| humulene | 0 | 0 | 0 | 0.009 (10.5%) | 0 | 0 |
| caryophyllene | 0 | 0 | 0 | 0.056 (65.1%) | 0 | 0 |
| other VOCs (tot) | 0.359 (1.5%) | 0.077 (0.2%) | 0.761 (2.2%) | 0.531 (0.9%) | 0.386 (3.9%) | 0.149 (5.0%) |
| BVOCs (tot) | 24.647 (100%) | 48.782 (100%) | 34.908 (100%) | 61.179 (100%) | 9.998 (100%) | 2.991 (100%) |
| TC | 25.7 (1.7) | 25.4 (5.6) | 23.1 (3.9) | 31.1 (7.0) | 24.3 (4.7) | 8.6 (2.5) |
| PARC | 1135 (406) | 699 (469) | 298 (254) | 627 (450) | 486 (342) | 83 (123) |
| PAR = 1500 μmol m−2 s−1 | Lower (zL) | Middle (zM) | Higher (zH) |
|---|---|---|---|
| A (µmol CO2 m−2 s−1) | 12.05 (3.01) | 21.30 (2.92) | 18.10 (4.89) |
| Tr (mmol H2O m−2 s−1) | 2.51 (0.43) | 2.24 (0.97) | 2.98 (0.47) |
| WUE (mmol CO2 mol−1 H2O) | 4.73 (0.55) | 12.99 (9.28) | 6.10 (2.13) |
| gs (mol H2O m−2 s−1) | 0.13 (0.03) | 0.17 (0.04) | 0.18 (0.05) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karlsson, T.; Klemedtsson, L.; Rinnan, R.; Holst, T. Leaf-Scale Study of Biogenic Volatile Organic Compound Emissions from Willow (Salix spp.) Short Rotation Coppices Covering Two Growing Seasons. Atmosphere 2021, 12, 1427. https://doi.org/10.3390/atmos12111427
Karlsson T, Klemedtsson L, Rinnan R, Holst T. Leaf-Scale Study of Biogenic Volatile Organic Compound Emissions from Willow (Salix spp.) Short Rotation Coppices Covering Two Growing Seasons. Atmosphere. 2021; 12(11):1427. https://doi.org/10.3390/atmos12111427
Chicago/Turabian StyleKarlsson, Tomas, Leif Klemedtsson, Riikka Rinnan, and Thomas Holst. 2021. "Leaf-Scale Study of Biogenic Volatile Organic Compound Emissions from Willow (Salix spp.) Short Rotation Coppices Covering Two Growing Seasons" Atmosphere 12, no. 11: 1427. https://doi.org/10.3390/atmos12111427
APA StyleKarlsson, T., Klemedtsson, L., Rinnan, R., & Holst, T. (2021). Leaf-Scale Study of Biogenic Volatile Organic Compound Emissions from Willow (Salix spp.) Short Rotation Coppices Covering Two Growing Seasons. Atmosphere, 12(11), 1427. https://doi.org/10.3390/atmos12111427