1. Introduction
The coastline of Lake Ontario, the furthest downstream of the Laurentian Great Lakes, is valued for its water resources and ecological services, but has a history of human disturbance. Large populations live in cities on the shores of the lake. They use water within one to two kilometers of the shoreline as a source of drinking water and as a receiver of wastewater. On the Canadian shoreline between the cities of St. Catharines and Oshawa, encompassing 160 kilometers of coastline, there are 12 wastewater plants. The plants service a collective population in excess of 5 million people. The proximity of so many people to a naturally oligotrophic water body [
1] poses a threat to water quality along the coastal margin where most land-based runoff and discharges first mix with the lake.
Surface runoff that is discharged from tributaries and storm water create mixing zones at the land–water interface frequently enriched in nutrients, major ions, suspended solids and fecal pollution indicators [
2,
3,
4,
5]. A characteristic feature of this coastline is watershed tributary inputs where an appreciable portion of the drainage area is urbanized. Treated wastewater, discharged at depths typically <10 m, create localized plumes with elevated nutrient levels. The phosphorus discharged from water pollution control plant (WPCP) outfalls has a high proportion of the phosphorus in a dissolved form, much of it as soluble reactive phosphorus (SRP), as contrasted with phosphorus attached to particles which typically dominates non-point sources such as tributaries and storm water [
6,
7]. The implication is that phosphorus loaded from a WPCP outfall has a more immediate and higher unit mass potential to stimulate primary productivity compared with tributary runoff. Considering the two most obvious sources of phosphorus loading at the shore of the lake, WPCPs and tributaries, a Lake Ontario phosphorus loading analysis in 2008 [
8] indicated that the 54 tributaries between St. Catharines and Oshawa loaded 494 mt of phosphorus compared with 607 mt from WPCPs over the same shoreline.
Eutrophication problems in western Lake Ontario are centered on the overabundance of the green algae
Cladophora which blankets much of the shallow lakebed and periodically fouls shorelines [
9,
10,
11,
12]. There is growing concern that regional nutrient loading is causing or contributing to the overabundance of
Cladophora [
9,
10,
11,
12,
13]. Phytoplankton levels in the nearshore are generally low. In recent years there have not been water-column algae blooms in this region of the lake. Lake-wide spring total phosphorus (TP) concentrations when the lake is mixed have declined over the last 35 years subsequent to phosphorus controls in the 1970s, albeit more slowly in the past 20 years. Recent levels range from 5 to 7 μg/L indicative of oligotrophic conditions [
14]. Studies of nutrient patterns in the nearshore of Lake Ontario confirm that nutrient enrichment occurs widely, particularly in shallow water, but that the spatial and temporal extents vary among regions [
3,
5,
15].
A key factor in the resurgence in
Cladophora, previously a problem in the 1960–1980s when the lake was more nutrient enriched, is the improved light climate of the lake following the invasion of dreissenid mussels [
16] in about 1990 [
17]. Kuczynski et al. [
18] have convincingly demonstrated that there has indeed been a resurgence in
Cladophora which can be directly attributed to the changing light climate. These studies have concluded that the growth potential for
Cladophora has increased markedly compared with the period prior to the deepening photic zone.
The only tool available for abatement of
Cladophora in Lake Ontario is the control of phosphorus inputs [
18]. Based on the most recent published estimate, the single largest phosphorus input to the lake is upstream from Lake Erie, accounting for 27% of the annual loading [
19]. A hurdle to advancing phosphorus management to abate
Cladophora is clarifying the mechanisms and spatial scales of phosphorus loading into Lake Ontario that drive the growth of
Cladophora.
The benthification (see [
20]) of the nearshore of Lake Ontario and other parts of the Great Lakes following the invasion of dreissenid mussels [
21,
22] has also raised questions on nutrient transfers at the lakebed [
20,
23,
24,
25]. The critical role of external nutrient loading at the lake-wide scale is less obvious at a regional scale. Here, the physical and biological transfers to the inshore from the offshore occur concurrently with local-area loading. A pivotal question in Lake Ontario at this time is the role of the flux of nutrients into the nearshore from the offshore routed through the lakebed by filter-feeding of dreissenid mussels.
