3.1. Phenomenon Analysis
Due to the cohesion of the riverbank, erosion occurred only when the stress produced by the shear flow exceeded a critical value [
46]. Observing the evolution of a meandering river channel, the river bend developed from upstream and downstream in the initial stage. With the development of meander, the flow condition downstream gradually matured, while the curvature of river increased. The downstream evolved according to its own evolution mechanism and played an important role in restricting the upstream. The inflow angle had a short-term effect on the evolution of the river channel and accelerated the development in the initial stage, but the river would reach a balance according to the flow and sediment condition in the long run.
Table 2 shows the changes in the basic variables in the evolution of the seven groups of experiments and the retained planform.
During the experiment, the bottom of the riverbank began to be scoured by the flow, creating a cantilever structure. Collapse would occur if the riverbank showed instability [
47]. However, for those channels with vegetation on their bank, even though scouring would still occur and the vegetation’s roots would be exposed to the air, which would lead to a local scour (
Figure 4a), less slump block phenomena occurred due to the reinforcement effect created by the vegetation roots. Therefore, the influence of vegetation on the erosion rate of river banks is closely related to its density, root development and arrangement mode. The scouring process could be divided into two processes: vegetation scouring and isolation, and vegetation collapse and washing away by flow. About 1–10 min passed before the plant showed instability. Since the initial velocity was about 0.35 m/s, the plant scoured easily and fell very quickly. About 2–20 min later, the plant washed away after it fell down, and the falling plant in the bed increased the local roughness and protected the bank from scouring (
Figure 4b). Consequently, the vegetation increased the local flow disturbance. At the initial period of the evolution of the river channel, the width and depth were relatively small and the flow was in a state of supercritical flow, which created a turbulent area in the vegetation riparian zone, and the bed sand particles encountered upward vortexes in the process leaping from the bed. When the sand fell out of the vortex, it entered the main stream, resulting in intense scouring along the vegetation belt. The main streamline scattered due to the flow turbulence caused by the vegetation, and staggered deposition occurred along the center of the channel, which led to the slow development of river meander. However, the vegetation had a reinforcing effect on the riverbank, which increased the resistance along the riverbank. As the evolution of the river channel continued, plants falling in the channel increased the local hydraulic roughness, and the flow velocity among the vegetation decreased, which increased the amount the deposition and markedly decreased the erosion rate along the riverbank.
3.2. Hydraulic Characteristics of Flow
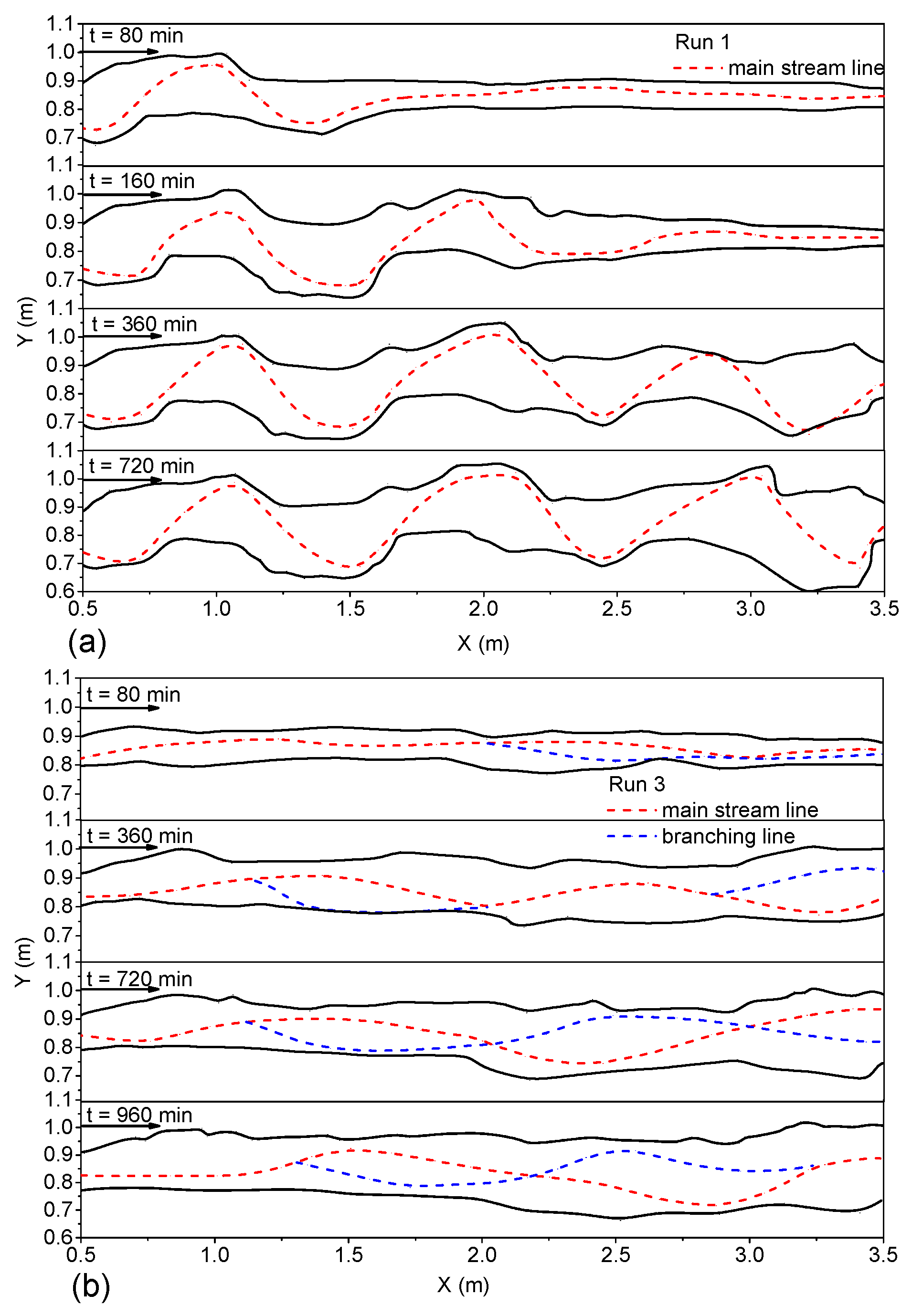
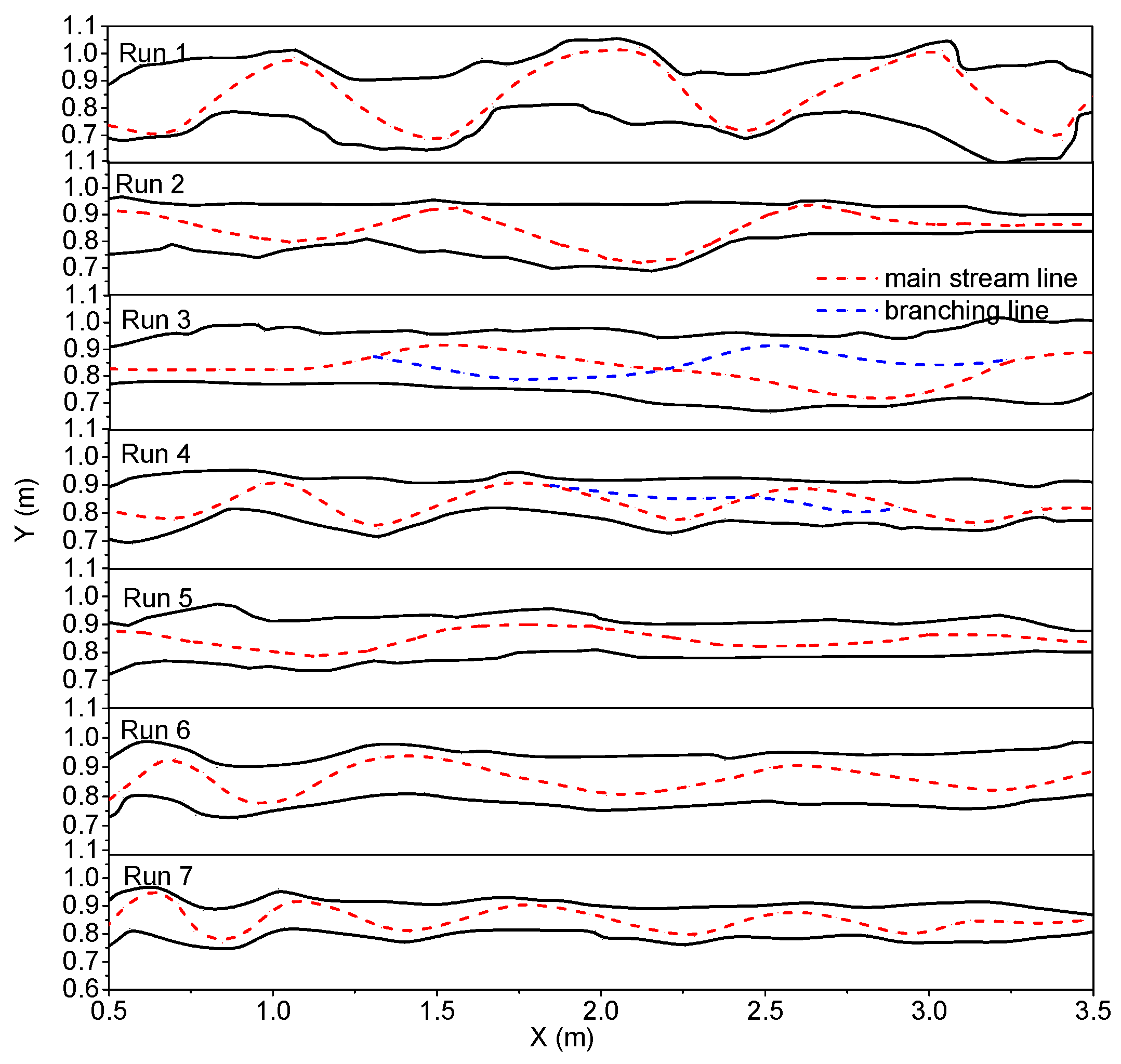
During the experiment, particles were manually scattered onto the river channel inlet, and most of the particles flowed to the axis of the water flow. The surface flow field was measured with PTV, and the particle trajectories were recorded by the fixed cameras. The trajectories were extracted, and the dynamic axis of flow at different times was plotted (
Figure 5a,b). Through the observation of different groups of river channel evolution, we found that, due to the influence of the upstream inflow angle, the upstream began to bend, thus affecting the downstream. The downstream evolution also had a constraining effect on the upstream, in accordance with the evolution mechanism of the river channel itself. In the initial stage, the downstream evolution lagged behind the upstream, and the curvature of bend had downward transitivity. The influence of inflow angle on the evolution of river bend was only short term. At the initial time, the evolution of the river bend was accelerated, but in the longer runs, the river channel reached its equilibrium state under different boundary conditions, such as water flow, sediment, etc. After the initial 80 min, the downstream river bank was basically limited to river bank collapse [
38], which was mainly due to upstream disturbances that had not yet fully propagated to the downstream. Observations of Run 1 demonstrated that, during river channel evolution, the flow dynamic axis was close to the inner-bank and leaned toward the downstream along the inner-bank rushing point (
Figure 5a). At 360 min, the flow dynamic axis maintained the same curvature, and a wavelength developed along the whole river channel; a typical meandering river channel formed.
The evolution process of Run 3 is shown in
