
Species of Fungi and Pollen in the PM1 and the Inhalable Fraction of Indoor Air in Homes


Abstract
:1. Introduction
2. Materials and Methods
2.1. Selection of Homes and Sampling of the Inhalable and PM1 Fraction in Five Homes
2.2. Extraction of Filters and Plating
2.3. qPCR of the Inhalable (GSP) and PM1 Fraction
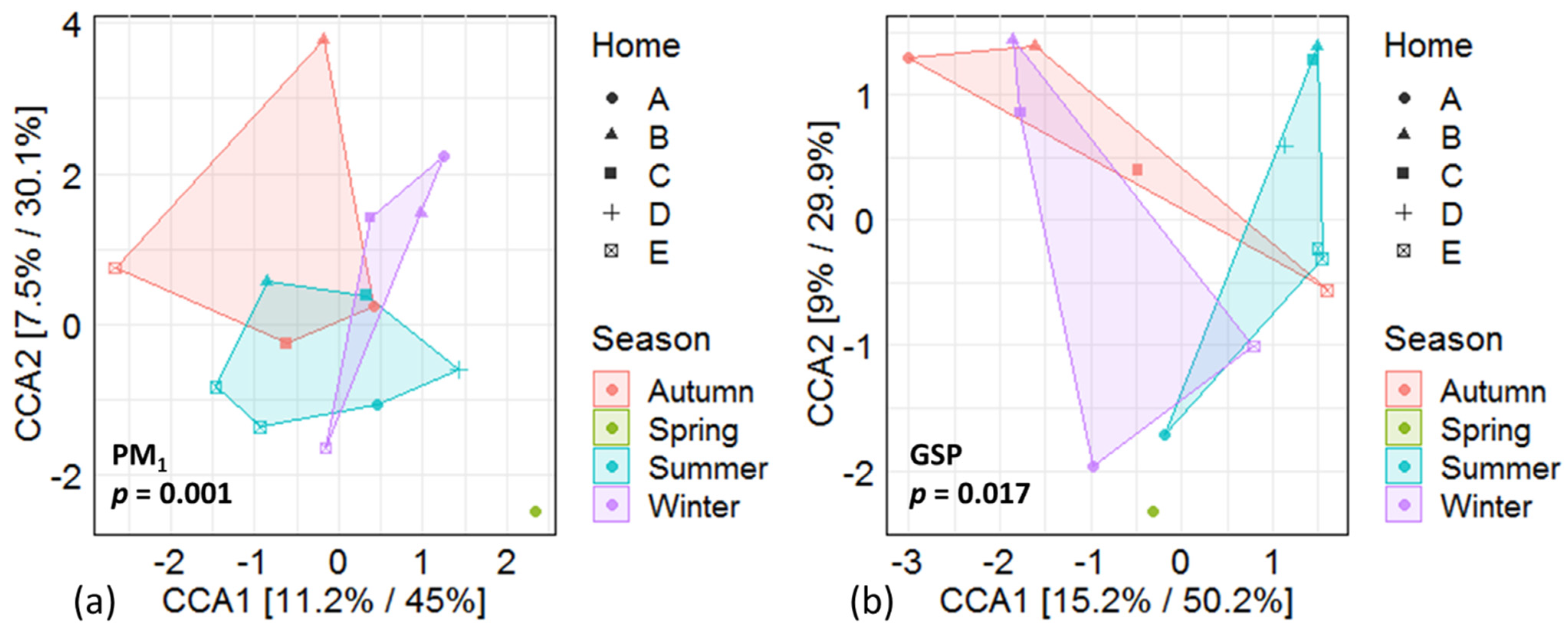
2.4. Data Treatment
3. Results
3.1. Detection of Different Fungi in PM1 and GSP Samples
3.2. PM1/GSP and Correlation between PM1 and GSP Samples—Fungi
3.3. Differences in Microbial Composition between Seasons—Fungi
3.4. Pollen
3.5. Bacteria
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
| SE */Sample | ||
|---|---|---|
| PM1 | GSP | |
| Fungal species | ||
| Acremonium strictum | Bd (7) | 10 |
| Alternaria alternata | Bd (0) | Bd (1) |
| Aspergillus fumigatus | 14 | 14 |
| Aspergillus niger | Bd (2) | 17 |
| Aspergillus versicolor | 9 | 86 |
| Chaetomium globosum | Bd (2) | 14 |
| Cladosporium cladosporioides | 17 | 54 |
| Cladosporium herbarum | 12 | 76 |
| Cladosporium sphaerospermum | 9 | 14 |
| Penicillium chrysogenum | Bd (0) | Bd (0) |
| Penicillium expansum | 12 | 12 |
| Rhizopus stolonifer | Bd (1) | 18 |
| Stachybotrys chartarum | Bd (1) | Bd (2) |
| Tricoderma viride | 11 | 11 |
| Ulocladium chartarum | 8 | 8 |
| Wallemia sebi | 19 | 55 |
| Fungal Group | ||
| Aspergillus glaucus group | Bd (3) | 17 |
| Mucor/Rhizopus group | Bd (13) | 30 |
| Asp/Pen/Pae group | 50 | 54 |
| Universal fungi | 338 | 4300 |
| Pollen | ||
| Betula | Bd (14) | 25 |
| Alnus/Corylus | 15 | 15 |
| Grasses/Artemisia | Bd (7) | 30 |
| Bacterial genus | ||
| Streptomyces spp. | 19 | 50 |
Appendix B
| Organisms | Sample Type Number | GSPa | GSPb | GSPc | GSPd | GSPe | GSPf | GSPg | GSPh | GSPi | GSPj | GSPk | GSPl | PMa | PMb | PMc | PMd | PMe | PMf | PMg | PMh | PMi | PMj | PMk | PMl |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| As. fumigatus | GSPa | 1.00 | |||||||||||||||||||||||
| As. versicolor | GSPb | −0.13 | 1.00 | ||||||||||||||||||||||
| Cl.cladosporioides | GSPc | 0.54 * | 0.19 | 1.00 | |||||||||||||||||||||
| Cl. herbarum | GSPd | 0.58 * | 0.14 | 0.94 ** | 1.00 | ||||||||||||||||||||
| Cl. sphaerospermum | GSPe | 0.67 ** | 0.08 | 0.90 ** | 0.95 ** | 1.00 | |||||||||||||||||||
| Alnus/Corylus | GSPf | 0.39 | 0.19 | 0.15 | 0.05 | 0.19 | 1.00 | ||||||||||||||||||
| Asp/Pen/Paegroup | GSPg | 0.39 | 0.75 ** | 0.49 | 0.45 | 0.42 | 0.36 | 1.00 | |||||||||||||||||
| Pen. expansum | GSPh | −0.23 | 0.69 ** | −0.28 | −0.31 | −0.29 | 0.39 | 0.38 | 1.00 | ||||||||||||||||
| Streptomyces spp. | GSPi | 0.69 ** | 0.18 | 0.60 * | 0.64 ** | 0.75 ** | 0.46 | 0.54 * | 0.05 | 1.00 | |||||||||||||||
| U. chartarum | GSPj | 0.10 | 0.33 | −0.02 | −0.05 | −0.07 | 0.65 ** | 0.48 | 0.54 * | 0.28 | 1.00 | ||||||||||||||
| Universal fungi | GSPk | 0.48 | 0.35 | 0.87 ** | 0.85 ** | 0.78 * | 0.37 | 0.56 * | 0.03 | 0.52 * | 0.19 | 1.00 | |||||||||||||
| W. sebi | GSPl | −0.05 | 0.30 | −0.14 | −0.22 | −0.21 | 0.46 | 0.04 | 0.56* | −0.09 | 0.32 | 0.19 | 1.00 | ||||||||||||
| As. fumigatus | PMa | 0.43 | 0.16 | 0.25 | 0.26 | 0.36 | 0.42 | 0.41 | 0.15 | 0.36 | 0.47 | 0.36 | −0.04 | 1.00 | |||||||||||
| As. versicolor | PMb | 0.56 * | 0.16 | 0.17 | 0.30 | 0.28 | 0.14 | 0.48 | 0.15 | 0.53 * | 0.26 | 0.26 | 0.09 | 0.46 | 1.00 | ||||||||||
| Cl.cladosporioides | PMc | 0.46 | 0.06 | 0.37 | 0.41 | 0.51 | 0.36 | 0.29 | 0.05 | 0.58 * | 0.14 | 0.48 | 0.28 | 0.48 | 0.60 * | 1.00 | |||||||||
| Cl. herbarum | PMd | 0.32 | −0.08 | 0.58 * | 0.61 * | 0.62 * | 0.12 | 0.31 | −0.39 | 0.49 | 0.15 | 0.46 | −0.27 | 0.46 | 0.44 | 0.58 * | 1.00 | ||||||||
| Cl. sphaerospermum | PMe | 0.53 * | 0.29 | 0.56 * | 0.62 * | 0.66 ** | 0.40 | 0.62 * | −0.01 | 0.79 ** | 0.31 | 0.54 * | −0.05 | 0.43 | 0.69 ** | 0.58 * | 0.77 ** | 1.00 | |||||||
