
Elevation-Dependent Changes to Plant Phenology in Canada’s Arctic Detected Using Long-Term Satellite Observations


Abstract
:1. Introduction
2. Materials and Methods
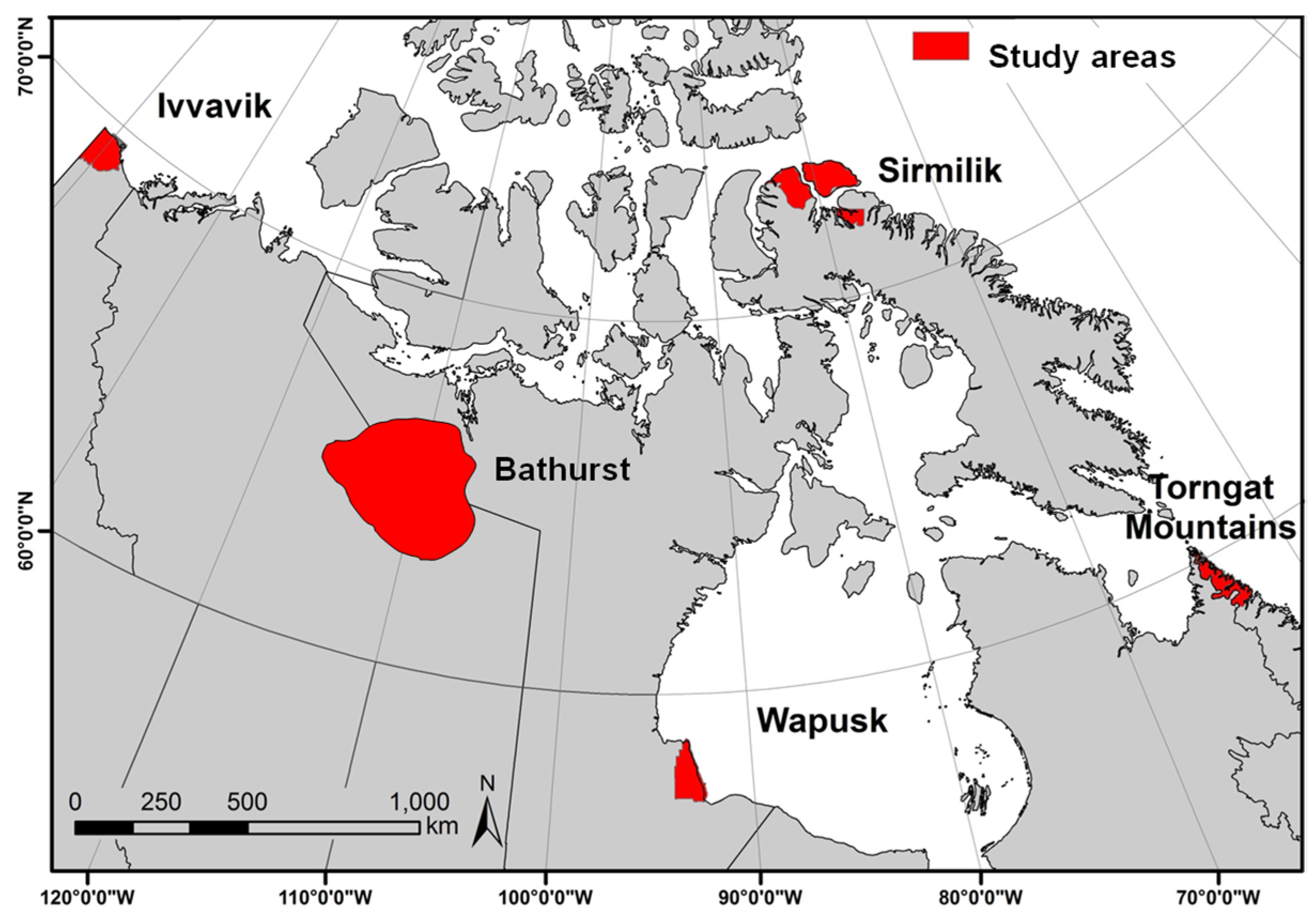
2.1. Study Areas
2.2. Data Sources
2.3. Methods
2.3.1. Determining the Dominant Land Cover Class at 1-km Resolution from Landsat-Based Maps
2.3.2. Constructing Seasonal Profile of Vegetation Index for a Land Cover Class in a Study Area
2.3.3. Estimating Seasonal Variations in Leaf Biomass for a Land Cover Class
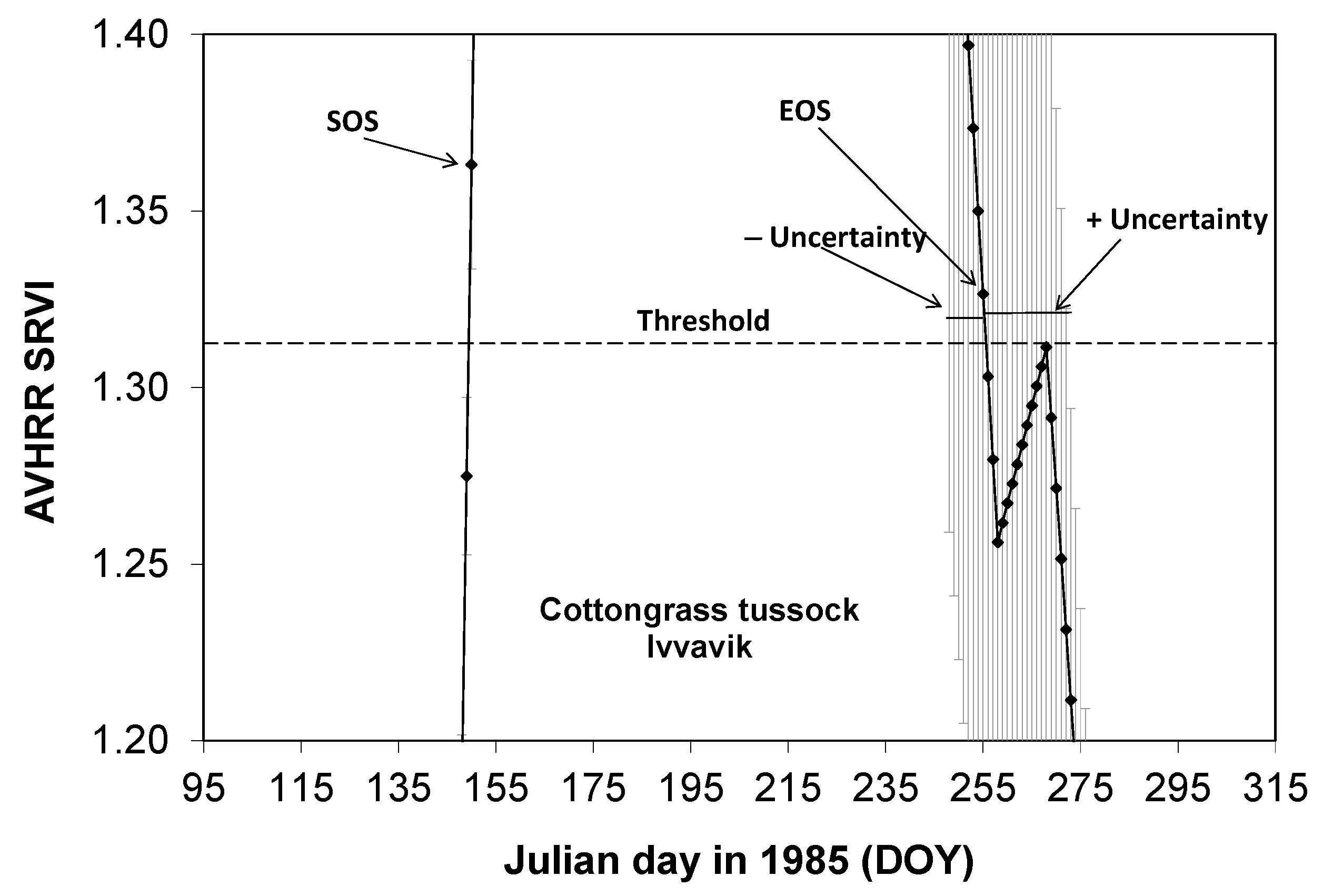
2.3.4. Detecting SOS, EOS, and GSL for a Land Cover Class in a Study Area
2.3.5. Detecting SOS, EOS, and GSL for a Land Cover Class in a Study Area