Figure 5b. The river channel evolved at 80 min; the main stream adjustment was accompanied by the formation of the downstream branch; and the branch node gradually moved upstream. Just at 180 min, the river formed a steady channel thread. At 360 min, due to the influence of the downstream branch, along the whole reach, two larger anabranches gradually formed. Between 1.5 and 2.0 m of the river reach, the main stream should have been bent, but due to the barrier of the vegetation bank, it developed straight. As seen from
Figure 6, along the left bank, three large silting bodies (dynamic sediment bodies) formed, and the water had to readjust its main path on the side of the silting bodies. As the evolution continued, at 600 min, the silting bodies near the right bank gradually disappeared, and those near the left bank gradually increased, forming a new silting body (
Figure 6, Dynamic Sediment Body 4). This phenomenon is often seen in actual rivers, as shown in
Figure 6b. On the Mataura River near a cattle flat, a tributary of the Red River, the vegetation zone is distributed on one bank side, whereas the other side contains meadow, and the main stream moves to the bank of the meadow, which has a lower scour resistance.
The asymmetry of riverbank erosion resistance (riverbank anti erosion properties) led to the swing of the main stream and the alternate development of the main stream and tributaries. The river channel will maintain the anabranching or braided pattern for a long time (
Figure 5b). However, this is not caused by the greater difference in erosion resistance on both river bank sides, but by the more frequent adjustment of the river regime. Taking Run 2 as an example, given the dense vegetation distributed on the left bank, the interaction between the two sides of the river bank did not produce a larger mutual disturbance, and the main stream deviated away from the left bank to seek a new hydrodynamic environment.
The existence of riparian vegetation slows down the lateral migration of the river. In laboratory experiments, the riparian vegetation has played a role in stabilizing the point bar, strengthening the river bank resistance, which will cause the river to develop in the direction of low bending. The time for the formation of a more stable flow dynamic axis of vegetated revetment river was longer than that without vegetation; especially for the situation where vegetation revetment was asymmetric on the left and right banks, and even the main stream line had had a meandering trend. Due to the influence of local vegetation flow resistance, the flow velocity along the cross-section was redistributed and the original bending trend was destroyed. Runs 2, 3 and 4 completed the riverbed adjustment at 400, 180 and 600 min, respectively.
Figure 7a,b shows the bed morphology of the 1.5–2.0 m reach in the case of riparian vegetation coverage of 40% in both unilateral and bilateral riparian areas. It can be seen that the Run 3 river bed was still in a state of disorder without a main stream trend, and Run 6 formed a clear main stream trend. It was found from the experimental observation. We found that when the vegetation was distributed on one side of the river bank, riparian vegetation cover along the river bank was dominant, and the adjustment process of flow dynamic axis was: scattered
micro bending/branching
meandering
straight (with scattered)
micro bending/branching.
Figure 8 shows the bed morphology of the 1.5–2.5 m reach of three groups of different riparian vegetation coverage at the stable time. The black part of the bed sand shows the main trajectory of sediment transport for extremely fine sediment, which reflects the concentration of the main stream and the increase in flow velocity under 80% vegetation coverage, which enhanced the sediment-carrying capacity and led to the concentrated scouring effect of the channel. As can be seen from
Figure 8a–c, in the absence of vegetation, the surface scour of the bed had a large scale and greater quantity, and the vortex action of the bend was obvious. When the river bank was planted with vegetation, the disturbance of vegetation to water flow destroyed the large-scale vortices that should have formed in the bend. The greater the vegetation coverage was, the more obvious the effect was; when the riverbank vegetation coverage was 80%, the riverbed had no obvious large-scale scoured pits.
As seen in
Figure 9, Runs 2, 3 and 4 included three groups of one-sided vegetation cover (left bank vegetation cover), showing the main stream, tributaries and final riparian erosion. At 960 min, a large deflection of the main stream formed, and the erosion rate of the two bank sides was different. Specifically, the center of the channel at the initial time was located at y = 0.85 m. Finally, the evolutions of the main stream lines were all inclined to the right bank. The curvature of the right bank line showed non-uniform distribution, and the curvature was larger; the left bank presented overall erosion recession, and the curvature was smaller. The difference in the erosion rate of the two bank sides will be analyzed in detail in
Section 3.3.