| Alnus/Corylus | PMf | 0.54 * | 0.15 | 0.32 | 0.27 | 0.37 | 0.76 ** | 0.51 | 0.16 | 0.71 ** | 0.58 * | 0.37 | 0.17 | 0.56 * | 0.57 * | 0.64 * | 0.57 * | 0.78 ** | 1.00 | ||||||
| Asp/Pen/Paegroup | PMg | 0.55 * | 0.24 | 0.51 | 0.56 * | 0.64 * | 0.38 | 0.56 * | 0.06 | 0.82 ** | 0.30 | 0.45 | −0.07 | 0.52 * | 0.73 ** | 0.62 * | 0.75 ** | 0.95 ** | 0.79 ** | 1.00 | |||||
| Pen. expansum | PMh | 0.21 | 0.45 | 0.09 | −0.01 | 0.08 | 0.42 | 0.49 | 0.38 | 0.22 | 0.39 | 0.23 | 0.17 | 0.81 ** | 0.42 | 0.32 | 0.18 | 0.33 | 0.48 | 0.39 | 1.00 | ||||
| Streptomyces spp. | PMi | 0.02 | 0.54 * | 0.16 | 0.20 | 0.21 | 0.25 | 0.55 * | 0.48 | 0.49 | 0.64 ** | 0.17 | 0.16 | 0.33 | 0.37 | 0.27 | 0.38 | 0.57 * | 0.46 | 0.62 * | 0.26 | 1.00 | |||
| U. chartarum | PMj | 0.12 | 0.33 | 0.08 | 0.11 | 0.07 | −0.03 | 0.41 | −0.06 | 0.24 | 0.13 | −0.05 | −0.23 | 0.33 | 0.48 | 0.29 | 0.42 | 0.52 * | 0.47 | 0.50 | 0.34 | 0.37 | 1.00 | ||
| Universal fungi | PMk | 0.53 * | 0.32 | 0.51 | 0.58 * | 0.60 * | 0.40 | 0.57 * | 0.20 | 0.61 * | 0.31 | 0.65 ** | 0.19 | 0.59 * | 0.71 ** | 0.83 ** | 0.64 * | 0.76 ** | 0.70 ** | 0.79 ** | 0.38 | 0.46 | 0.44 | 1.00 | |
| W. sebi | PMl | −0.32 | 0.28 | 0.10 | 0.08 | 0.06 | 0.22 | 0.15 | 0.08 | 0.21 | 0.24 | 0.13 | 0.02 | −0.22 | −0.35 | 0.04 | 0.06 | 0.12 | 0.10 | −0.05 | −0.14 | 0.23 | 0.04 | −0.13 | 1.00 |
References
- Denning, D.W.; O’Driscoll, B.R.; Hogaboam, C.M.; Bowyer, P.; Niven, R.M. The link between fungi and severe asthma: A summary of the evidence. Eur. Respir. J. 2006, 27, 615–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dellavalle, C.T.; Triche, E.W.; Leaderer, B.P.; Bell, M.L. Effects of ambient pollen concentrations on frequency and severity of asthma symptoms among asthmatic children. Epidemiology 2012, 23, 55–63. [Google Scholar] [CrossRef] [Green Version]
- Green, B.J.; Tovey, E.R.; Sercombe, J.K.; Blachere, F.M.; Beezhold, D.H.; Schmechel, D. Airborne fungal fragments and allergenicity. Med. Mycol. 2006, 44, 245–255. [Google Scholar] [CrossRef] [Green Version]
- Kildesø, J.; Würtz, H.; Nielsen, K.F. Quantification of the release of fungal spores from water damaged plasterboards. In Proceedings of the Danish-Finnish Workshop on Moulds in Buildings, Helsingør, Denmark, 7–8 October 1999. [Google Scholar]
- Afanou, K.A.; Straumfors, A.; Skogstad, A.; Nilsen, T.; Synnes, O.; Skaar, I.; Hjeljord, L.; Tronsmo, A.; Green, B.J.; Eduard, W. Submicronic fungal bioaerosols: High-resolution microscopic characterization and quantification. Appl. Environ. Microbiol. 2014, 80, 7122–7130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madsen, A.M.; Larsen, S.T.; Koponen, I.K.; Kling, K.I.; Barooni, A.; Karottki, D.G.; Tendal, K.; Wolkoff, P. Generation and characterization of indoor fungal aerosols for inhalation studies. Appl. Environ. Microbiol. 2016, 82, 2479–2493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afanou, K.A.; Straumfors, A.; Skogstad, A.; Skaar, I.; Hjeljord, L.; Skare, Ø.; Green, B.J.; Tronsmo, A.; Eduard, W. Profile and morphology of fungal aerosols characterized by field emission scanning electron microscopy (FESEM). Aerosol Sci. Technol. 2015, 49, 423–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, B.J.; Schmechel, D.; Summerbell, R.C. Aerosolized fungal fragments. In Fundamentals of Mold Growth in Indoor Environments and Strategies for Healthy Living; Wageningen Academic Publishers: Wageningen, The Netherlands, 2011; Volume 8, pp. 211–243. [Google Scholar]
- Górny, R.L. Filamentous microorganisms and their fragments in indoor air—A review. Ann. Agric. Environ. Med. 2004, 11, 185–197. [Google Scholar] [PubMed]
- Madsen, A.M.; Schlünssen, V.; Olsen, T.; Sigsgaard, T.; Avci, H. Airborne fungal and bacterial components in PM1 dust from biofuel plants. Ann. Occup. Hyg. 2009, 53, 749–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tendal, K.; Madsen, A.M. Exposure to airborne microorganisms, hyphal fragments, and pollen in a field of organically grown strawberries. Aerobiologia 2011, 27, 13–23. [Google Scholar] [CrossRef]