This article examines the distribution of
Cladophora over urban shoreline in Lake Ontario to identify associations with nutrient sources. This work stems from a 2008 study [
9] conducted on a lake-wide scale which found that the nuisance biomass of
Cladophora was limited to urban areas, and correlated to area nutrient enrichment. The finding suggested that nutrient management to abate
Cladophora may best be focused on area nutrient sources. Further studies in 2012 and 2013 examined the generality of the 2008 findings across a regional nutrient gradient and further examined how nutrient enrichment in an urban setting promotes the proliferation of
Cladophora. However, unlike in 2008, the distribution of
Cladophora was broadly similar among regions that varied in degree of urban development. The four study areas, three of which were adjacent to urbanized shoreline and a fourth with little development, had similar, high biomass levels [
10]. Three possibilities were considered to explain this unexpected finding: (i) that internal nutrient loading linked to refocusing of phosphorus from other parts of the lake by filter feeding of dreissenid mussels accounted for a portion of the nutrient supply for
Cladophora; (ii) differences in light regime among areas and, degree of light limitation affected biomass production, and, (iii) the
Cladophora sampling methods could not adequately differentiate the abundance of
Cladophora among areas [
10].
Embedded within this regional study was a more detailed examination of environmental conditions over one of the urban study areas. This examination, reported here, considered how typical local-scale ranges of nutrient and physical conditions may affect the growth of Cladophora, to contrast and compare with the coarse grain analysis at the regional scale. The possible reasons for the unexpected findings at the regional scale noted above are also examined further.
This article examines the nutrient regime, and water quality, over the shoreline which features discharge from a WPCP, runoff from urban tributaries and stormwater outfalls. Measurements of dissolved nutrients directly above the lakebed are used to represent the nutrient regime in proximity of Cladophora. The heavy colonization of the lakebed by dreissenid mussels is considered as a possible correlate with water quality and Cladophora distribution. Finally, the distribution of Cladophora in relation to potential sources of nutrients is examined.
4. Discussion
4.1. Urban Influences on Nearshore Water Quality
Macro-scale gradients, on the order of 10s to 100s of meters, in nutrients and other water quality features were evident from the field sensor measurements. Locations near the WPCP diffuser, a tributary mouth and, in close proximity to the shoreline were most influenced by the adjacent shoreline. Nutrients, suspended solids, major ions and chlorophyll