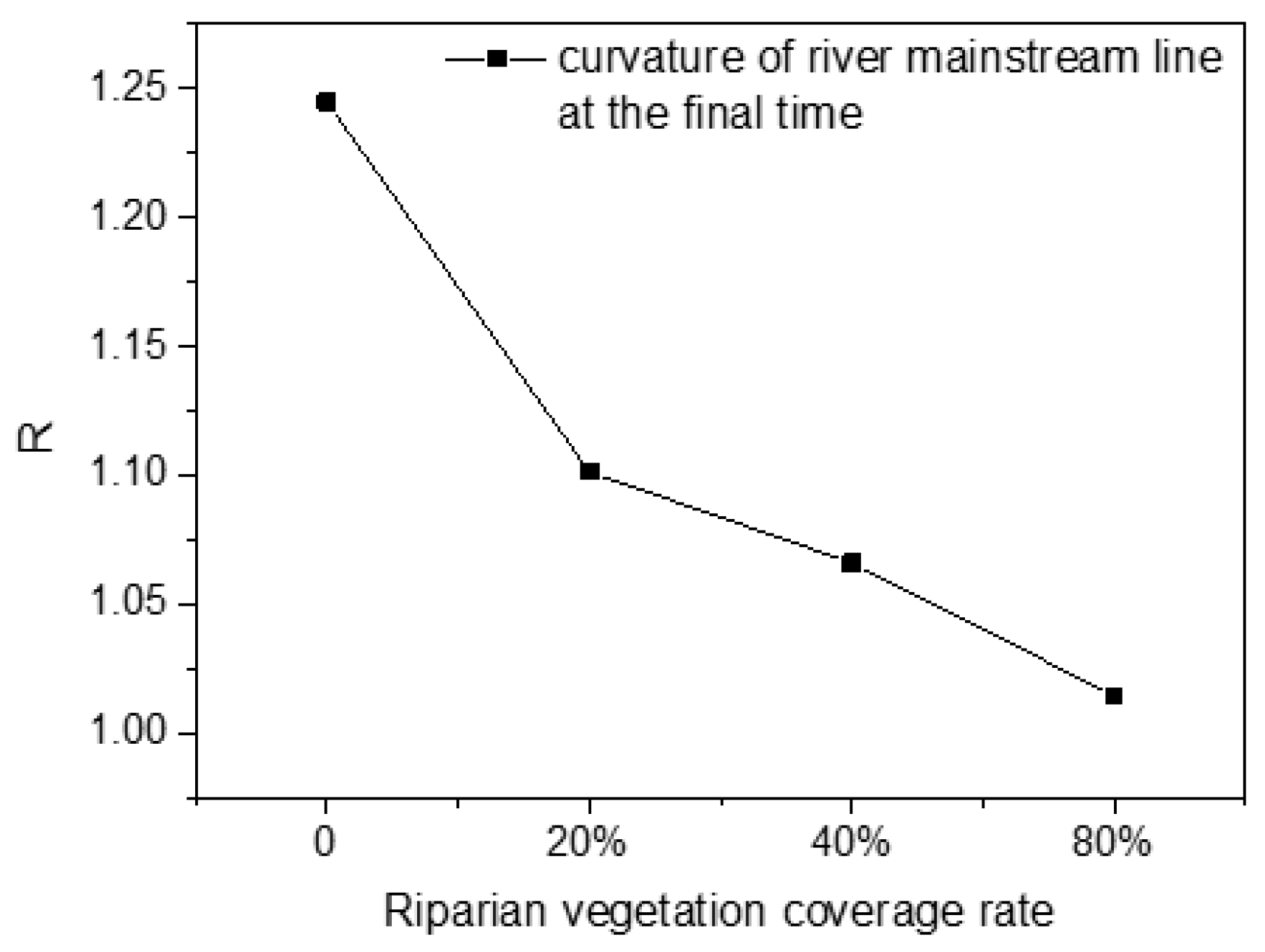
The relationship between the final curvature of river channel main stream and the vegetation coverage rate of bilateral riverbank cover is shown in
Figure 10. As can be seen from the diagram, vegetation coverage effectively reduced the curvature of the river channel. With the increase in the coverage rate, the curvature of the river decreased gradually and tended to one (
Figure 11a,b). When the vegetation coverage reached 80%, the curvature was 1.01, indicating that the river was close to a straight river type.
Ikeda et al. [
48] argued that the rates of meander migration depend on the velocity perturbation
from the mean velocity (the different between the velocity near the outside bank and the mean velocity).
represents the magnitude of the shear forces of the flow acting on the river banks [
49] in m/s. An erodibility coefficient
can be defined as the anti-erosion ability of the bank shear forces and has a strong relationship with vegetation coverage. The rate of meander migration
is the ratio of riparian erosion area to average riparian length in a given period of time [
50], in mm/min. The rate of meander migration can be expressed as:
For given channel geometry and flow conditions, the velocity distribution within a bend determines the distribution of flow shear forces on the river banks and bed [
51]. In this paper, the liner model of meander migration was used to explain the integrated influences of channel curvature on lateral migration rates [
48]. The model was based on the perturbation expansion on the curvature of the equation of motion for flow and sediment and was used to estimate linear cross-section profiles for depth-averaged flow velocities and bed elevations. The model has been recognized and validated by scholars with actual rivers [
48,
52,
53,
54]. We analyzed the relationship between the rates of meander migration and the velocity perturbation based on experimental measurement data and comparisons with actual river conditions [
50] and analyzed the meander migration characteristics in different riparian vegetation coverage.
The velocity perturbation equals the maximum near-bank velocity minus the mean cross-section velocity, that is . The maximum near-bank velocity is considered as the maximum near-bank surface velocity. The mean velocity is the discharge divided by the channel cross-section area. According to the difference in the final curvature along the river channel, which was divided into three sections for analysis (0.5–1.5 m, 1.5–2.5 m and 2.5–3.5 m), we respectively calculated the corresponding migration rate and velocity perturbation at different time intervals, because of the rapid widening of the channel at the initial stage and the riparian vegetation having fewer effects on the flow. Therefore, the relationship between the velocity perturbation and meander migration rate was analyzed only at 80–960 min. The time period was divided into three time intervals: 80–360 min, 360–720 min and 720–960 min. The maximum and mean velocity at the end of each stage were used for calculating the velocity perturbation.