- Lee, S.A.; Liao, C.H. Size-selective assessment of agricultural workers’ personal exposure to airborne fungi and fungal fragments. Sci. Total Environ. 2014, 466–467, 725–732. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, A.; Reponen, T.; Rylander, R. Airborne fungal cell fragments in homes in relation to total fungal biomass. Indoor Air 2013, 23, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Singh, U.; Reponen, T.; Cho, K.J.; Grinshpun, S.A.; Adhikari, A.; Levin, L.; Indugula, R.; Green, B.J. Airborne endotoxin and β-D-glucan in PM1 in agricultural and home environments. Aerosol Air Qual. Res. 2011, 11, 376–386. [Google Scholar] [CrossRef]
- Reponen, T.; Seo, S.C.; Grimsley, F.; Lee, T.; Crawford, C.; Grinshpun, S.A. Fungal fragments in moldy houses: A field study in homes in New Orleans and Southern Ohio. Atmos. Environ. 2007, 41, 8140–8149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gou, H.; Lu, J.; Li, S.; Tong, Y.; Xie, C.; Zheng, X. Assessment of microbial communities in PM1 and PM10 of Urumqi during winter. Environ. Pollut. 2016, 214, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Wei, M.; Chen, J.; Zhu, C.; Li, J.; Lv, G.; Xu, X.; Zheng, L.; Sui, G.; Li, W.; et al. Fungi diversity in PM2.5 and PM1 at the summit of Mt. Tai: Abundance, size distribution, and seasonal variation. Atmos. Chem. Phys. 2017, 17, 11247–11260. [Google Scholar] [CrossRef] [Green Version]
- Cho, S.H.; Seo, S.C.; Schmechel, D.; Grinshpun, S.A.; Reponen, T. Aerodynamic characteristics and respiratory deposition of fungal fragments. Atmos. Environ. 2005, 39, 5454–5465. [Google Scholar] [CrossRef]
- Frankel, M.; Hansen, E.W.; Madsen, A.M. Effect of relative humidity on the aerosolization and total inflammatory potential of fungal particles from dust-inoculated gypsum boards. Indoor Air 2014, 24, 16–28. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Hwang, G.B.; Jung, J.H.; Lee, D.H.; Lee, B.U. Generation characteristics of fungal spore and fragment bioaerosols by airflow control over fungal cultures. J. Aerosol Sci. 2010, 41, 319–325. [Google Scholar] [CrossRef]
- Seo, S.C.; Choung, J.T.; Cehn, B.T.; Lindsley, W.G.; Kim, K.Y. The level of submicron fungal fragments in homes with asthmatic children. Environ. Res. 2014, 131, 71–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paulskn, E.; Søgaard, J.; Anderskn, K.E. Occupational dermatitis in Danish gardeners and greenhouse workers (III). Compositae-related symptoms. Contact Dermat. 1998, 38, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Suphioglu, C.; Singh, M.B.; Taylor, P.; Knox, R.B.; Bellomo, R.; Holmes, P.; Puy, R. Mechanism of grass-pollen-induced asthma. Lancet 1992, 339, 569–572. [Google Scholar] [CrossRef]
- Davies, J.M. Grass pollen allergens globally: The contribution of subtropical grasses to burden of allergic respiratory diseases. Clin. Exp. Allergy 2014, 44, 790–801. [Google Scholar] [CrossRef] [PubMed]
- Taylor, P.E.; Flagan, R.C.; Valenta, R.; Glovsky, M.M. Release of allergens as respirable aerosols: A link between grass pollen and asthma. J. Allergy Clin. Immunol. 2002, 109, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Spieksma, F.T.M.; Kramps, J.A.; Van der Linden, A.C.; Nikkels, B.H.; Plomp, A.; Koerten, H.K.; Dijkman, J.H. Evidence of grass-pollen allergenic activity in the smaller micronic atmospheric aerosol fraction. Clin. Exp. Allergy 1990, 20, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Hughes, D.D.; Mampage, C.B.A.; Jones, L.M.; Liu, Z.; Stone, E.A. Characterization of Atmospheric Pollen Fragments during Springtime Thunderstorms. Environ. Sci. Technol. Lett. 2020, 7, 409–414. [Google Scholar] [CrossRef]