3. Results
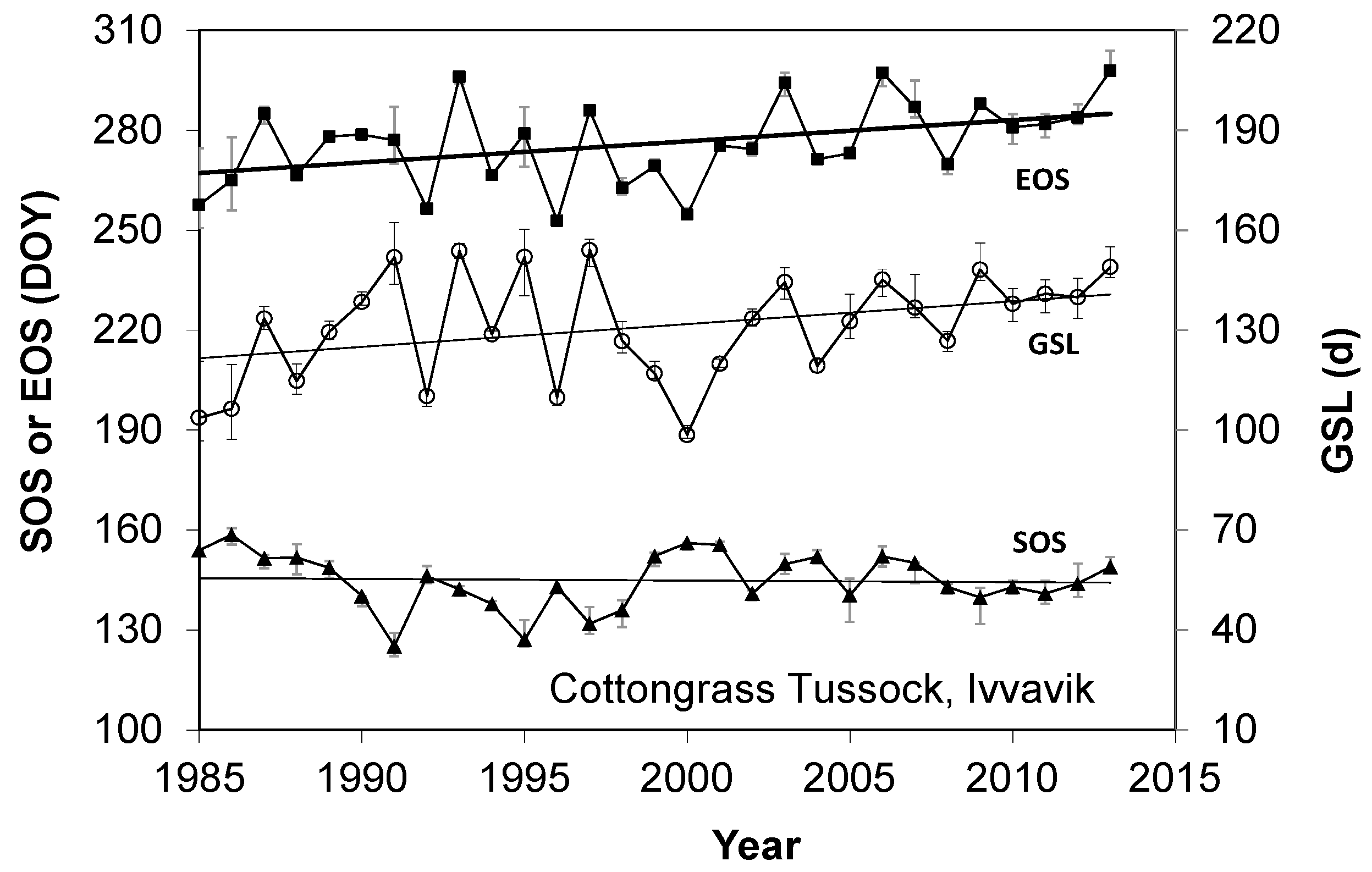
3.1. Long Terms Trends in SOS, EOS, and GSL during 1985–2013 over the Five Study Areas
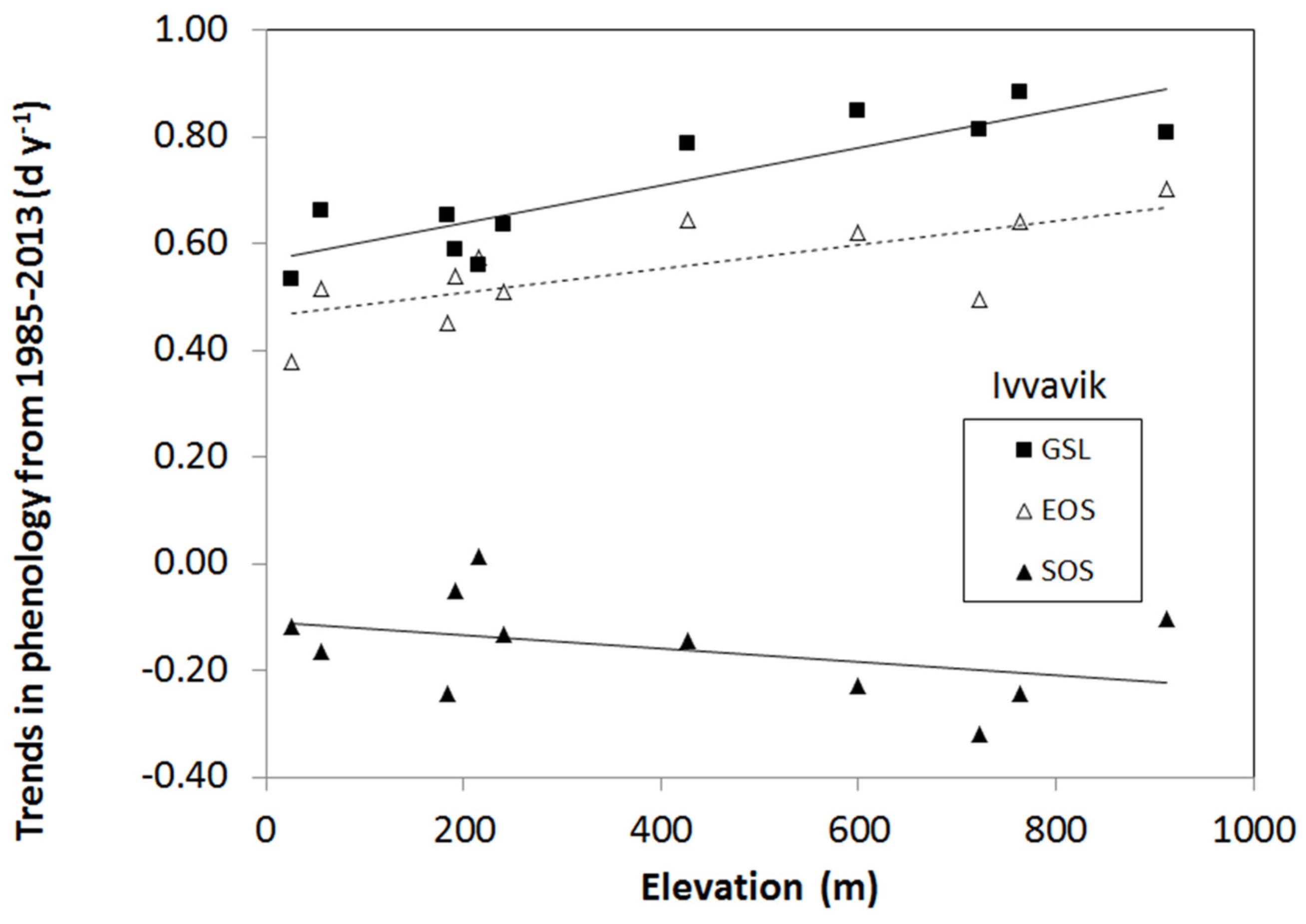
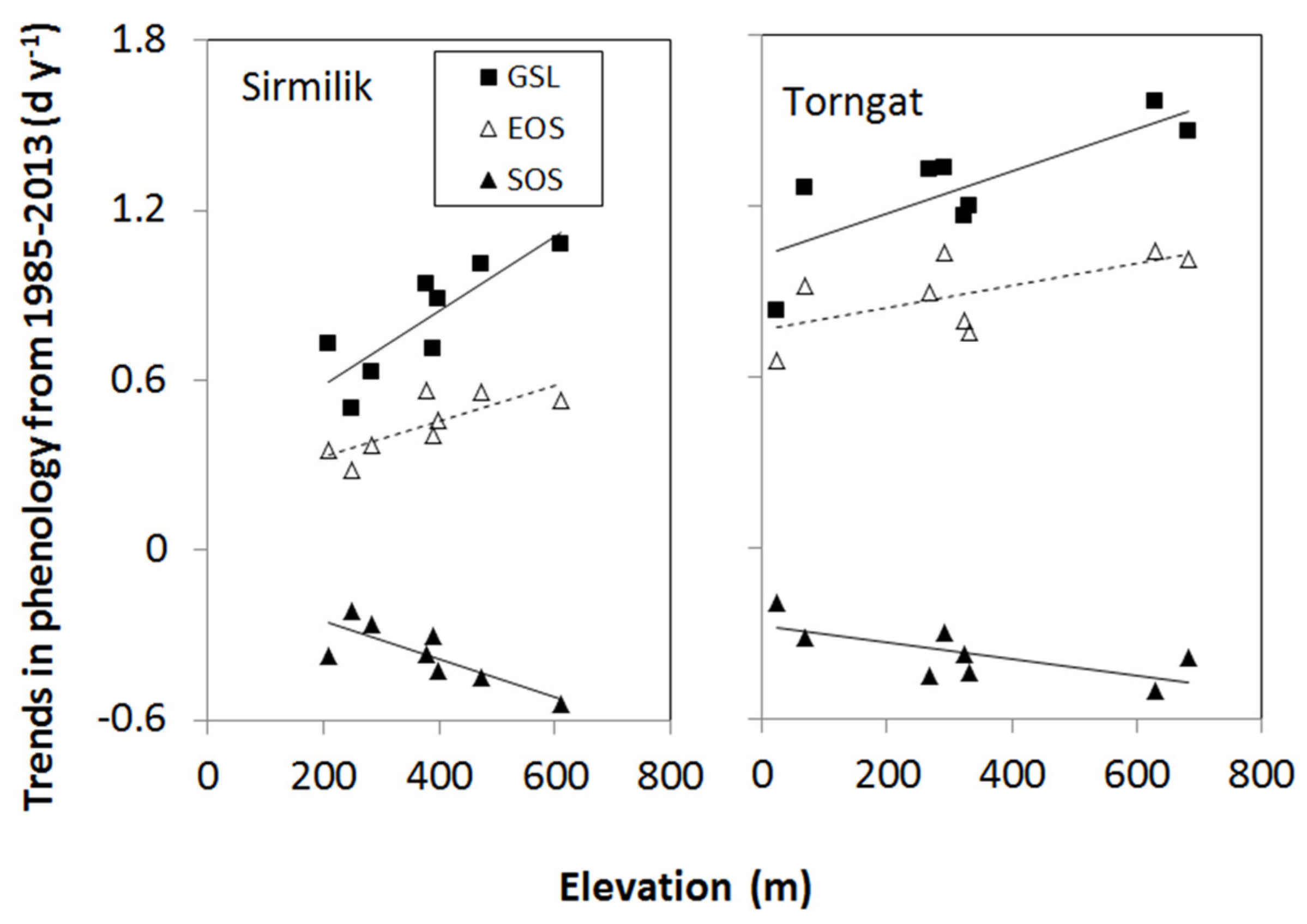
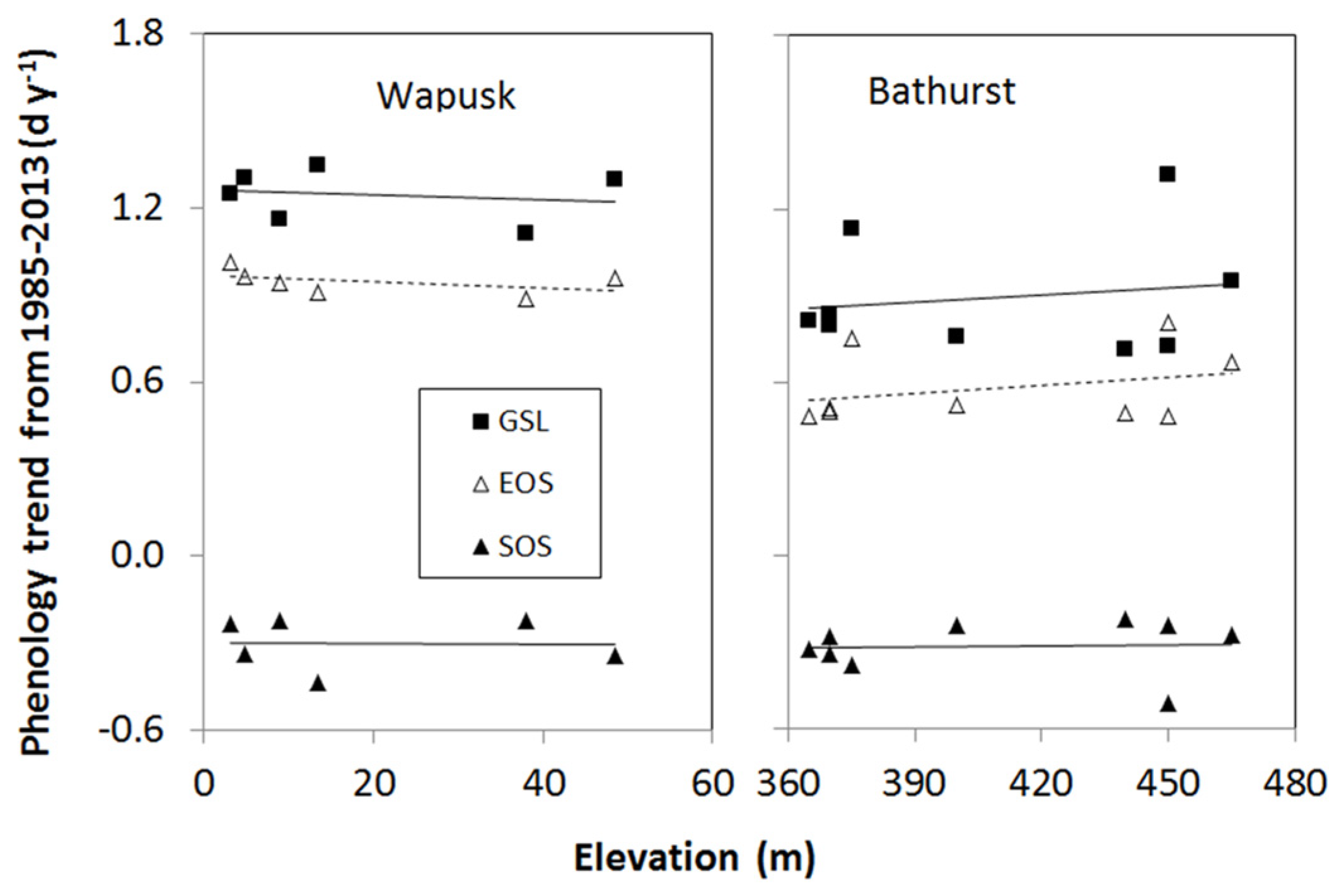
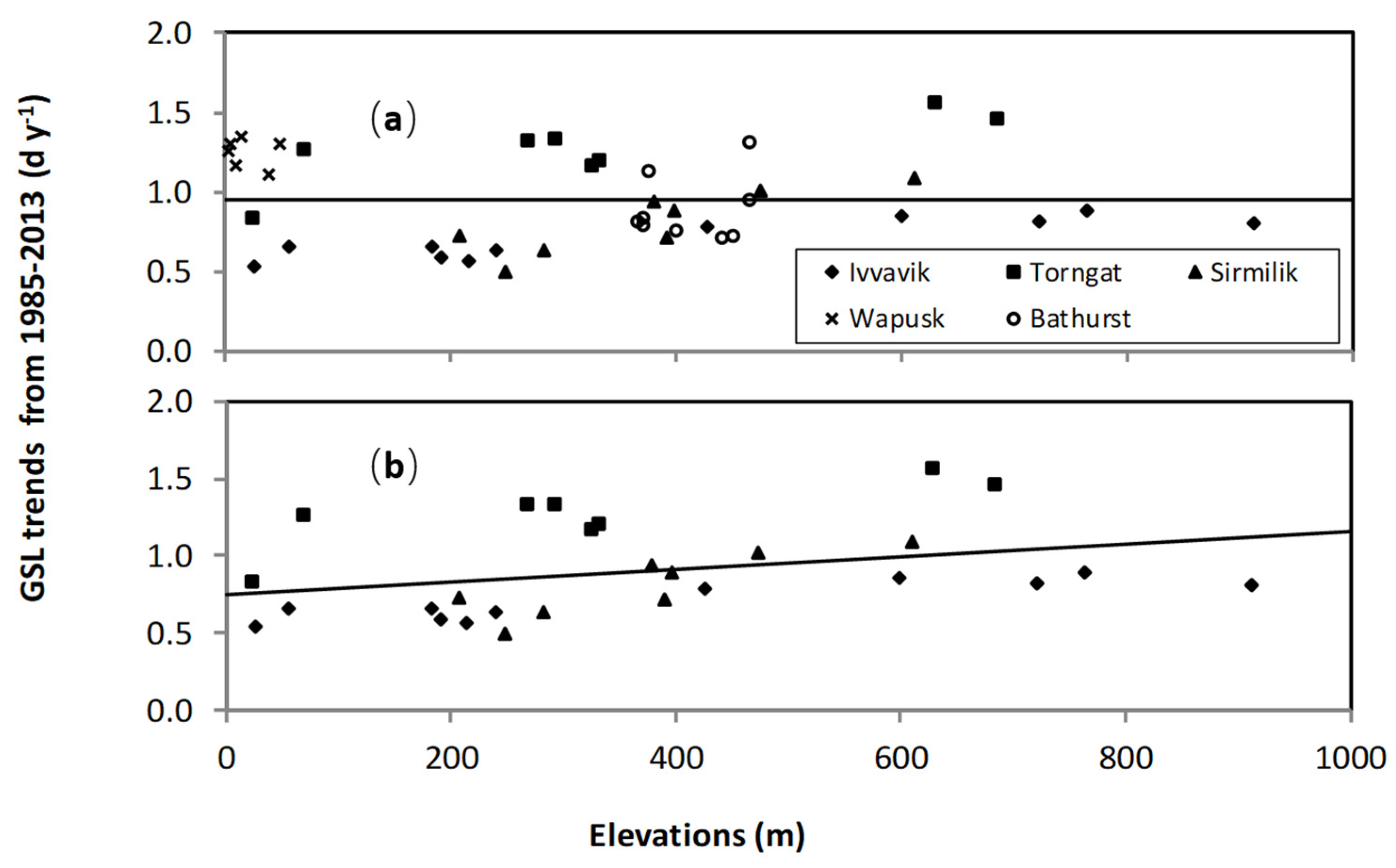
3.2. Elevation Dependency of Trends in Plant Phenology
4. Discussion
4.1. Hypotheses for Explaining the Elevation Dependency of Phenology Trends
4.2. Causes That Could Mask the Elevation Dependency
5. Conclusions
- (1)
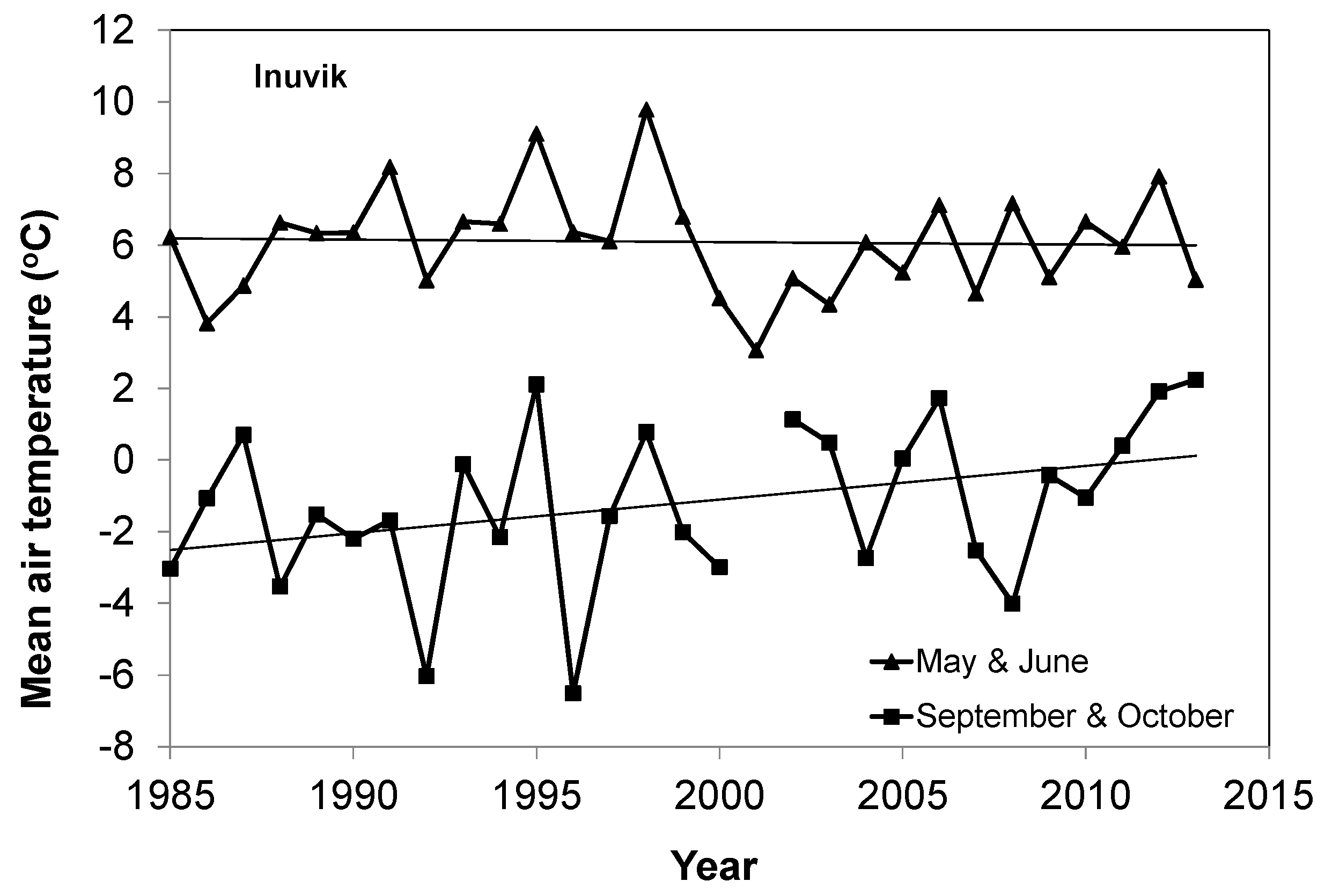