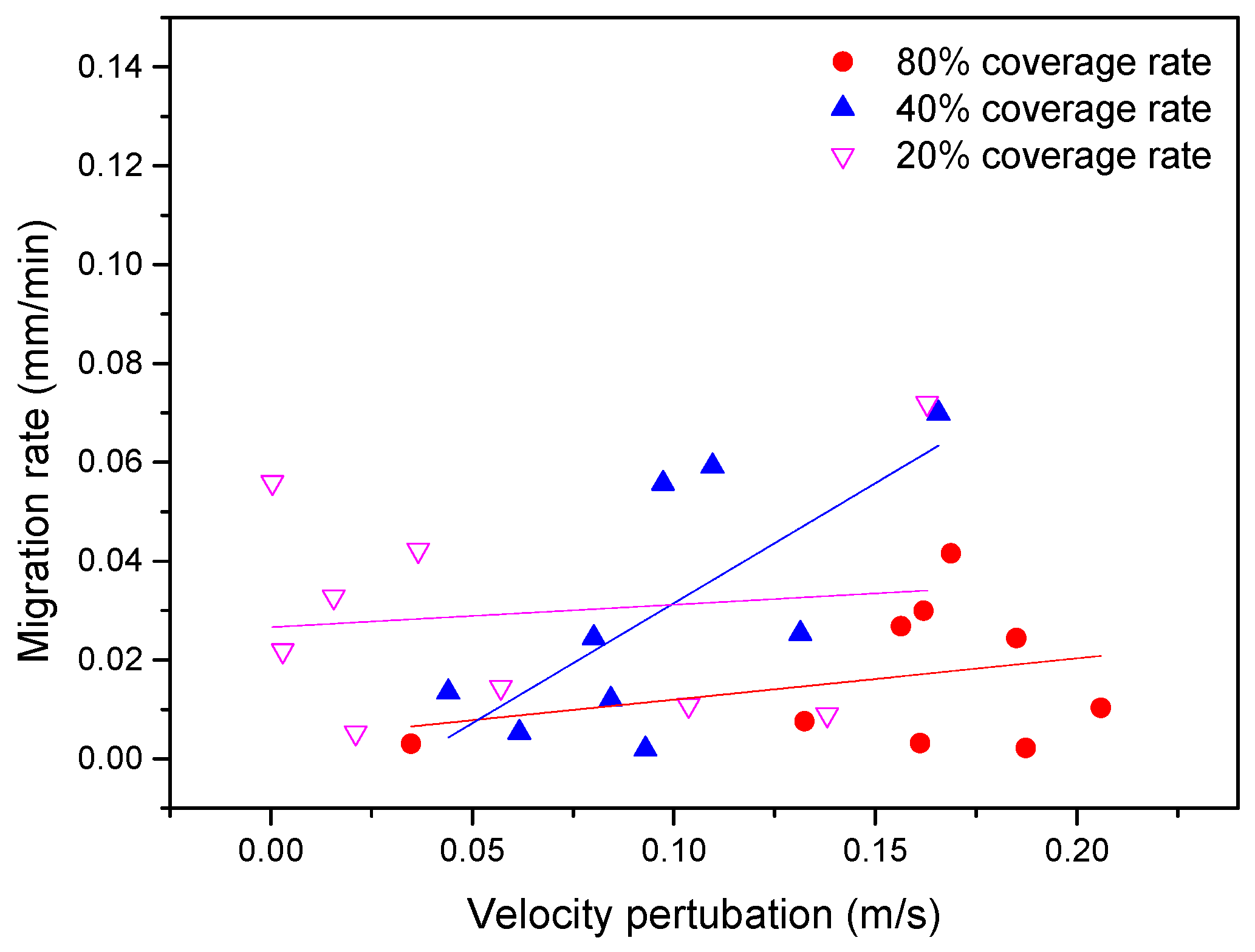
The results reflect a strong effect of riparian vegetation on rates of river channel migration. As shown in
Figure 12, for similar magnitudes of flow shear stress (expressed by velocity perturbation
), the migration rate for 20% riparian vegetation coverage was generally higher than that for 80% riparian vegetation coverage. The migration rate for a 40% riparian vegetation coverage rate was mostly higher than for an 80% riparian vegetation coverage. For the similar meander migration rate, the greater the vegetation coverage rate, the greater the flow shear stress, which is why riparian vegetation coverage leads the channel to become narrow and deep. The linear regressions corresponding to the three vegetation coverages were all positively related, indicating that the overall larger meander migration required a large flow shear stress. The slope of linear regressions drawn for the 40% riparian vegetation coverage rate values tended to be steeper, indicating that the channel meander migration rates under this coverage were more sensitive to increases in flow shear; the whole channel formed a large inhomogeneous lateral migration. The slope of linear regressions drawn for 80% and 20% riparian vegetation coverage rates were relatively smooth. The 80% riparian vegetation coverage had the lowest meander migration rates.
3.3. River Bank Evolution
The evolution of a meandering river channel is characterized by overall erosion recession of the river bank and the enlargement of the meandering amplitude (
Figure 4a). As the initial river bend is not in accordance with the river facies relation, the river channel gradually evolved from the initial straight river channel. The river widening process showed a trend of nonlinear exponential decrease [
55]. During the initial stage, a rapid widening of the channel would take place and the water flow could complete the initial bed-forming role in the short term [
56,
57]. After 80 min, the river channel extending was shown by the development of sediment erosion and deposition. We observed that the evolution of river banks can be divided into three periods: rapid extension, transition and stable. Therefore, according to the experimental observation of river channel evolution law, the river channel evolution was divided into four time intervals: 0–80 min, 80–360 min, 360–720 min and 720–960 min, to study the variation law of the channel widening rate. Each stage takes its intermediate time as the correspondent time of the average river bank migration rate.
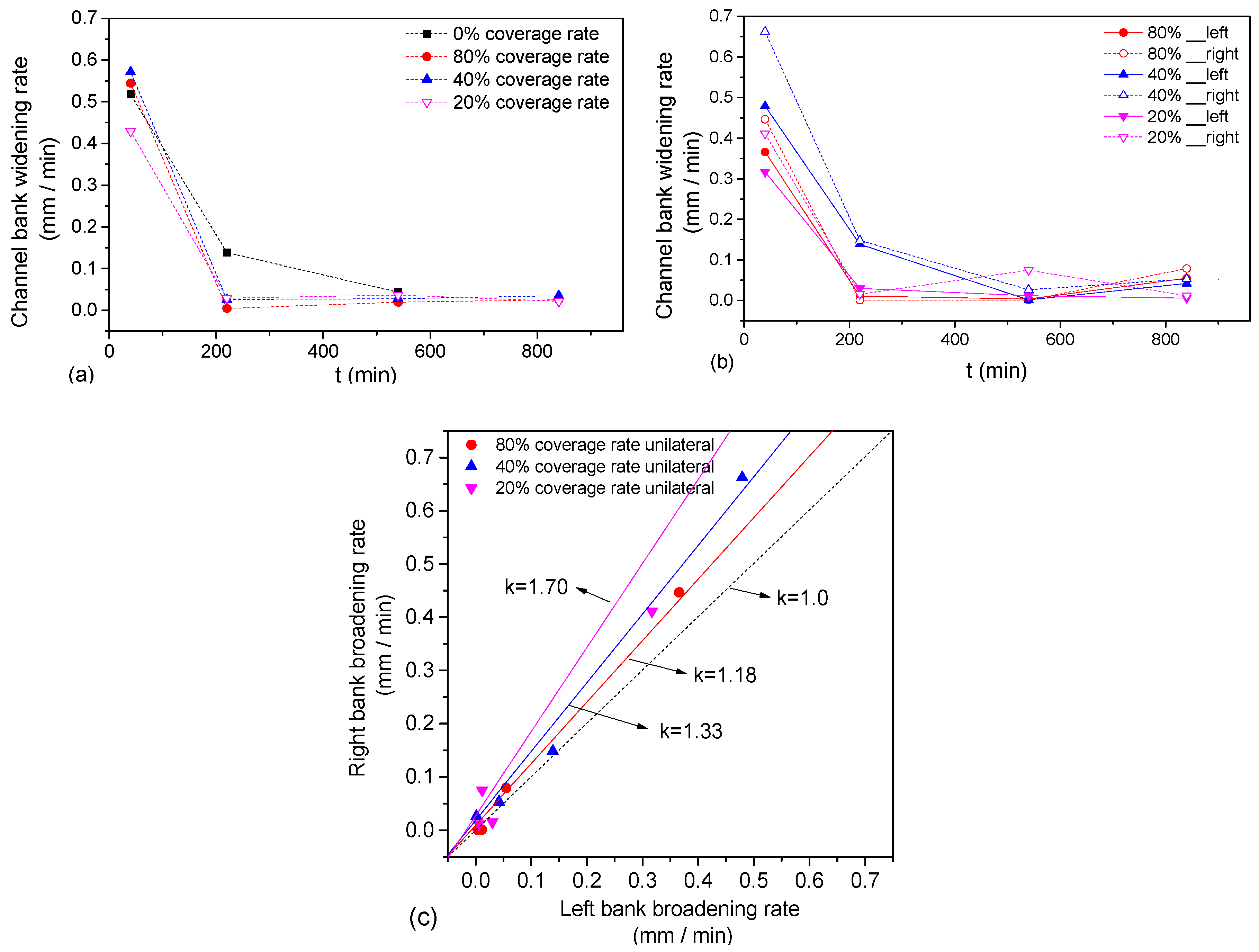
The average channel widening rate (
) during the whole period was calculated, and the widening rate of the river banks corresponding to different river channel evolution times was plotted for bilateral vegetation cover (
Figure 13a) and unilateral vegetation cover (
Figure 13b), in which the rate of river banks migration at corresponding points indicates average riparian erosion rates over a period of time. As seen in
Figure 13a, the average widening rate of the four sets of bilateral riparian vegetation cover decreased exponentially with time. For the narrow and deep channel with small initial width, the vegetation disturbance of the two groups with denser vegetation coverage (80%, 40%) was greater than that for sparse vegetation cover (20%) and no vegetation cover. With river channel widening, the disturbance of the water flow weakened and the inhibition of vegetation on the erosion of river banks enhanced, and the greater the vegetation coverage rate, the smaller the rate of channel bank migration. For the full time intervals, the average channel bank widening rate for no vegetation cover was 2.2-times that of the 80% vegetation coverage, 1.79-times that of the 40% vegetation coverage and 2.09-times that of the 20% vegetation coverage (
Table 3). The coverage of riparian vegetation effectively slowed the widening rate of the river bank. As seen in
Figure 13b, for unilateral vegetation cover, the average widening rate of the river banks was 40% > 80% > 20%.