- Mensah-Attipoe, J.; Saari, S.; Veijalainen, A.M.; Pasanen, P.; Keskinen, J.; Leskinen, J.T.T.; Reponen, T. Release and characteristics of fungal fragments in various conditions. Sci. Total Environ. 2016, 547, 234–243. [Google Scholar] [CrossRef] [PubMed]
- Górny, R.L.; Reponen, T.; Willeke, K.; Schmechel, D.; Robine, E.; Boissier, M.; Grinshpun, S.A. Fungal fragments as indoor air biocontaminants. Appl. Environ. Microbiol. 2002, 68, 3522–3531. [Google Scholar] [CrossRef] [Green Version]
- Kildesø, J.; Würtz, H.; Nielsen, K.F.; Kruse, P.; Wilkins, K.; Thrane, U.; Gravesen, S.; Nielsen, P.A.; Schneider, T. Determination of fungal spore release from wet building materials. Indoor Air 2003, 13, 148–155. [Google Scholar] [CrossRef]
- Madsen, A.M. Effects of Airflow and Changing Humidity on the Aerosolization of Respirable Fungal Fragments and Conidia of Botrytis cinerea. Appl. Environ. Microbiol. 2012, 78, 3999–4007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afanou, K.A.; Straumfors, A.; Skogstad, A.; Nayak, A.P.; Skaar, I.; Hjeljord, L.; Tronsmo, A.; Eduard, W.; Green, B.J. Indirect immunodetection of fungal fragments by field emission scanning electron microscopy. Appl. Environ. Microbiol. 2015, 81, 5794–5803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johansson, E.; Reponen, T.; Meller, J.; Vesper, S.; Yadav, J. Association of Streptomyces community composition determined by PCR-denaturing gradient gel electrophoresis with indoor mold status. Environ. Monit. Assess. 2014, 186, 8773–8783. [Google Scholar] [CrossRef] [PubMed]
- Mazza, P.; Noens, E.E.; Schirner, K.; Grantcharova, N.; Mommaas, A.M.; Koerten, H.K.; Muth, G.; Flärdh, K.; Van Wezel, G.P.; Wohlleben, W. MreB of Streptomyces coelicolor is not essential for vegetative growth but is required for the integrity of aerial hyphae and spores. Mol. Microbiol. 2006, 60, 838–852. [Google Scholar] [CrossRef] [PubMed]
- Jussila, J.; Pelkonen, J.; Kosma, V.M.; Mäki-Paakkanen, J.; Komulainen, H.; Hirvonen, M.R. Systemic immunoresponses in mice after repeated exposure of lungs to spores of Streptomyces californicus. Clin. Diagn. Lab. Immunol. 2003, 10, 30–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kagen, S.L.; Fink, J.N.; Schlueter, D.P.; Kurup, V.P.; Fruchtman, R.B. Streptomyces albus: A new cause of hypersensitivity pneumonitis. J. Allergy Clin. Immunol. 1981, 68, 295–299. [Google Scholar] [CrossRef]
- Frankel, M.; Bekö, G.; Timm, M.; Gustavsen, S.; Hansen, E.W.; Madsen, A.M. Seasonal variations of indoor microbial exposures and their relation to temperature, relative humidity, and air exchange rate. Appl. Environ. Microbiol. 2012, 78, 8289–8297. [Google Scholar] [CrossRef] [Green Version]
- Kenny, L.C.; Aitken, R.J.; Baldwin, P.E.J.; Beaumont, G.C.; Maynard, A.D. The sampling efficiency of personal inhalable aerosol samplers in low air movement environments. J. Aerosol Sci. 1999, 30, 627–638. [Google Scholar] [CrossRef]
- Kenny, L.C.; Gussman, R.A. A direct approach to the design of cyclones for aerosol-monitoring applications. J. Aerosol Sci. 2000, 31, 1407–1420. [Google Scholar] [CrossRef]
- Choi, S.-J. The Effect of Outdoor Air and Indoor Human Activity on Mass Concentrations of Size-Selective Particulate in Classrooms. Korean J. Environ. Heal. Sci. 2008, 34, 137–147. [Google Scholar] [CrossRef] [Green Version]
- Green, B.J.; Schmechel, D.; Tovey, E.R. Detection of Aerosolized alternaria alternata conidia, hyphae, and fragments by using a novel double-immunostaining technique. Clin. Diagn. Lab. Immunol. 2005, 12, 1114–1116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanaani, H.; Hargreaves, M.; Ristovski, Z.; Morawska, L. Fungal spore fragmentation as a function of airflow rates and fungal generation methods. Atmos. Environ. 2009, 43, 3725–3735. [Google Scholar] [CrossRef] [Green Version]
- Górny, R.L.; Ławniczek-Wałczyk, A. Effect of two aerosolization methods on the release of fungal propagules from a contaminated agar surface. Ann. Agric. Environ. Med. 2012, 19, 279. [Google Scholar] [PubMed]