- The elevation dependency likely played an amplifier role upon a climate warming signal. When there was climate warming observed at weather stations in low-lying locations in a study area, we found significant increases in the magnitude of plant phenology change. For example, the magnitudes of long-term trends in EOS, SOS, and GSL increased significantly with elevation during 1985–2013 in all three mountainous study areas, except the SOS in the Ivvavik National Park.
- (2)
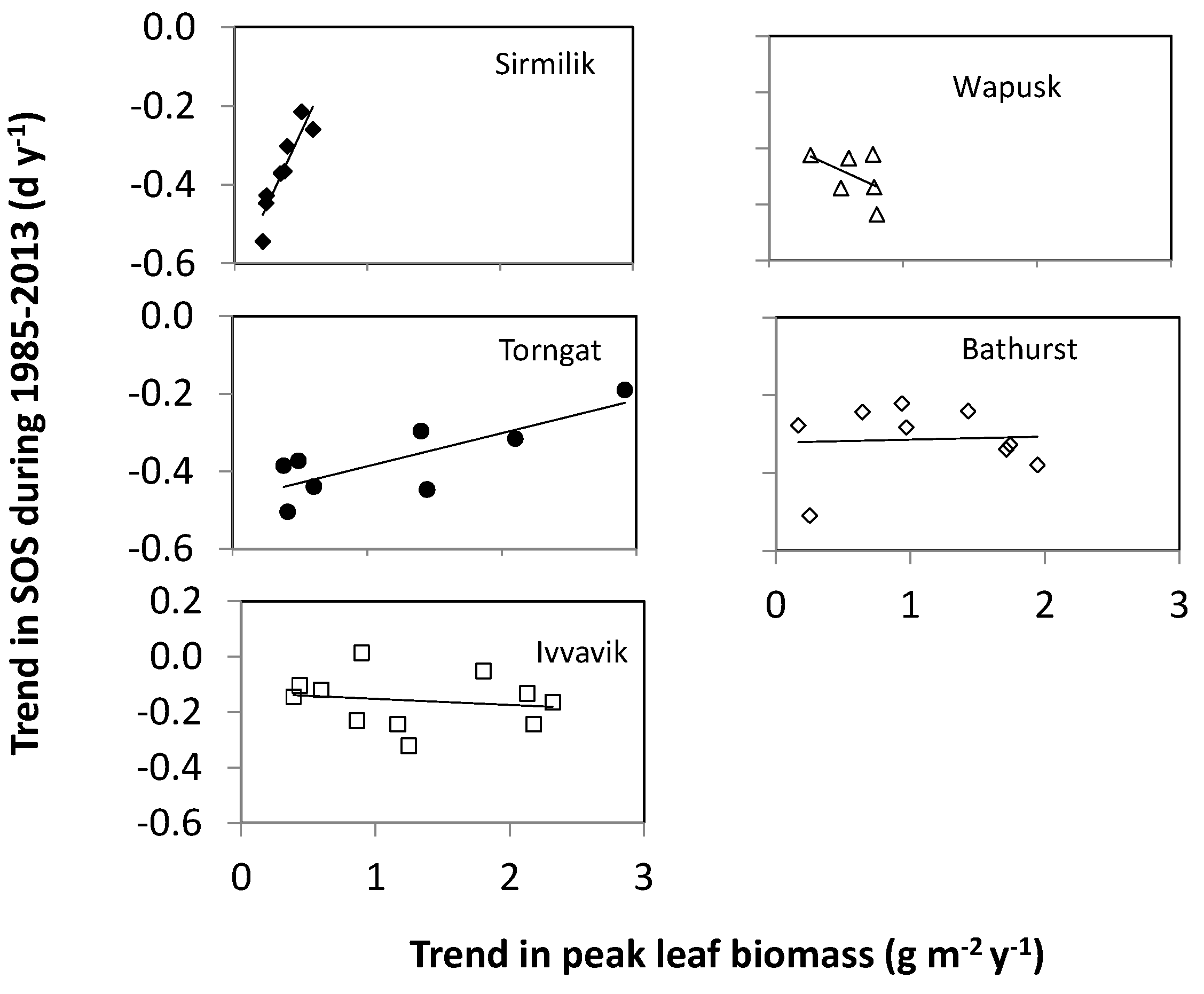
- There was a similar variation of vegetation types from low-lying locations to high elevation for all three mountainous study areas. If the variation of vegetation types was the main driver of changes in the long-term trends of SOS, then we should also have found a similar increase of its magnitude with elevation in the Ivvavik National Park. However, there was no significant increase in the magnitude of the long-term trend in SOS from 1985 to 2013 with elevation in the Ivvavik National Park. Correspondingly, the spring temperature at the Inuvik climate station located in the low-lying area of Ivvavik National Park decreased slightly by 0.2 °C from 1985 to 2013. Thus, long-term trends of SOS in the Ivvavik National Park indicated that the variation of vegetation types was not the primary driver of changes in the long-term trends of plant phenology. In addition, the peak leaf biomass values varied substantially for different land cover classes in the two flat study areas, similar to those in the three mountainous study areas. If the variation of vegetation types were the main driver of changes in plant phenology, then we would have detected a significant relationship in the long-term trends between the peak leaf biomass and plant phenology for all five study areas. However, we only found significant relationships in the long-term trends between the peak leaf biomass and SOS over the Sirmilik and the Torngat Mountains National Parks, but not the two flat study areas. Again, these results suggested that the variation of vegetation types was not the primary driver of changes in the long-term trends of plant phenology.