The unilateral coverage of the riparian vegetation led to asymmetry of the river bank evolution. The average widening rate of the left bank corresponding to different times was taken as abscissa and the average widening rate of the right bank as the ordinate, as shown in
Figure 13c. We fitted the ratio curve of the average widening rate of the left and right banks. For 80% vegetation coverage, the average widening rate of the right bank was 1.18-times that of the left bank; and for 40%, the average widening rate of the right bank was 1.33-times that of the left bank. For 20%, the average widening rate of the right bank was 1.7-times that of the left bank (
Table 4). We found that the ratio for the average widening rate of the left and right banks decreased with increasing vegetation coverage.
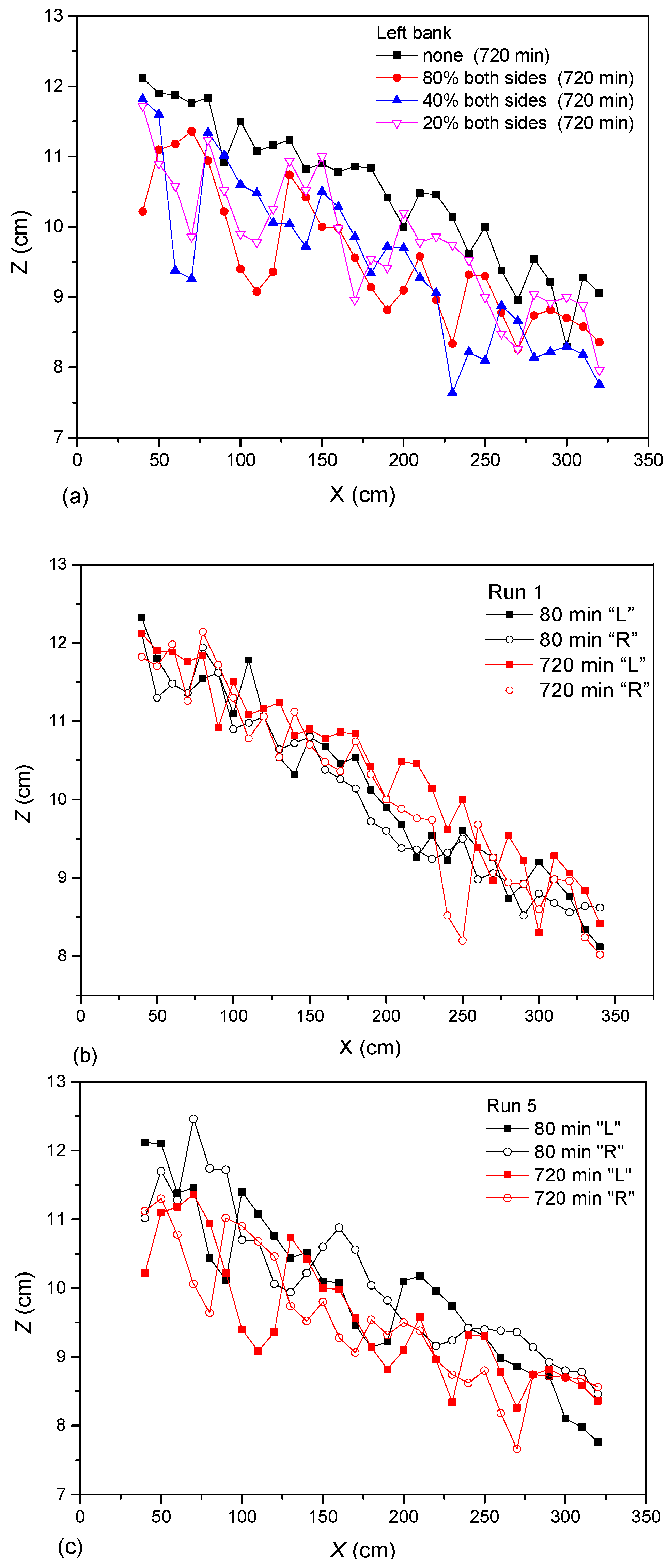
3.4. Near-Bank Bed Evolution
The effects of riparian vegetation cover on near-bank bed evolution are shown in
Figure 14a. The four groups of experiments (Runs 1, 5, 6 and 7) reached relative dynamic stability after running 720 min, so we collected the near-bank bed elevation under different riparian vegetation cover to make a comparison. It was obvious that the vegetation coverage had a strong influence on the near-bank elevation, leading to the increase in the trend of overall scour depth and bed surface fluctuation. The fluctuation amplitude of the bed elevation increased with the vegetation coverage. The influence of vegetation coverage was nonlinear. Smaller vegetation coverage was likely to form deep scour locally. Large-scale fluctuations in the near-bank bed elevation curves were related to the curvature radius of meander, and the distribution of pools along the elevation curves reflected the periodic distribution of the bend wavelength. However, the two were not in complete agreement with each other, and the pools were not only at the bend apex. In addition to large-scale fluctuations, there were also some small-scale fluctuations that had smaller fluctuation cycles, such as Run 6. The small pool at x = 1.7 m shows the history of river bend evolution. For no vegetation coverage (Run 1) in
Figure 14a, except for large-scale fluctuations, there were a large number of small periodic fluctuations along the channel bank. The alternating development of deep troughs and the shoals showed the historical process of the development of scouring and silting of channel sediment.
The longitudinal variation in near-bank bed elevations of Runs 1 and 5 are shown in
Figure 14b,c, respectively. We measured the near-bank elevation for four typical moments (80 min, 360 min, 720 min and 960 min). The initial time (80 min) and relative stability time (720 min) were used to investigate the influence of time on bed morphology.
Figure 14b,c show that the variation trends in the left and right bank profiles were basically the same. The main difference was a periodic phase lag existing between them. For Run 1 (
Figure 14b), under the action of flow and upstream sediment addition, the mean bed elevation did not change, and the undulation magnitude of bed elevation increased sharply. On the contrary, for Run 5 (
Figure 14c), the bed elevation had an obvious tendency of upward migration from 80 min–720 min, and the undulation of bed elevation became rougher, while the undulation magnitude remained the same with time. For the no vegetation condition, the bed topography changed weakly at the beginning, then gradually increased. For the river covered with riparian vegetation, because of the severe disturbance of the banks, the bed topography changed intensely, with the maturity of pools and point bars, and the flow and bed were still in the dynamic equilibrium state.