- Madsen, A.M.; Kruse, P.; Schneider, T. Characterization of microbial particle release from biomass and building material surfaces for inhalation exposure risk assessment. Ann. Occup. Hyg. 2006, 50, 175–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afanou, A.K.; Straumfors, A.; Eduard, W. Fungal aerosol composition in moldy basements. Indoor Air 2019, 29, 780–790. [Google Scholar] [CrossRef] [PubMed]
- Li, D.-W.; Kendrick, B. A year-round comparison of fungal spores in indoor and outdoor air. Mycologia 1995, 29, 190–195. [Google Scholar] [CrossRef]
- Hammed, A.A.A.; Khodr, M.I. Suspended particulates and bioaerosols emitted from an agricultural non-point source. J. Environ. Monit. 2001, 3, 206–209. [Google Scholar] [CrossRef]
- Liu, Z.; Li, A.; Hu, Z.; Sun, H. Study on the potential relationships between indoor culturable fungi, particle load and children respiratory health in Xi’an, China. Build. Environ. 2014, 80, 105–114. [Google Scholar] [CrossRef]
- Kim, K.Y.; Kim, C.N. Airborne microbiological characteristics in public buildings of Korea. Build. Environ. 2007, 42, 2188–2196. [Google Scholar] [CrossRef]
- White, J.K.; Nielsen, J.L.; Madsen, A.M. Potential Respiratory Deposition and Species Composition of Airborne Culturable, Viable, and Non-Viable Fungi during Occupancy in a Pig Farm. Atmosphere 2020, 11, 639. [Google Scholar] [CrossRef]
- Knudsen, S.M.; Gunnarsen, L.; Madsen, A.M. Airborne fungal species associated with mouldy and non-mouldy buildings—Effects of air change rates, humidity, and air velocity. Build. Environ. 2017, 122, 161–170. [Google Scholar] [CrossRef]
- Zeng, Q.Y.; Westermark, S.O.; Rasmuson-Lestander, Å.; Wang, X.R. Detection and quantification of Cladosporium in aerosols by real-time PCR. J. Environ. Monit. 2006, 8, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.Y.; Westermark, S.O.; Rasmuson-Lestander, Å.; Wang, X.R. Detection and quantification of Wallemia sebi in aerosols by real-time PCR, conventional PCR, and cultivation. Appl. Environ. Microbiol. 2004, 70, 7295–7302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Betelli, L.; Duquenne, P.; Grenouillet, F.; Simon, X.; Scherer, E.; Géhin, E.; Hartmann, A. Development and evaluation of a method for the quantification of airborne Thermoactinomyces vulgaris by real-time PCR. J. Microbiol. Methods 2013, 92, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Pietarinen, V.M.; Rintala, H.; Hyvärinen, A.; Lignell, U.; Kärkkäinen, P.; Nevalainen, A. Quantitative PCR analysis of fungi and bacteria in building materials and comparison to culture-based analysis. J. Environ. Monit. 2008, 10, 655–663. [Google Scholar] [CrossRef]
- Brandl, H. Bioaerosols in Indoor Environment—A Review with Special Reference to Residential and Occupational Locations. Open Environ. Biol. Monit. J. 2011, 4, 83–96. [Google Scholar] [CrossRef] [Green Version]
- Adams, R.I.; Miletto, M.; Taylor, J.W.; Bruns, T.D. Dispersal in microbes: Fungi in indoor air are dominated by outdoor air and show dispersal limitation at short distances. ISME J. 2013, 7, 1262–1273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrett, M.H.; Hooper, B.M.; Cole, F.M.; Hooper, M.A. Airborne fungal spores in 80 homes in the Latrobe Valley, Australia: Levels, seasonality and indoor-outdoor relationship. Aerobiologia 1997, 13, 121–126. [Google Scholar] [CrossRef]
- Grinn-Gofroń, A.; Rapiejko, P. Occurrence of Cladosporium spp. and Alternaria spp. spores in Western, Northern and Central-Eastern Poland in 2004–2006 and relation to some meteorological factors. Atmos. Res. 2009, 93, 747–758. [Google Scholar] [CrossRef]
- Kaarakainen, P.; Rintala, H.; Vepsäläinen, A.; Hyvärinen, A.; Nevalainen, A.; Meklin, T. Microbial content of house dust samples determined with qPCR. Sci. Total Environ. 2009, 407, 4673–4680. [Google Scholar] [CrossRef]