a were elevated and co-varied broadly in areas affected by runoff and discharges, similar to other areas of developed Great Lakes shoreline [
3,
5].
A mixing area near the WPCP diffuser was usually the largest spatial feature evident from the field sensor measurements. The orientation was approximately parallel to the shore, consistent with the pattern in current directions (
Table 4). Leon et al. [
36], using a combination of field and modeling studies conducted in 2007 and 2008, similarly reported that the mixing zone associated with the WPCP diffuser was the most apparent and prevalent water quality feature of the area. Field nitrate was the strongest overall tracer as inferred from correlations between field measurements and lab-measured TP and DP concentrations (
Table 3). Previous studies have used nitrate to infer the mixing areas associated with discharge from the Duffins Creek WPCP [
13,
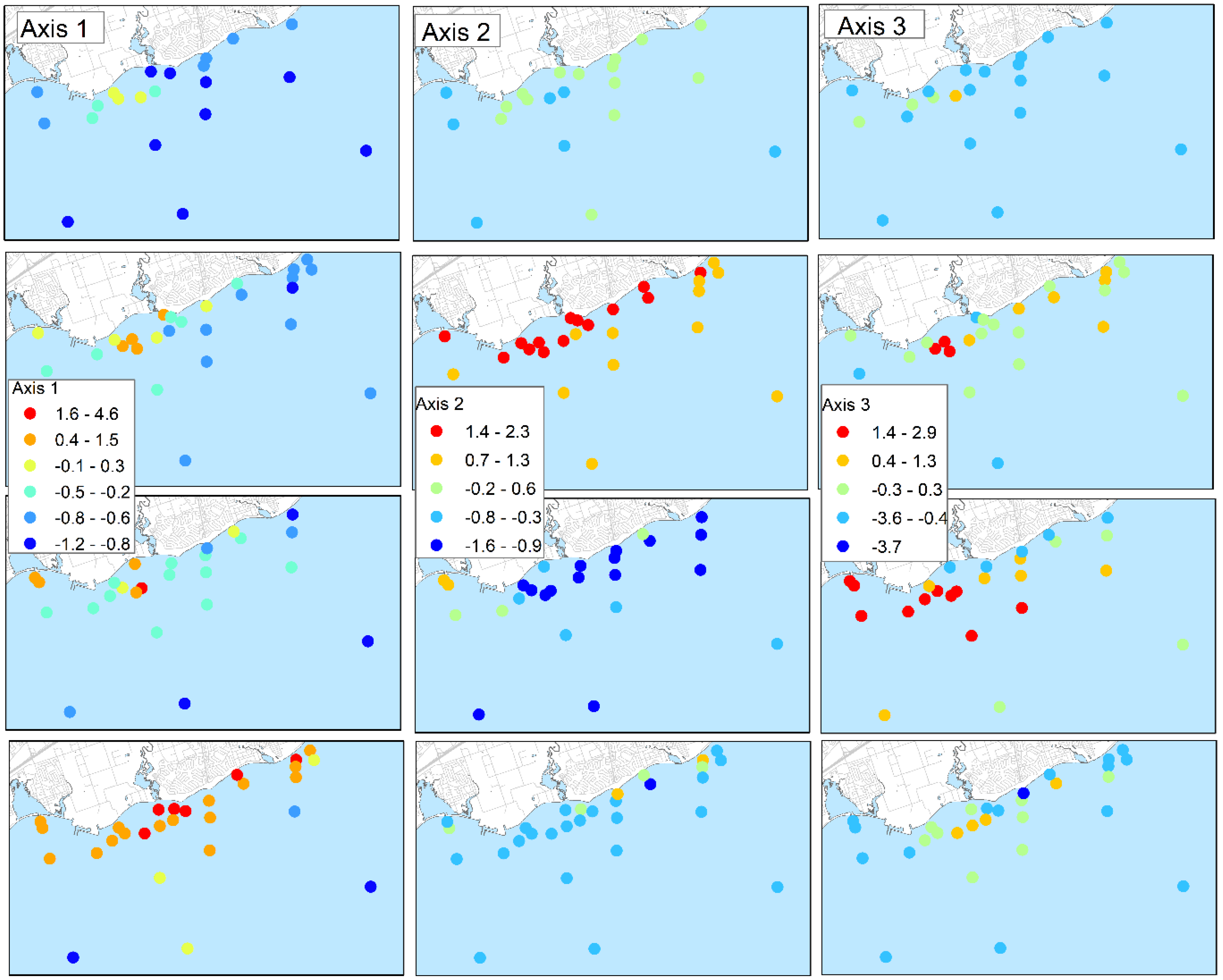
37]. However, the dimensions of the mixing areas at the lake surface could not be delineated using the field sensor data because of the interacting influence of other area loading sources. This is evident from the PCA analysis of lab-based water quality data. The third axis separated locations based on the influence of the discharge from the WPCP, yet nitrate + nitrite did not correlate strongly with the axis as contrasted with ammonia + ammonium, TP and DP. This suggests that enrichment of nitrate + nitrite is less specific to loading from the outfall than ammonia + ammonium. The field conductivity and CDOM fluorescence also depict mixing areas in the vicinity of the WPCP diffuser but the spatial patterns are also influenced by other discharges along the shoreline.
The limit of navigation, and data collection, was the 3-m depth contour which constrained the ability to resolve shoreside (<2 m) conditions where tributary and stormwater discharges are most strongly focused and often dispersed directly adjacent to the shoreline by alongshore flow [
2,
38]. At times, conductivity and CDOM fluorescence were elevated at points along the shoreward edge of the study area. Field sensor data is difficult to interpret when lakeward signals are not well captured. Nevertheless, it is clear that phosphorus concentrations were elevated along the immediate shoreline due to runoff into the lake.
Water column productivity was low, as inferred from chlorophyll
a. The low concentrations of chlorophyll
a and the absence of visible blooms suggest limited effects on phytoplankton from area discharges and runoff. This does not mean that phytoplankton growth was not affected, only that lake mixing is sufficient to disperse added phytoplankton production to lake background levels. Water column TP concentrations measured near the lake surface also suggest broadly oligotrophic [
14] conditions, however, the higher concentrations in the vicinity of the WPCP diffuser and at shallow depths strongly indicate areas of enrichment (
Figure 2).