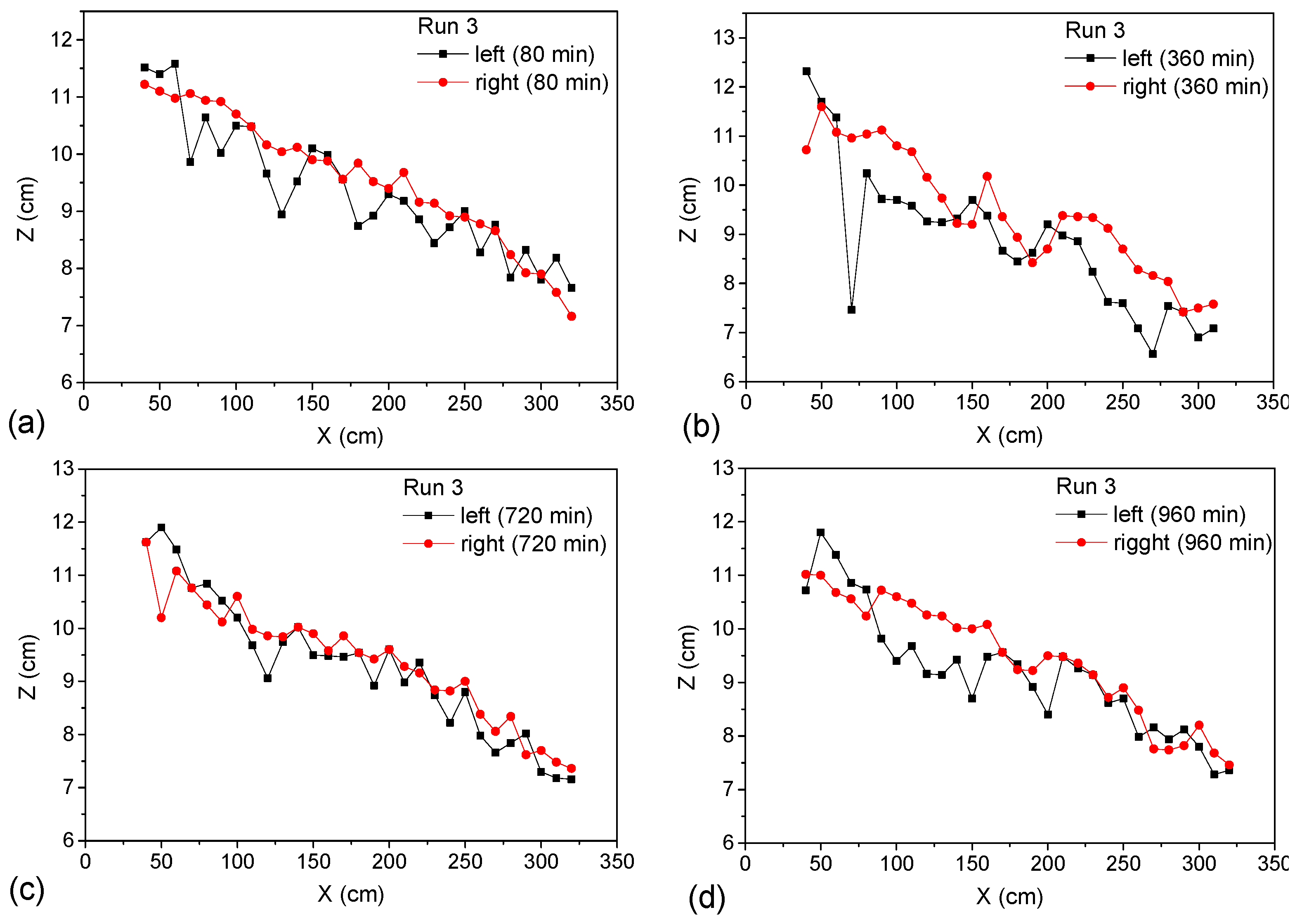
Taking the Run 3 case as an example, the influence of the unilateral vegetation cover on the near-bank bed elevations was analyzed, as shown in
Figure 15. Run 3 is covered with vegetation on the left bank and no vegetation on the right bank. The vegetation coverage of the river channel was different; the corresponding bank strength was different; and the corresponding near-bank elevation was also very different during the river channel evolution. As can be seen from
Figure 15, the near-bank elevations of the left bank were obviously lower than those of the right bank. This may be because, although the riparian vegetation increased the riverbank erosion resistance, it also increased the instability of flow and the disturbance to the river bank. In the initial stage, the intense disturbance action was mainly reflected in many small-scale undulations along the river bank. With the constant erosion of the left bank, these riverbed undulations propagated to the center of the river channel; thus, the cross-section flow structure and the bed morphology near the right bank were disturbed. At 360 min, alternately distributed large-scale point bars and pools developed on the left, while there were also unstable large-scale pools appearing on the right. As the evolution continued, the bed morphology constantly changed and adapted to the flow structure. At 720 min, large-scale point bars and pools disappeared and converted to small-scale bed undulation. The development of left and right banks was more uniform this time. Subsequently, due to the imbalance of riparian characteristics, the river bed continued to adjust and develop. For this unilateral vegetation-covered channel, the imbalance in riparian characteristics led to the continuous disequilibrium evolution of bed morphology and to the unstable evolution of the river dynamic axis.
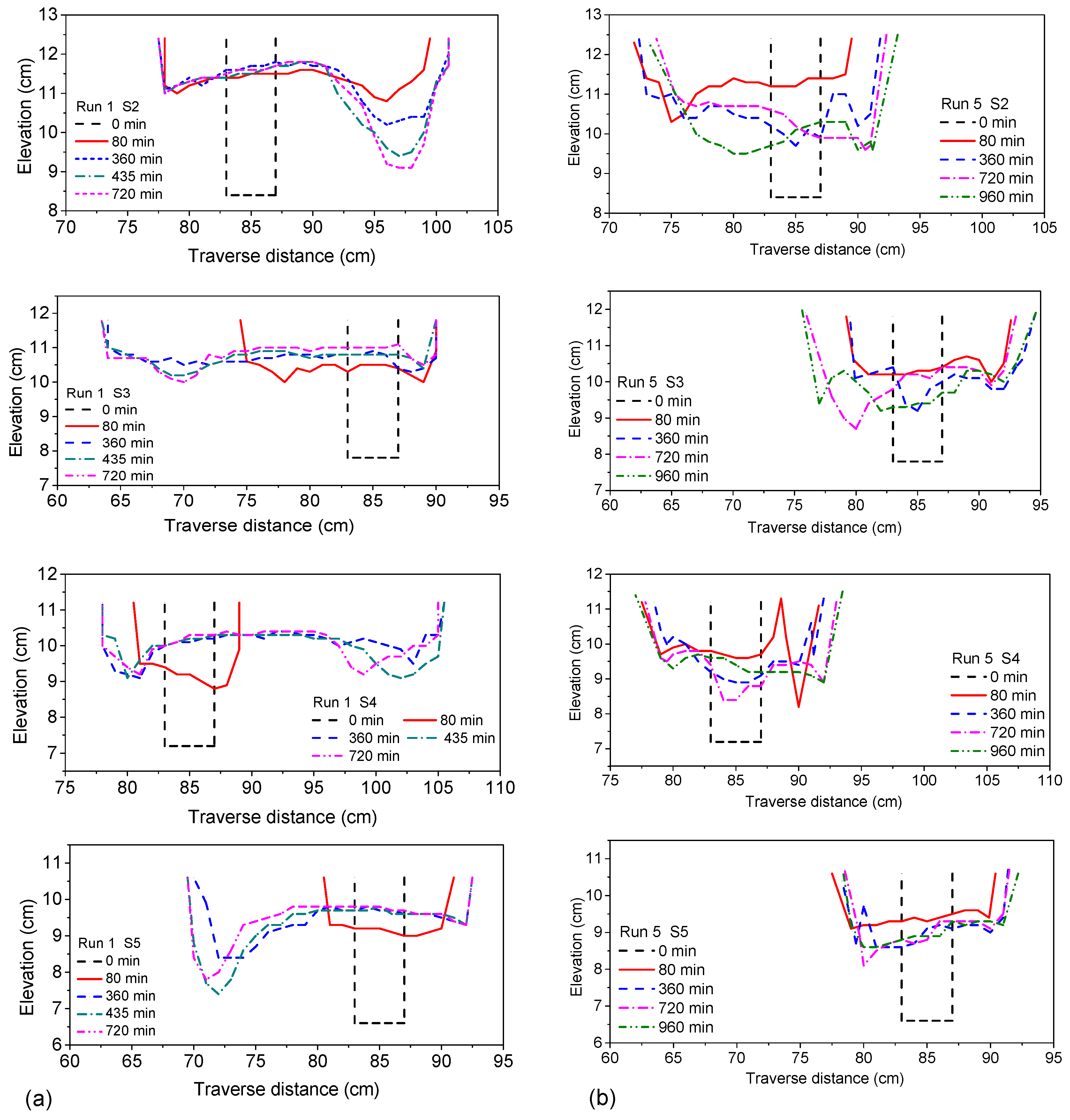
3.5. Stability of Meanders
The cross-section of the river bed mainly reflected the process of erosion and deposition of the riverbed and also reflected its widening process. Four runs (1, 3, 5 and 6) under different conditions were selected for comparative analysis (
Figure 16a–d, respectively). The focused time frame was from 0 min to the final stable time. We chose the section of the second to fifth as the typical sections to study ignoring the impact of entry and exit, and the corresponding transverse section of x = 1.0 m (S2), x = 1.5 m (S3), x = 2.0 m (S4) and x = 2.5 m (S5). The cross-section was chosen to approximate the half wavelength of the meandering channel (Run 1), and the spacing was about 0.5 m.