- Reboux, G.; Bellanger, A.P.; Roussel, S.; Grenouillet, F.; Sornin, S.; Piarroux, R.; Dalphin, J.C.; Millon, L. Indoor mold concentration in Eastern France. Indoor Air 2009, 19, 446–453. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.; Tendal, K.; Frederiksen, M.W.; Uhrbrand, K.; Li, Y.; Madsen, A.M. Strong variance in the inflammatory and cytotoxic potentials of Penicillium and Aspergillus species from cleaning workers’ exposure in nursing homes. Sci. Total Environ. 2020, 724, 138231. [Google Scholar] [CrossRef]
- Reboux, G.; Rocchi, S.; Vacheyrou, M.; Millon, L. Identifying indoor air Penicillium species: A challenge for allergic Patients. J. Med. Microbiol. 2019, 68, 812–821. [Google Scholar] [CrossRef] [PubMed]
- Lewis, C.W.; Smith, J.E.; Anderson, J.G.; Murad, Y.M. The Presence of Mycotoxin-Associated Fungal Spores Isolated from the Indoor Air of the Damp Domestic Environment and Cytotoxic to Human Cell Lines. Indoor Built Environ. 1994, 3, 323–330. [Google Scholar] [CrossRef]
- D’Amato, G.; Cecchi, L.; Bonini, S.; Nunes, C.; Annesi-Maesano, I.; Behrendt, H.; Liccardi, G.; Popov, T.; Van Cauwenberge, P. Allergenic pollen and pollen allergy in Europe. Allergy Eur. J. Allergy Clin. Immunol. 2007, 62, 976–990. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, A. The effects of climate change on the birch pollen season in Denmark. Aerobiologia 2002, 18, 253–265. [Google Scholar] [CrossRef]
- Yli-Panula, E.; Fekedulegn, D.B.; Green, B.J.; Ranta, H. Analysis of airborne Betula pollen in Finland; a 31-year perspective. Int. J. Environ. Res. Public Health 2009, 6, 1706–1723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peternel, R.; Srnec, L.; Hrga, I.; Hercog, P.; Čulig, J. Airborne pollen of Betula, Corylus and Alnus in Zagreb, Croatia. A three-year record. Grana 2005, 44, 187–191. [Google Scholar] [CrossRef]
- Stach, A.; García-Mozo, H.; Prieto-Baena, J.C.; Czarnecka-Operacz, M.; Jenerowicz, D.; Silny, W.; Galán, C. Prevalence of Artemisia species pollinosis in western Poland: Impact of climate change on aerobiological trends, 1995-2004. J. Investig. Allergol. Clin. Immunol. 2007, 17, 39–47. [Google Scholar] [PubMed]
- Oteros, J.; Bartusel, E.; Alessandrini, F.; Núñez, A.; Moreno, D.A.; Behrendt, H.; Schmidt-Weber, C.; Traidl-Hoffmann, C.; Buters, J. Artemisia pollen is the main vector for airborne endotoxin. J. Allergy Clin. Immunol. 2019, 143, 369–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grote, M.; Vrtala, S.; Niederberger, V.; Wiermann, R.; Valenta, R.; Reichelt, R. Release of allergen-bearing cytoplasm from hydrated pollen: A mechanism common to a variety of grass (poaceae) species revealed by electron microscopy. J. Allergy Clin. Immunol. 2001, 108, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Taylor, P.E.; Jacobson, K.W.; House, J.M.; Glovsky, M.M. Links between pollen, atopy and the asthma epidemic. Int. Arch. Allergy Immunol. 2007, 144, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Spieksma, F.T.M.; Nikkels, B.H.; Dijkman, J.H. Seasonal appearance of grass pollen allergen in natural, pauci-micronic aerosol of various size fractions. Relationship with airborne grass pollen concentration. Clin. Exp. Allergy 1995, 25, 234–239. [Google Scholar] [CrossRef] [PubMed]
- Losappio, L.; Heffler, E.; Contento, F.; Cannito, C.; Rolla, G. Thunderstorm-related asthma epidemic owing to Olea Europaea pollen sensitization. Allergy Eur. J. Allergy Clin. Immunol. 2011, 66, 1510–1511. [Google Scholar] [CrossRef]
- Taylor, P.; Goldberg, C.; Buch, H.; Moseholm, L.; Weeke, E.R. Airborne Pollen Records in Denmark, 1977–1986. Grana 1988, 27, 209–217. [Google Scholar] [CrossRef]
- Lumpkins, E.D.; Corbit, S.L.; Tiedeman, G.M. Airborne fungi survey. I. Culture plate survey of the home environment. Ann. Allergy 1973, 31, 361–370. [Google Scholar] [PubMed]