- (3)
- As exemplified by the long-term trends of SOS and spring temperature in the Ivvavik National Park during 1985 and 2013, if there was no increase in temperature, then an amplified no-increase with elevation would still be no-increase. Therefore, when data are pooled from different climate zones regionally or globally, the elevation dependency can be partially or totally masked because of this no-warming or even cooling sites.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McBean, G.; Alekseev, G.; Chen, D. Arctic Climate: Past and Present; IPCC Working Group I Report; Cambridge University Press: Cambridge, UK, 2005; p. 127. [Google Scholar]
- IPCC. IPCC, Climate Change 2013: The Physical Science Basis, Contribution of Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013; p. 1535. [Google Scholar]
- Beniston, M.; Rebetez, M. Regional behavior of minimum temperatures in Switzerland for the period 1979–1993. Theor Appl. Climatol. 1996, 53, 231–243. [Google Scholar] [CrossRef]
- Liu, X.; Cheng, Z.; Yan, L.; Yin, Z.Y. Elevation dependency of recent and future minimum surface air temperature trends in the Tibetan Plateau and its surroundings. Glob. Planet. Chang. 2009, 68, 164–174. [Google Scholar] [CrossRef]
- Ding, M.J.; Li, L.H.; Nie, Y. Spatial-temporal variation of spring phenology in Tibetan Plateau and its linkage to climate change from 1982 to 2012. J. Mt. Sci. 2016, 13, 83–94. [Google Scholar] [CrossRef]
- Thompson, L.G.; Mosley-Thompson, E.; Davis, M.E. Tropical glacier and ice core evidence of climate change on annual to millennial time scales. Clim. Chang. 2003, 59, 137–155. [Google Scholar] [CrossRef]
- Schmidt, L.S.; Aðalgeirsdóttir, G.; Guðmundsson, S. The importance of accurate glacier albedo for estimates of surface mass balance on Vatnajökull: Evaluating the surface energy budget in a regional climate model with automatic weather station observations. Cryosphere 2017, 11, 1665–1684. [Google Scholar] [CrossRef] [Green Version]
- Boers, N. Observation-based early-warning signals for a collapse of the Atlantic Meridional Overturning Circulation. Nat. Clim. Chang. 2021, 11, 680–688. [Google Scholar] [CrossRef]
- Barry, R.G. Mountain Weather and Climate, 3rd ed; Cambridge University Press: New York, NY, USA, 2008. [Google Scholar]
- Grünewald, T.; Bühler, Y.; Lehning, M. Elevation dependency of mountain snow depth. Cryosphere Discuss. 2014, 8, 3665–3698. [Google Scholar]
- Giorgi, F.; Hurrell, J.W.; Marinucci, M.R. Elevation dependency of the surface climate change signal: A model study. J. Clim. 1997, 10, 288–296. [Google Scholar] [CrossRef] [Green Version]
- Rangwala, I.; Miller, J.R. Twentieth century temperature trends in Colorado’s San Juan Mountains. Arct. Antarct. Alp. Res. 2010, 42, 89–97. [Google Scholar] [CrossRef]
- Kuhn, M.; Olefs, M. Elevation-Dependent Climate Change in the European Alps 2020. Available online: https://www.semanticscholar.org/paper/Elevation-Dependent-Climate-Change-in-the-European-Kuhn-Olefs/9932cb9c2475e75415387843d660b9135a24e2ae (accessed on 7 June 2021).
- Vuille, M.; Bradley, R.S. Mean annual temperature trends and their vertical structure in the tropical Andes. Geophys Res Let. 2000, 27, 3885–3888. [Google Scholar] [CrossRef] [Green Version]
- Pepin, N.C. Twentieth-century change in the climate record for the Front Range, Colorado, USA. Arct. Antarct. Alp. Res. 2000, 32, 135–146. [Google Scholar] [CrossRef]
- Pepin, N.C.; Losleben, M.L. Climate change in the Colorado Rocky Mountains: Free air versus surface temperature trends. Int. J. Clim. 2002, 22, 311–329. [Google Scholar] [CrossRef]
- Pepin, N.C.; Seidel, D.J. A global comparison of surface and free-air temperatures at high elevations. J. Geophys. Res. 2005, 110, D03104. [Google Scholar] [CrossRef]
- Pepin, N.C.; Lundquist, J.D. Temperature trends at high elevations: Patterns across the globe. Geophys Res Let. 2008, 35, L14701. [Google Scholar] [CrossRef] [Green Version]
- Pepin, N.; Bradley, R.; Diaz, H.F. Elevation-dependent warming in mountain regions of the world. Nat. Clim. Chang. 2015, 5, 424–430. [Google Scholar] [CrossRef] [Green Version]
- Gordo, O.; Sanz, J.J. Impact of climate change on plant phenology in Mediterranean ecosystems. Glob. Chang. Biol. 2010, 16, 1082–1106. [Google Scholar] [CrossRef]
- Borner, A.P.; Kielland, K.; Walker, M.D. Effects of Simulated Climate Change on Plant Phenology and Nitrogen Mineralization in Alaskan Arctic Tundra. Arct. Antarct. Alp. Res. 2008, 40, 27–38. [Google Scholar] [CrossRef] [Green Version]
- Myneni, R.; Keeling, C.; Tucker, C. Increased plant growth in the northern high, latitudes from 1981 to 1991. Nature 1997, 386, 698–702. [Google Scholar] [CrossRef]
- Karlsen, S.R.; Elvebakk, A.; Høgda, K.A.; Grydeland, T. Spatial and Temporal Variability in the Onset of the Growing Season on Svalbard, Arctic Norway—Measured by MODIS-NDVI Satellite Data. Remote Sens. 2014, 6, 8088–8106. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Foy, N.; Olthof, I.; Zhang, Y.; Fraser, R.; Latifovic, R.; Poitevin, J.; Zorn, P.; McLennan, D. A Biophysically-based and Objective Satellite Seasonality Observation Method for Applications over the Arctic. Int. J. Remote Sens. 2014, 35, 6742–6763. [Google Scholar] [CrossRef]
- Chen, W.; Zorn, P.; White, L. Decoupling between Plant Productivity and Growing Season Length under a Warming Climate in Canada’s Arctic. Am. J. Clim. Chang. 2016, 5, 344–359. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Adamczewski, J.Z.; White, L. Cumulative impacts of habitat conditions on caribou phenology quantified using satellite observations for the Bathurst herd in Arctic Canada. Polar Biol. 2018, 41, 953–967. [Google Scholar] [CrossRef]