For Run 1, observations showed that the riverbed overall expanded and silted up in the initial state, then along the concave bank, the scour deepened, obviously forming the main channel. The main channel alternatingly swung from the left bank to the right bank, forming a stable meandering channel. For Run 3, unilateral vegetation cover of its riverbanks, the channel was in the stage of adjustment in the initial 80 min, rapidly widening with siltation uplifting, the same as in Run 1. With the continued evolution of the river channel, unlike Run 1, Run 3 showed an obvious unilateral scour along the vegetation covered side, and the main channel swung to the left. Especially after x ≥ 2.0 m, there was a special phenomenon of one side shoal and the other side a deep trough.
In the period of continuous extension of the river channel, Run 3 showed some unique processes, which were different from the phenomena of many previous river evolution experiments. Compared with the continuous silting up of the riverbed of Run 1, the riverbed of Run 3 showed first aggradation and then degradation. The main reason for this phenomenon was the unilateral perturbation, which led to the asynchrony of the two river bank sides, and the water flow showed constant disturbance. The riverbed was always in the process of constant siltation and erosion. This strong asynchronous perturbation was also shown in the cross-section of the riverbed, showing multi-groups of alternating distributions of secondary deep troughs and shoals. Seen from
Figure 16c, although the left bank was constantly deeply scouring, the lateral extension was equal to or less than the right bank, especially evident in the downstream x ≥ 2.0 m. Finally, a secondary channel was formed at x = 2.0 m, with a new first deep trough and multiple secondary deep troughs. At the same time, the initial first deep trough was abandoned, and the left bank point bar had become the new river bank of the secondary channel. Vegetation played an obvious role in riparian protection.
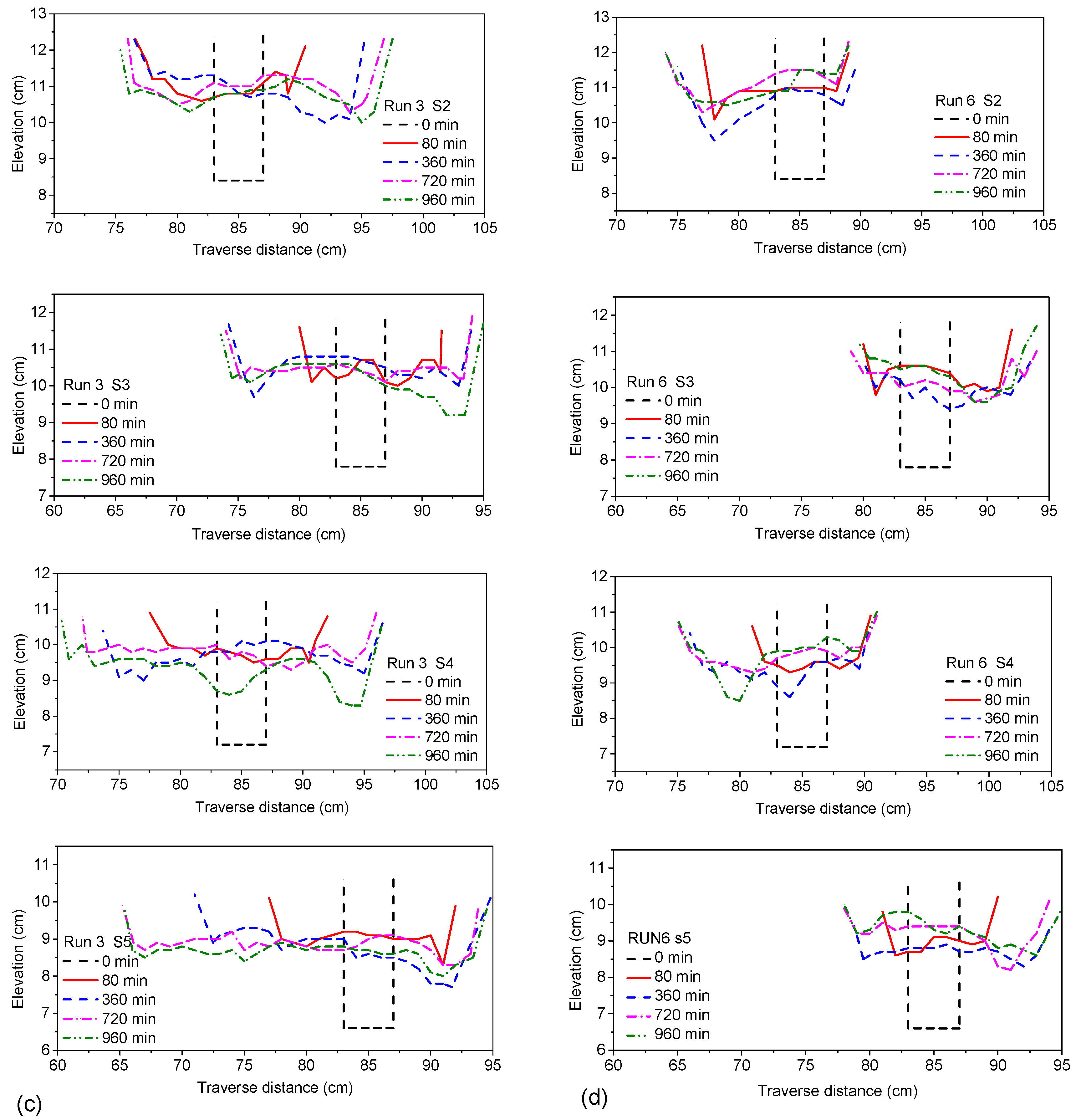
For Runs 5 and 6, with bilateral vegetation cover on the riverbanks, after the rapidly widened and siltation uplifted phenomena (t < 80 min), most of its river channel reaches showed the phenomenon of first brushing deep and then silting up. This showed that the bilateral vegetation cover on the riverbank effectively controlled the continuous deep brushing of the riverbed. Looking at
Figure 16d, we found that a continuous alternate shoal along the riverbank finally formed, showing a stable duplex bed forming with less secondary deep troughs. S2, S3 and S4 present the phenomenon of narrowing and deposition on both bank sides. Therefore, the channel with bilateral vegetation cover had a relatively stable bed morphology that could effectively restrain the continuous riverbank erosion process compared to the channel with unilateral vegetation cover. The formed stable width-depth radio was lower than under no or unilateral vegetation cover. Therefore, a narrow and deep channel more easily formed for the channel with bilateral vegetation cover during the flood period.