- Grinshpun, S.A.; Reponen, T.; Willeke, K. Aerosol characteristics of airborne actinomycetes and fungi. J. Aerosol Sci. 1997, 28, 667–668. [Google Scholar] [CrossRef]
- Reponen, T.; Grinshpun, S.A.; Conwell, K.L.; Wiest, J.; Anderson, M. Aerodynamic versus physical size of spores: Measurement and implication for respiratory deposition. Grana 2001, 40, 119–125. [Google Scholar] [CrossRef]

| This Study | Fragments Found in Previous Studies | |||||||
|---|---|---|---|---|---|---|---|---|
| Number of Positive Samples out of 15 Samples | PM1/GSP (GM, %) | |||||||
| PM1 | GSP | PM1/GSP | Fragment/Spore | Place of Sampling | Method of Quantification | Reference | ||
| Fungal species | ||||||||
| Acremonium strictum | Bd | 8 | Bd | - | - | - | - | |
| Alternaria alternata | Bd | Bd | Bd | NM | Laboratory c | No. by microscopy | [41] | |
| Aspergillus fumigatusb | 1 | 14 | 0.37% | ~1% | Laboratory | No. by microscopy | [7] | |
| Aspergillus niger | Bd | 2 | Bd | 0–19.2% | Laboratory | No. particles | [42] | |
| Aspergillus versicolorb | 6 | 15 | 0.59% | 100% | Laboratory | No. by microscopy | [18] | |
| <30% | Laboratory | No. of particles | [30] | |||||
| ~2% | Laboratory | No. by microscopy | [7] | |||||
| 10% | Laboratory | No. of particles | [28] | |||||
| ~50% | Laboratory | No. of particles | [43] | |||||
| Chaetomium globosum | Bd | 2 | Bd | - | - | - | - | |
| Cladosporium cladosporioidesb | 5 | 13 | 0.71% | 0–22.4% | Laboratory | No. of particles | [42] | |
| 2% | Laboratory | No. of particles | [28] | |||||
| ~50% | Laboratory | No. of particles | [43] | |||||
| Cladosporium herbaruma, b | 9 | 15 | 0.63% | - | - | |||
| Cladosporium sphaerospermuma, b | 2 | 15 | 1.08% | - | - | |||
| Penicillium chrysogenum | Bd | Bd | Bd | ~0.01% | Laboratory | No. of particles | [44] | |
| ~2% | Laboratory | No. by microscopy | [7] | |||||
| ~40% | Laboratory | No. of particles | [43] | |||||
| Penicillium expansumb | 1 | 8 | 16.47% | - | - | - | - | |
| Rhizopus stolonifer | Bd | 1 | Bd | - | - | - | - | |
| Stachybotrys chartarum | Bd | Bd | Bd | 51,400% | Laboratory | No. by microscopy | [18] | |
| Tricoderma viride | Bd | 4 | Bd | - | - | - | ||
| Ulocladium chartarumb | 1 | 2 | 39.64% | ~15% | Laboratory | No. of particles | [30] | |
| Wallemia sebib | 2 | 11 | 1.16% | - | - | - | - | |
| Fungal Groups | ||||||||
| Aspergillus glaucus group | Bd | 6 | Bd | - | - | - | - | |
| Mucor/Rhizopus group | Bd | 14 | Bd | - | - | - | - | |
| Asp/Pen/Pae group a, b | 12 | 15 | 4.68% | - | - | |||
| Universal fungia, b | 15 | 15 | 2.95% | ~35% | Open field | No. by microscopy | [11] | |
| <1% | Laboratory gypsum | No. particles | [19] | |||||
| ~5% | Laboratory gypsum | (1→3)-β-D-glucan | [19] | |||||
| 0.3–1.3% | Laboratory gypsum | No. particles | [6] | |||||
| 170% | Basement | No. by microscopy | [45] | |||||
| 1–216% | Homes | (1→3)-β-D-glucan | [15] | |||||
| 6.3% | Home | No. by microscopy | [46] | |||||
| 0.08–19.5% | Farms | (1→3)-β-D-glucan | [12] | |||||
| Pollen | ||||||||
| Betula | Bd | 4 | Bd | - | - | - | - | |
| Alnus/Corylusa, b | 4 | 10 | 16.70% | - | - | - | - | |
| Grasses/Artemisia | Bd | 6 | Bd | <62% | Outdoor | No. by microscopy | [26] | |
| Bacterial genus | ||||||||
| Streptomyces spp. b | 9 | 15 | 3.66% | - | Field | Cultivable | [47] | |
| PM1 | Season | Spring | Summer | Autumn | Winter | ||||
|---|---|---|---|---|---|---|---|---|---|
| p-Value | β | p-Value | β | p-Value | β | p-Value | β | p-Value | |
| Fungal species | |||||||||
| Aspergillus fumigatus | <0.0001 | 1.94 | <0.0001 | 0.32 | 0.05 | 0.103 | 0.54 | Ref. | - |
| Aspergillus versicolor | 0.0757 | 2.74 | 0.050 | 0.40 | 0.58 | −0.91 | 0.28 | Ref. | - |
| Cladosporiumcladosporioides | 0.25 | 1.44 | 0.11 | 0.51 | 0.30 | −0.088 | 0.87 | Ref. | - |
| Cladosporium herbarum | 0.095 | 2.28 | 0.07 | 1.76 | 0.027 | 0.91 | 0.24 | Ref. | - |