Temporal changes in water quality due to both area and lake-wide influences were evident throughout the study. Site scores on the third PCA axis noted above varied among sampling events, at times suggesting limited influence of the WPCP discharge, as viewed at the lake surface, and at other times, such as in October 2012, suggesting more widely distributed effects on water quality. Lake-wide seasonal changes due to phytoplankton and lake warming were also evident from the PCA analysis. Chlorophyll
a is positively associated and nitrate + nitrite and silica are negatively associated with PCA axis 2, consistent with the summer drawdown of these nutrients by phytoplankton [
39,
40]. The association with turbidity likely stems from seasonal development of whiting in Lake Ontario which results in increased turbidity as calcium carbonate precipitates [
41].
Water quality in 2012 and 2013 is broadly similar to that described using a comparable sampling approach in 2008 [
2]. Other earlier studies have also described water quality in the area [
13,
15,
36] which lies within the eastern extent of the footprint of the Greater Toronto Area (GTA) on the north shore of the lake. A west to east gradient of decreasing nutrient richness associated with the urban development of western Lake Ontario and the distribution of large rivers has been evident in studies since the mid-1970s. Lake-wide analysis of nutrient distribution in 1981 and 1982 [
42] indicates a declining gradient in TP on the north shore away from the GTA. More recently, in 2008, higher TP concentrations were reported in the GTA than further east on the north shore [
3], similar to findings in the regional component of the present study which found a decreasing gradient in TP and DP concentrations on the north shore extending from the City of Toronto to less developed shoreline further east [
10]. Phosphorus levels averaged over the Pickering-Ajax study area were intermediate to the end points of the gradient [
10].
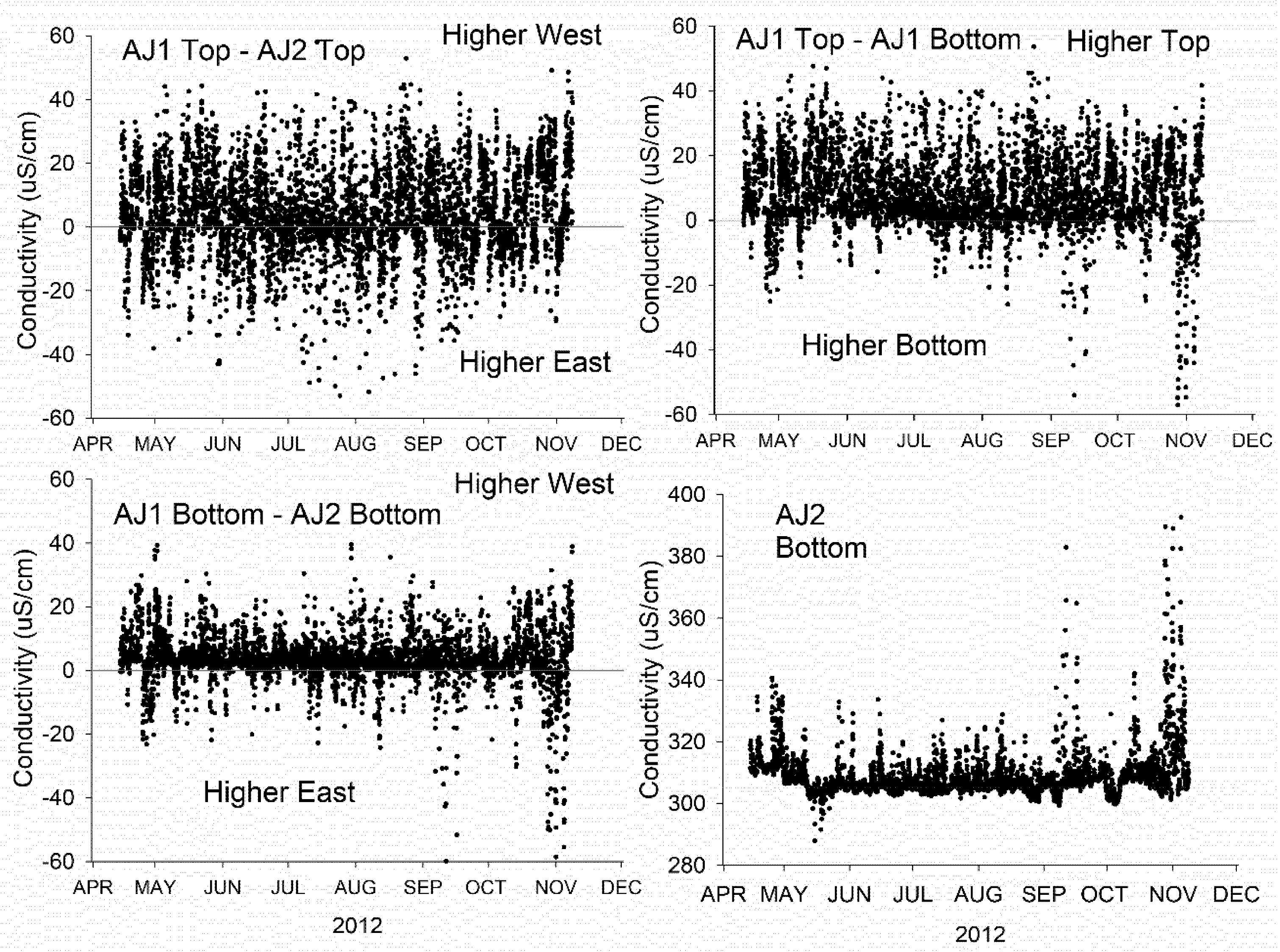
Limited chemistry sampling was conducted through the water column, making it difficult to discern the variability due to depth-variable mixing of discharges. Of concern is that the discharge at the WPCP diffuser results in mixing zones within the water column that are not adequately represented by the near surface or lakebed sampling reported here. Lake-deployed conductivity sensors indicated that conductivity was frequently elevated near the lake surface as compared with near the lakebed in the area of the diffuser. This suggests that the discharge is often buoyant and nutrient enrichment should often be evident at the lake surface, albeit at levels below maximal levels within the water column adjacent.
The Duffins WPCP is a large continuous source of bio-available phosphorus as noted in earlier studies [
13,