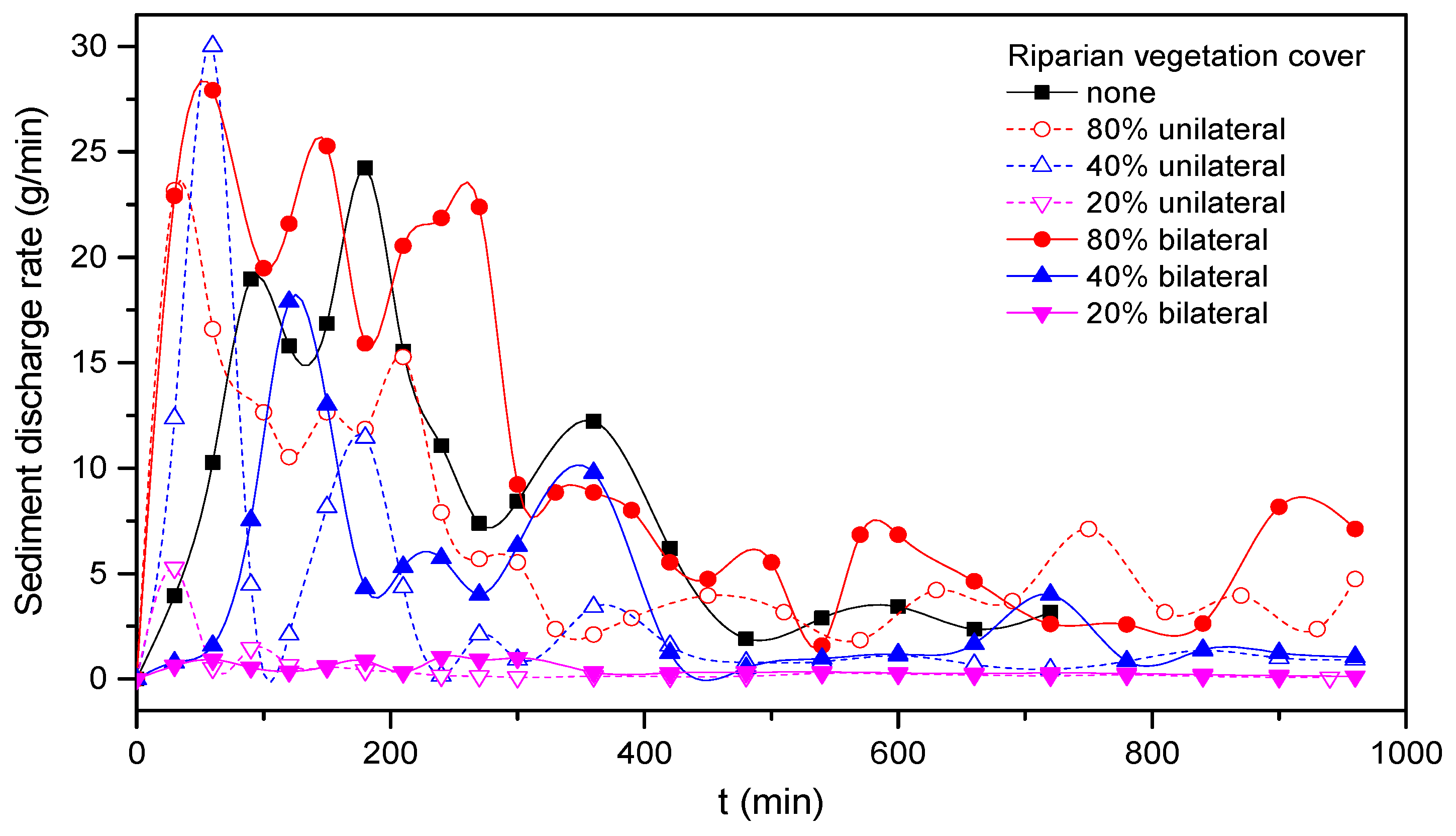
3.6. Sediment Transport Rate
As shown in
Figure 17, under the condition of initial excavation and a small-scale straight path, the fluctuation in the channel sediment transport rate decreased with the adjustment of water flow and channel, and finally reached a relatively stable state. In terms of trough measurements, we found that there was a common rule for different vegetation coverage rates: the sediment transport rate of unilateral vegetation cover was greater than that of bilateral vegetation cover at the initial stage, but was lower at the stable stage. In Run 3, due to the strong unilateral disturbance and the instability of flow structure, the sediment transport was strong at the initial stage, and the gradient and amplitude of the sediment transport rate curve were larger than for runs without vegetation. As for Run 6, because of the mutual inhibition between the two sides, the sediment transport rate was smaller than Run 3 and then had an increasing trend at the initial stage.
For the experimental scale of this small river channel, the maximum allowable dry fine sand passing with the flow was about 1.5 g/min for recovering the bedload sediment deficit, whereas the suspended load was derived from bank erosion as the channel progressed [
32]. For the seven groups of experiments, the final relatively stable sediment transport rates are shown in
Table 5. For 20% and 40% vegetation coverage rates under unilateral and bilateral riverbank vegetation coverage, the average sediment transport rates were significantly lower than without vegetation cover. When the flow passed through the vegetation zone, the existence of riparian plants increased the local hydraulic roughness, decreased the flow velocity and increased the sedimentation, and the vegetation zone effectively inhibited the lateral scour of the river channel. However, for the 80% vegetation coverage rate, the sediment transport rate increased.
The reasons for these findings are as follows. Although the planted riparian vegetation played a role in a flexible bank to a certain extent, when the planting of vegetation was very dense, vegetation revetment became a hard constraint, similar to a riparian vegetation wall. Although riparian vegetation increased riparian intensity to the greatest extent, it increased the disturbance of the river channel. Although the lateral scour was suppressed, the vertical scour of the river channel increased. It can be clearly seen from the cross-section diagram of Run 5 in
Section 3.2. At the same time, because of the inhibition of the vegetation on the lateral scour, the width of the channel was relatively small, the main stream flow velocity relatively increased and the sediment carrying capacity of the flow was enhanced. Therefore, the influence of riparian vegetation coverage on river channel sediment transport rate can be positive and negative and has a strong relationship with the density of vegetation. A certain amount of vegetation cover can effectively reduce the sediment transport rate.
From different riparian vegetation coverage rates, we found that sediment transport increased with increasing coverage rate, and good consistency was maintained between unilateral and bilateral banks. The influence of the degree of coverage on the sediment transport rate was not a simple linear relationship; the amount of sediment transported corresponding to 20% vegetation coverage was much lower than that of the 40% and 80% vegetation coverage. This is because, under 20% vegetation coverage, the local disturbance of riparian vegetation to water flow was much smaller than that of the other two conditions, especially in the initial stage of river channel evolution. However, this had a good reinforcement effect on the river bank, reducing the erosion.




















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}