| Cladosporium sphaerospermum | 0.17 | 1.40 | 0.047 | 0.61 | 0.12 | 0.38 | 0.35 | Ref. | - |
| Penicillium expansum | 0.0019 | 2.94 | 0.0002 | 0.26 | 0.43 | 0.32 | 0.39 | Ref. | - |
| Ulocladium chartarum | 0.76 | 0.50 | 0.36 | 0.08 | 0.79 | −0.06 | 0.85 | Ref. | - |
| Wallemia sebi | 0.23 | 0.078 | 0.91 | 0.12 | 0.74 | 0.79 | 0.075 | Ref. | - |
| Fungal Group | |||||||||
| Asp/Pen/Pae group | 0.15 | 1.58 | 0.037 | 0.56 | 0.17 | 0.28 | 0.52 | Ref. | |
| Universal fungi | 0.12 | 0.74 | 0.030 | 0.22 | 0.22 | 0.02 | 0.91 | Ref. | - |
| Pollen | |||||||||
| Alnus/Corylus | 0.26 | 1.33 | 0.058 | 0.255 | 0.49 | 0.36 | 0.39 | Ref. | - |
| Bacterial genus | |||||||||
| Streptomyces spp. | 0.0041 | 2.47 | 0.0035 | −0.21 | 0.52 | 1.10 | 0.015 | Ref. | - |
| GSP | Season | Spring | Summer | Autumn | Winter | ||||
|---|---|---|---|---|---|---|---|---|---|
| p-Value | β | p-Value | β | p-Value | β | p-Value | β | p-Value | |
| Fungal species | |||||||||
| Acremonium strictum | 0.13 | 1.74 | 0.065 | 0.98 | 0.071 | 0.24 | 0.66 | Ref. | - |
| Aspergillus fumigatus | 0.057 | 0.62 | 0.22 | 0.64 | 0.037 | −0.11 | 0.69 | Ref. | - |
| Aspergillus niger | 0.0057 | 1.77 | 0.0011 | 0.67 | 0.0087 | 0.47 | 0.05 | Ref. | - |
| Aspergillus versicolor | 0.20 | 0.77 | 0.10 | 0.017 | 0.95 | 0.39 | 0.17 | Ref. | - |
| Chaetomium globosum | 0.56 | −0.18 | 0.78 | −0.05 | 0.90 | −0.51 | 0.22 | Ref. | - |
| Cladosporiumcladosporioides | 0.0002 | 1.44 | 0.016 | 2.12 | <0.0001 | 1.37 | 0.0013 | Ref. | - |
| Cladosporium herbarum | 0.0025 | 1.05 | 0.061 | 1.501 | 0.0003 | 0.66 | 0.06 | Ref. | - |
| Cladosporium sphaerospermum | 0.0085 | 1.28 | 0.042 | 1.36 | 0.0014 | 0.57 | 0.13 | Ref. | - |
| Penicillium expansum | 0.014 | 0.95 | 0.15 | −0.93 | 0.02 | 0.14 | 0.73 | Ref. | - |
| Rhizopus stolonifera | 0.42 | 0.82 | 0.20 | −0.17 | 0.61 | −0.12 | 0.74 | Ref. | - |
| Tricoderma viride | 0.29 | −0.17 | 0.53 | −0.29 | 0.07 | −0.11 | 0.48 | Ref. | - |
| Ulocladium chartarum | 0.0048 | 1.39 | 0.008 | −0.19 | 0.41 | 0.74 | 0.016 | Ref. | - |
| Wallemia sebi | 0.13 | 0.011 | 0.99 | −0.58 | 0.22 | 0.59 | 0.23 | Ref. | - |
| Fungal Groups | |||||||||
| Aspergillus glaucus group | 0.44 | 0.037 | 0.91 | 0.0004 | 0.99 | 0.27 | 0.18 | Ref. | - |
| Mucor/Rhizopus group | 0.0003 | 1.78 | 0.0005 | 1.29 | <0.0001 | 0.64 | 0.018 | Ref. | - |
| Asp/Pen/Pae group | 0.062 | 1.27 | 0.01 | 0.34 | 0.17 | 0.44 | 0.11 | Ref. | - |
| Universal fungi | 0.018 | 0.86 | 0.029 | 0.77 | 0.004 | 0.55 | 0.03 | Ref. | - |
| Pollen | |||||||||
| Betula | 0.056 | 1.05 | 0.059 | 0.48 | 0.11 | 0.87 | 0.015 | Ref. | - |
| Alnus/Corylus | 0.23 | 1.58 | 0.15 | −0.061 | 0.92 | 0.89 | 0.18 | Ref. | - |
| Grasses/Artemisia | 0.13 | 2.91 | 0.02 | 0.72 | 0.29 | 0.59 | 0.41 | Ref. | - |
| Bacterial genus | |||||||||
| Streptomyces spp. | 0.27 | 1.05 | 0.096 | 0.55 | 0.13 | 0.31 | 0.42 | Ref. | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, R.; Pørneki, A.D.; Lindgreen, J.N.; Li, Y.; Madsen, A.M. Species of Fungi and Pollen in the PM1 and the Inhalable Fraction of Indoor Air in Homes. Atmosphere 2021, 12, 404. https://doi.org/10.3390/atmos12030404
Lu R, Pørneki AD, Lindgreen JN, Li Y, Madsen AM. Species of Fungi and Pollen in the PM1 and the Inhalable Fraction of Indoor Air in Homes. Atmosphere. 2021; 12(3):404. https://doi.org/10.3390/atmos12030404
Chicago/Turabian StyleLu, Rui, Ann Dorte Pørneki, Jonas N. Lindgreen, Yanpeng Li, and Anne Mette Madsen. 2021. "Species of Fungi and Pollen in the PM1 and the Inhalable Fraction of Indoor Air in Homes" Atmosphere 12, no. 3: 404. https://doi.org/10.3390/atmos12030404
APA StyleLu, R., Pørneki, A. D., Lindgreen, J. N., Li, Y., & Madsen, A. M. (2021). Species of Fungi and Pollen in the PM1 and the Inhalable Fraction of Indoor Air in Homes. Atmosphere, 12(3), 404. https://doi.org/10.3390/atmos12030404