- Wiken, E.B. (Ed.) Terrestrial Ecozones of Canada; Ecological Land Classification Series No. 19; Environment Canada: Hull, QC, Canada, 1986; p. 26 + map. [Google Scholar]
- Latifovic, R.; Cihlar, J.; Chen, J. Comparison of BRDF models for the normalization of satellite optical data to a standard sun-target-sensor geometry. IEEE Trans. Geosci. Remote Sens. 2003, 41, 1889–1898. [Google Scholar] [CrossRef]
- Latifovic, R.; Trishchenko, A.; Chen, J. Generating historical AVHRR 1 km baseline satellite data records over Canada suitable for climate change studies. Can. J. Remote Sens. 2005, 31, 324–348. [Google Scholar] [CrossRef] [Green Version]
- Khlopenkov, K.; Trishchenko, A. SPARC: New cloud, clear-snow/ice and cloud shadow detection scheme for historical AVHHR 1-km observations over Canada. J. Atmos. Ocean. Technol. 2006, 24, 322–343. [Google Scholar] [CrossRef]
- Latifovic, R.; Pouliot, D.; Dillabaugh, C. Identification and correction of systematic error in NOAA AVHRR long-term satellite data record. Remote Sens. Environ. 2012, 127, 84–97. [Google Scholar] [CrossRef]
- Chen, W.; Li, J.; Zhang, Y. Relating biomass and leaf area index to non-destructive measurements for monitoring changes in arctic vegetation. Arctic 2009, 62, 281–294. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Chen, W.; Leblanc, S.G.; Henry, G. Digital photograph analysis for measuring percent plant cover in the Arctic. Arctic 2010, 63, 315–326. [Google Scholar] [CrossRef] [Green Version]
- Olthof, I.; Latifovic, R.; Pouliot, D. Development of a circa 2000 land cover map of northern Canada at 30 m resolution from Landsat. Can. J. Remote Sens. 2009, 35, 152–165. [Google Scholar] [CrossRef]
- Olthof, I.; Fraser, R.H. Mapping northern land cover fractions using Landsat ETM+. Remote Sens. Environ. 2007, 107, 496–509. [Google Scholar] [CrossRef]
- Li, J.; Chen, W. A rule-based method for mapping Canada’s wetlands using optical, radar and DEM data. Int. J. Remote Sens. 2005, 26, 5051–5069. [Google Scholar] [CrossRef]
- Fraser, R.H.; McLennan, D.; Ponomarenko, S.; Olthof, I. Image-based predictive ecosystem mapping in Canadian arctic parks. Int. J. Appl. Earth Obs. Geoinf. 2012, 14, 129–138. [Google Scholar] [CrossRef]
- Chen, W.; Foy, N.; Olthof, I. Evaluating and reducing errors in seasonal profiles of AVHRR vegetation indices over a Canadian northern national park using cloudiness index. Int. J. Remote Sens. 2013, 34, 4320–4343. [Google Scholar] [CrossRef]
- Chen, W.; Chen, W.R.; Li, J. Mapping aboveground and foliage biomass over the Porcupine caribou habitat in northern Yukon and Alaska using Landsat and JERS-1/SAR data. In Remote Sensing of Biomass: Principles and Applications; Fatoyinbo, T., Ed.; InTECH: Ottawa, ON, Canada, 2012; pp. 231–252. ISBN 978-953-51-0313-4. [Google Scholar]
- Chen, W.; Zorn, P.; Chen, Z. Propagation of errors associated with scaling foliage biomass from field measurements to remote sensing data over a Canada’s northern national park. Remote Sens. Environ. 2013, 130, 205–218. [Google Scholar] [CrossRef]
- Jamieson, M.A.; Trowbridge, A.M.; Raffa, K.F.; Lindroth, R.L. Consequences of climate warming and altered Precipitation Patterns for Plant-Insect and Multitrophic Interactions. Plant Physiol. 2012, 160, 1719–1727. [Google Scholar] [CrossRef] [Green Version]
- Carbognani, M.; Bernareggi, G.; Perucco, F. Microclimatic controls and warming effects on flowering time in alpine snowbeds. Oecologia 2016, 182, 573–585. [Google Scholar] [CrossRef] [PubMed]
- Mulder, C.P.H.; Iles, D.T.; Rockwell, R.F. Increased variance in temperature and lag effects alter phenological responses to rapid warming in a subarctic plant community. Glob. Chang. Biol. 2017, 23, 801–814. [Google Scholar] [CrossRef]
- Sturm, M.; McFadden, J.P.; Liston, G.E.; Chapin, F.S., III; Racine, C.H.; Holmgren, J. Snow-shrub interactions in Arctic tundra: A hypothesis with climate implications. J. Clim. 2001, 14, 336–344. [Google Scholar] [CrossRef] [Green Version]
- Liston, G.E.; McFadden, J.P.; Sturm, M.; Pielke, R.A., Sr. Modeled changes in Arctic tundra snow, energy, and moisture fluxes due to increased shrubs. Glob. Chang. Biol. 2002, 9, 17–32. [Google Scholar] [CrossRef]
- Sweet, S.K.; Gough, L.; Griffin, K.L.; Boelman, N.T. Tall Deciduous Shrubs Offset Delayed Start of Growing Season Through Rapid Leaf Development in the Alaskan Arctic Tundra. Arct. Antarct. Alp. Res. 2014, 46, 682–697. [Google Scholar] [CrossRef]
- Berner, L.T.; Alexander, H.D.; Loranty, M.M. Biomass allometry for alder, dwarf birch, and willow in boreal forest and tundra ecosystems of far northeastern Siberia and north-central Alaska. For. Ecol. Manag. 2015, 337, 110–118. [Google Scholar] [CrossRef]
- Sturm, M.; Schimel, J.; Michaelson, G. Winter biological processes could help convert arctic tundra to shrubland. BioScience 2005, 55, 17–26. [Google Scholar] [CrossRef]
- Cohen, J.; Pulliainnen, J.; Ménard, C.B. effect of reindeer grazing on snowmelt, albedo and energy balance based on satellite data analysis. Remote Sens. Environ. 2013, 135, 107–117. [Google Scholar] [CrossRef]
- Loranty, M.M.; Berner, L.T.; Goetz, S.J. Vegetation controls on northern high latitude snow-albedo feedback: Observations and CMIP5 model predictions. Glob. Chang. Biol. 2013, 20, 594–606. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Area | Area (km2) | Latitude/Longitude at the Venter | Ecozone | Elevation Range (m) | Closest Climate Station |
|---|---|---|---|---|---|
| Ivvavik | 9750 | 69°36′00″ N 140°10′00″ W | Taiga cordillera southern Arctic and | 0–1760 | Inuvik |