36], however, placing it in context with other sources requires data which is not fully available. Tributary load estimates are not available for 2012 and 2013. Makarewicz et al. [
8] reported a loading of 14.8 mT to the study area in 2008 from monitoring of Duffins Creek and Caruthers Creek which suggests that the WPCP load of phosphorus far exceeds the tributary load, even without consideration of the relative bioavailability of phosphorus. However, Booty et al. [
43] using different load estimation techniques reported higher and wide-ranging phosphorus loads from Duffins Creek. Estimates ranged from 13 to 17, 57 to 73 and 69 to 92 mT in 2007, 2008 and 2009, respectively. Load estimates from shoreline stormwater are not available but of interest considering the proximity to the shallow water where Cladophora is abundant. Loading of phosphorus to Lake Ontario varies among years due to weather patterns affecting tributary loading (e.g., [
43]) as well as varying municipal loading. For example, the discharge of phosphorus from the Duffins Creek WPCP in 2008 was nearly double that of 2012 and 2013; annual phosphorus loading was 90.2 mT in 2008 [
8] compared with 42.8 and 43.8 mT in 2012 and 2013, respectively.
A highly uncertain element of the phosphorus budget relates to the transport of nutrients across the offshore boundary into the study area driven by onshore circulation. At first look, the influx of offshore water will lower water column concentrations because of the low TP concentrations in the offshore of the lake, likely diluting the phosphorus supply in the nearshore water column. However, this overlooks the capacity of filter feeding by
Dreissena on the nearshore lakebed [
22] to extract phytoplankton, and with it phosphorus, from the water column as offshore water circulates inshore. The current and temperature data collected in 2012 demonstrates that the nearshore water column is loaded with offshore water during periods of onshore circulation. Phosphorus extracted from offshore water and sequestered on the nearshore lakebed represents an uncertain phosphorus load that needs to be considered in a phosphorus budget for the study area.
4.2. Nearbed Concentrations of Dissolved Nutrients to Detect Enrichment Affecting Benthic Algae
Concentrations of SRP, DP and TIN measured at the lakebed suggested enrichment from area sources, however, the spatial associations were weak and often uncertain. Concentrations were measured approximately 10 cm above the lakebed, which in principal, reduces the uncertainty due to assimilation and mixing within the water column and yields a more direct measure of nutrient levels in proximity to the growing algae. However, inferences on external sources are constrained by the fact that internal flux of nutrients at the lakebed may obscure spatial associations between external loading and nearbed concentrations.
Taken together, the results from seven sampling events in 2012 and 2013 provide mixed evidence of local-area enrichment of SRP. Higher nearbed concentrations of SRP were detected near the WPCP diffuser and over the alongshore axis away from the outfall during the August and October 2012 sampling events. In contrast, at other times there was no obvious pattern other than high relative values at a site near the diffuser. There was little consistent pattern in SRP levels along the shoreline at the shallowest sites (<5 m depth) where concentrations were generally lower than adjacent locations in deeper water. It is likely that biological drawdown lowers concentrations at times such that enrichment is less evident. In addition, mixing plumes are sometimes buoyant, reducing contact time with the lakebed as suggested in the area of the WPCP diffuser by the conductivity sensor data. Concentrations were often highest at the deepest sites where biological utilization of SRP was likely reduced compared with better illuminated shallower sites where Cladophora was abundant.
Nearbed SRP concentrations were mostly <2 µg/L, with area-wide average values among sampling events ranging from 0.8 to 2.9 µg/L, with the exception of October 2012. There are few comparable lakebed measurements of SRP for nearshore Lake Ontario. Ozersky et al. [
44] reported SRP concentrations ranging from 0.9 to 2.8, averaging 1.9 µg/L, in samples collected 20 cm above the lakebed in June and July of 2006. Spring-time water column concentrations in the lake offshore when the lake is mixed provide an indirect reference point. Dove and Chapra [
14] report concentrations of 1.4 and 2.0 µg/L for the spring of 2012 and 2013, respectively, with mean spring concentrations since 2010 ranging from 1.4 to 3.1 µg/L.
Concentrations of DP are expected to be proportionally less affected by biological uptake than SRP and may better depict enrichment patterns. The linear regressions predicting SRP from DP (
Table 7) also suggest that DP should also be an indicator of bio-available phosphorus. At times, nearbed DP concentrations were higher at sites in proximity to the WPCP diffuser and the mouth of Duffins Creek. In May 2012, concentrations were higher at sites <10-m in depth compared with sites 20 m in depth, indicating enrichment in the shallows. Concentrations were higher on the east compared with the west shoreline (
Table 5), suggesting loading along the eastern shoreline or alongshore transport of discharge from Duffins Creek. In August 2012, DP was higher in the area of the diffuser, a pattern also seen in SRP. A stronger influence of the WPCP on DP was seen in October 2012 with higher levels to the west of the diffuser. In contrast, the results for June 2012 and June and July 2013 suggest little spatial pattern other than a possible source of enrichment on the eastern shoreline. Collectively, the results suggest periodic enrichment of the lakebed due to the wastewater discharge and shoreline runoff. The limited increase in DP concentration with depth compared with SRP is consistent with more limited bioavailability and biological draw down of DP compared with SRP.
Nearbed concentrations of inorganic nitrogen also provide evidence of the influence of WPCP discharge on the lakebed nutrient regime. Concentrations were higher near the WPCP diffuser at times, most notably in October 2012. External loading of TIN should be more evident compared to SRP because of the proportionally lower biological utilization of the total supply. Unlike phosphorus, TIN concentrations are thought to remain above growth-limiting levels for phytoplankton (e.g., [
45]) and
Cladophora [
9,
18]. Linear regressions predicting SRP and TP from TIN were significant for three and two of seven sampling events, respectively (
Table 7), suggesting that lakebed TIN may be a predicator of external loading of phosphorus, such as in August 2012, but that this is not consistently the case. Offshore loading of TIN from the hypolimnion may mimic or mask external loading during mid to late summer when TIN concentration gradients develop through the water column. During the summer, TIN becomes depleted in the epilimnion [
2,
39,
40] and consequently onshore transport of hypolimnetic water results in gradients of TIN in the nearshore both with depth within the water column and with proximity to shore. The upwelling over the study area in 2012 (
Figure 6) and reported in other studies [
2,
46] suggests that variability in nearbed TIN concentrations is to be anticipated irrespective of local-area loading.
4.3. Interpreting the Distribution of Cladophora
Benthic blooms of
Cladophora, suggesting strong growth under nutrient enriched conditions, were evident over the study area. The shallow lakebed was widely covered by
Cladophora. Biomass of
Cladophora exceeded the criteria of 30 and 50 g/m
2, used to infer nuisance conditions [
47], in some samples. However, there was no clear or consistent difference in the amounts of
Cladophora among sub-areas of the shoreline.