| Sirmilik | 22,200 | 72°50′4″ N 80°34′55″ W | Arctic cordillera and northern Arctic | 0–1944 | Pond Inlet |
| Torngat | 9700 | 59°22’12″ N 63°38’48″ W | Arctic cordillera | 0–1652 | Nain |
| Wapusk | 11,475 | 57°51′11″ N 93°22′35″ W | Hudson plains | 0–86 | Churchill |
| Bathurst | 112,000 | 65°8′5″ N 111°7′ W | Southern Arctic | 365–465 | Lupin |
| Label | Land Cover Class Name | Mean Elevation (M) | Trends (d y−1) | ||
|---|---|---|---|---|---|
| SOS | EOS | GSL | |||
| I1 | Alpine slope | 764 | −0.24 | 0.64 *** | 0.88 ** |
| I2 | Willow-horsetail wet slope | 599 | −0.23 | 0.62 ** | 0.85 ** |
| I3 | Rock lichen | 912 | −0.10 | 0.70 *** | 0.81 * |
| I4 | Willow-birch moist slope | 722 | −0.32 | 0.49 ** | 0.81 ** |
| I10 | Willow floodplain | 215 | 0.01 | 0.57 ** | 0.56 * |
| I18 | Cotton-grass tussock | 241 | −0.13 | 0.51 ** | 0.63 * |
| I20 | Willow-sedge pediment drainage channel | 56 | −0.16 | 0.52 * | 0.66 ** |
| I22 | Sand/silt | 426 | −0.14 | 0.64 ** | 0.79 * |
| I23 | Hedysar-avens inactive alluvial Terrace | 26 | −0.12 | 0.38 | 0.53 |
| I25 | Willow-coltsfoot drainage channel | 183 | −0.24 | 0.45 | 0.65 * |
| I26 | Sedge tussock | 192 | −0.05 | 0.54 ** | 0.59 * |
| S1 | Tussock graminoid tundra | 249 | −0.22 | 0.28 | 0.50 |
| S2 | Wet sedge | 282 | −0.26 | 0.37 | 0.63 * |
| S3 | Moist-dry graminoids/dwarf shrub | 391 | −0.30 | 0.41 * | 0.71 * |
| S7 | Prostrate dwarf shrub | 208 | −0.37 ** | 0.36 | 0.73 ** |
| S8 | Sparsely vegetated bedrock | 379 | −0.37 | 0.57 | 0.94 * |
| S9 | Sparsely vegetated till-colluvium | 474 | −0.45 * | 0.56 * | 1.01 ** |
| S10 | Bare soil/cryptogam crust-frost boils | 398 | −0.43 * | 0.46 ** | 0.89 ** |
| S12 | Barren | 611 | −0.54 * | 0.53 | 1.08 |
| T16 | Deciduous shrub (>75% cover) | 69 | −0.32 | 0.92 *** | 1.27 *** |
| T23 | Herb-shrub | 292 | −0.30 | 1.08 *** | 1.34 *** |
| T24 | Shrub-herb-lichen-bare | 23 | −0.19 | 0.66 ** | 0.84 ** |
| T26 | Lichen-shrubs-herb, bare soil, rock outcrop | 267 | −0.45 | 0.90 *** | 1.33 *** |
| T28 | Low veg cover (bare soil, rock outcrop) | 629 | −0.50 | 1.05 ** | 1.57 ** |
| T35 | Lichen barren | 331 | −0.44 | 0.76 *** | 1.20 *** |
| T36 | Lichen-shrub-herb-bare | 325 | −0.37 | 0.80 *** | 1.16 ** |
| T38 | Rock outcrop low vegetation cover | 684 | −0.39 | 1.01 ** | 1.46 * |
| W3 | Dryas heath upland | 3 | −0.24 | 1.02 *** | 1.25 *** |
| W6 | Lichen peat plateau bog | 38 | −0.22 | 0.89 *** | 1.11 *** |
| W9 | Lichen melt pond bog | 48 | −0.34 | 0.96 *** | 1.30 *** |
| W10 | Sedge fen | 5 | −0.34 | 0.97 *** | 1.31 *** |
| W11 | Shrub fen | 13 | −0.44 | 0.91 *** | 1.35 *** |
| W12 | Shrub sedge fen | 9 | −0.22 | 0.94 *** | 1.16 ** |
| B16 | Shrub moist | 365 | −0.33 ** | 0.48 ** | 0.81 *** |
| B17 | Shrub mesic | 370 | −0.34 * | 0.50 ** | 0.84 *** |
| B23 | Herb | 400 | −0.24 | 0.52 *** | 0.76 *** |
| B26 | Lichen-shrubs-herb, bare soil, rock outcrop | 370 | −0.28 * | 0.51 ** | 0.80 *** |
| B28 | Low veg cover (bare soil, rock outcrop) | 465 | −0.28 * | 0.67 ** | 0.95 ** |
| B35 | Lichen barren | 440 | −0.22 | 0.49 * | 0.71 ** |
| B36 | Lichen-shrub-herb-bare | 450 | −0.24 | 0.48 * | 0.72 ** |
| B38 | Rock outcrop, low vegetation cover | 450 | −0.51 ** | 0.81 *** | 1.32 *** |
| B41 | Low vegetation cover | 375 | −0.38 ** | 0.75 *** | 1.13 *** |
| Climate Station | Study Area | Period | Trended Change (Days) | p-Value |
|---|---|---|---|---|
| Pond Inlet | Sirmilik | Spring | 1.12 | 0.297 |
| Fall | 3.33 | 0.013 | ||
| Churchill | Wapusk | Spring | 0.08 | 0.647 |
| Fall | 3.10 | 0.003 | ||
| Nain | Torngat | Spring | 1.45 | 0.056 |
| Fall | 2.11 | 0.001 | ||
| Inuvik | Ivvavik | Spring | −0.19 | 0.863 |
| Fall | 2.63 | 0.069 | ||
| Lupin | Bathurst | Spring | 1.50 | 0.145 |
| Fall | 2.73 | 0.011 |
| Study Area | Phenology | Slope (d y−1 m−1) | Intercept (d y−1) | R2 | p-Value | n |
|---|---|---|---|---|---|---|
| Ivvavik | SOS | −0.000130 | −0.108 | 0.17 | 0.210 | 11 |
| EOS | 0.000224 | 0.463 | 0.52 | 0.013 | 11 | |
| GSL | 0.000355 | 0.566 | 0.77 | 0.0004 | 11 | |
| Sirmilik | SOS | −0.000660 | −0.119 | 0.65 | 0.016 | 8 |
| EOS | 0.000617 | 0.210 | 0.59 | 0.025 | 8 | |
| GSL | 0.001308 | 0.322 | 0.71 | 0.008 | 8 | |
| Torngat | SOS | −0.000300 | −0.272 | 0.48 | 0.057 | 8 |
| EOS | 0.000387 | 0.764 | 0.40 | 0.092 | 8 | |
| GSL | 0.000746 | 1.025 | 0.63 | 0.019 | 8 | |
| Wapusk | SOS | −0.000170 | −0.296 | 0.001 | 0.944 | 6 |
| EOS | −0.001070 | 0.968 | 0.21 | 0.367 | 6 | |
| GSL | −0.000900 | 1.264 | 0.035 | 0.722 | 6 | |
| Bathurst | SOS | 0.000123 | −0.364 | 0.003 | 0.886 | 9 |
| EOS | 0.000911 | 0.206 | 0.087 | 0.441 | 9 | |
| GSL | 0.000789 | 0.570 | 0.025 | 0.683 | 9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, W.; White, L.; Leblanc, S.G.; Latifovic, R.; Olthof, I. Elevation-Dependent Changes to Plant Phenology in Canada’s Arctic Detected Using Long-Term Satellite Observations. Atmosphere 2021, 12, 1133. https://doi.org/10.3390/atmos12091133
Chen W, White L, Leblanc SG, Latifovic R, Olthof I. Elevation-Dependent Changes to Plant Phenology in Canada’s Arctic Detected Using Long-Term Satellite Observations. Atmosphere. 2021; 12(9):1133. https://doi.org/10.3390/atmos12091133
Chicago/Turabian StyleChen, Wenjun, Lori White, Sylvain G. Leblanc, Rasim Latifovic, and Ian Olthof. 2021. "Elevation-Dependent Changes to Plant Phenology in Canada’s Arctic Detected Using Long-Term Satellite Observations" Atmosphere 12, no. 9: 1133. https://doi.org/10.3390/atmos12091133
APA StyleChen, W., White, L., Leblanc, S. G., Latifovic, R., & Olthof, I. (2021). Elevation-Dependent Changes to Plant Phenology in Canada’s Arctic Detected Using Long-Term Satellite Observations. Atmosphere, 12(9), 1133. https://doi.org/10.3390/atmos12091133