Unlike biomass, concentrations of phosphorus in
Cladophora, an indicator of phosphorus sufficiency [
48], varied over the study area with higher levels near the WPCP diffuser. The higher Qp values indicate enrichment and algal growth under less phosphorus-limited conditions in the absence of confounding effects of other growth-limiting factors. Growth rates are predicted to increase strongly as Qp rises to 0.1%, less rapidly to 0.2%, reaching an asymptote after which the growth rate is unresponsive to increasing cellular phosphorus [
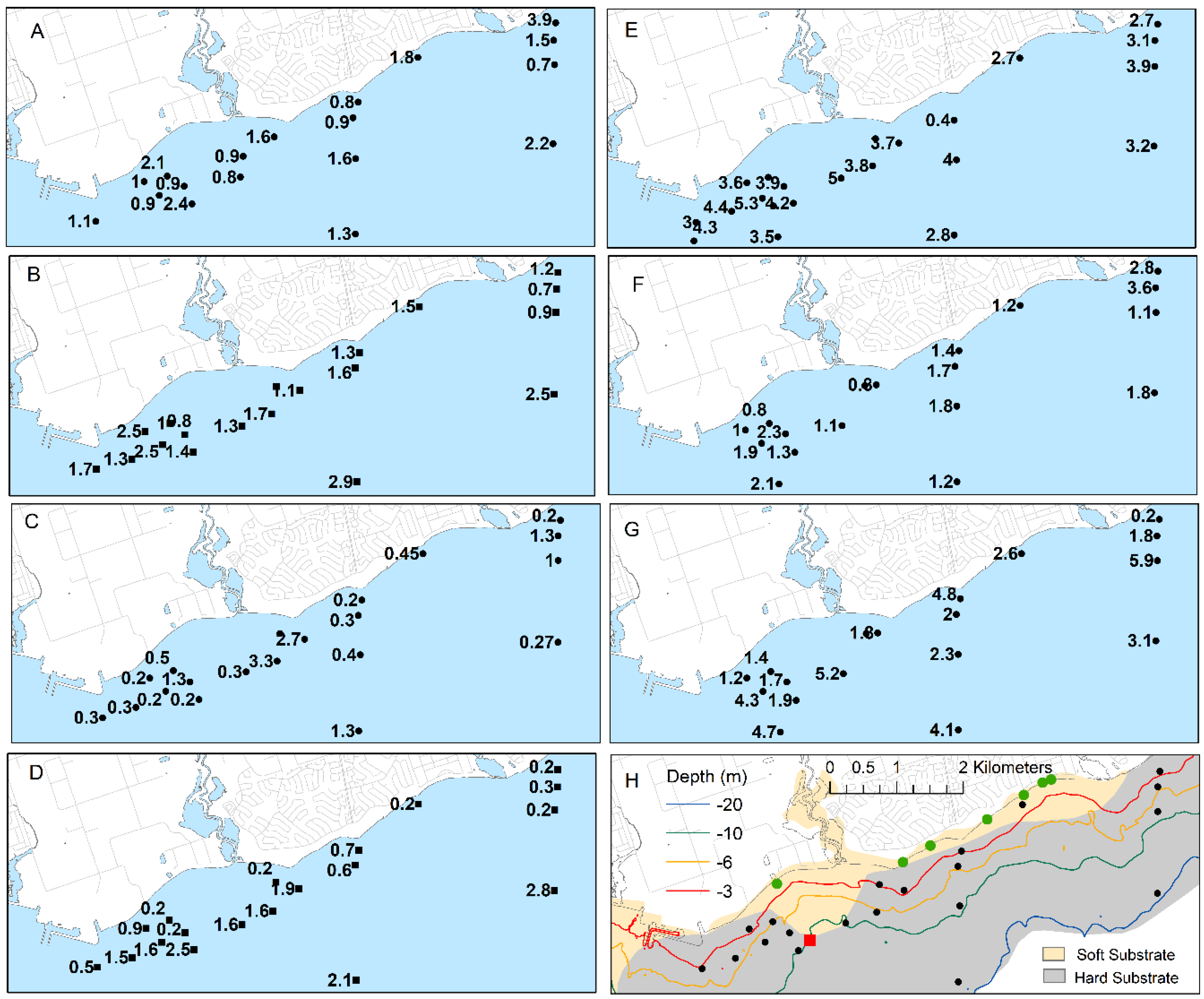
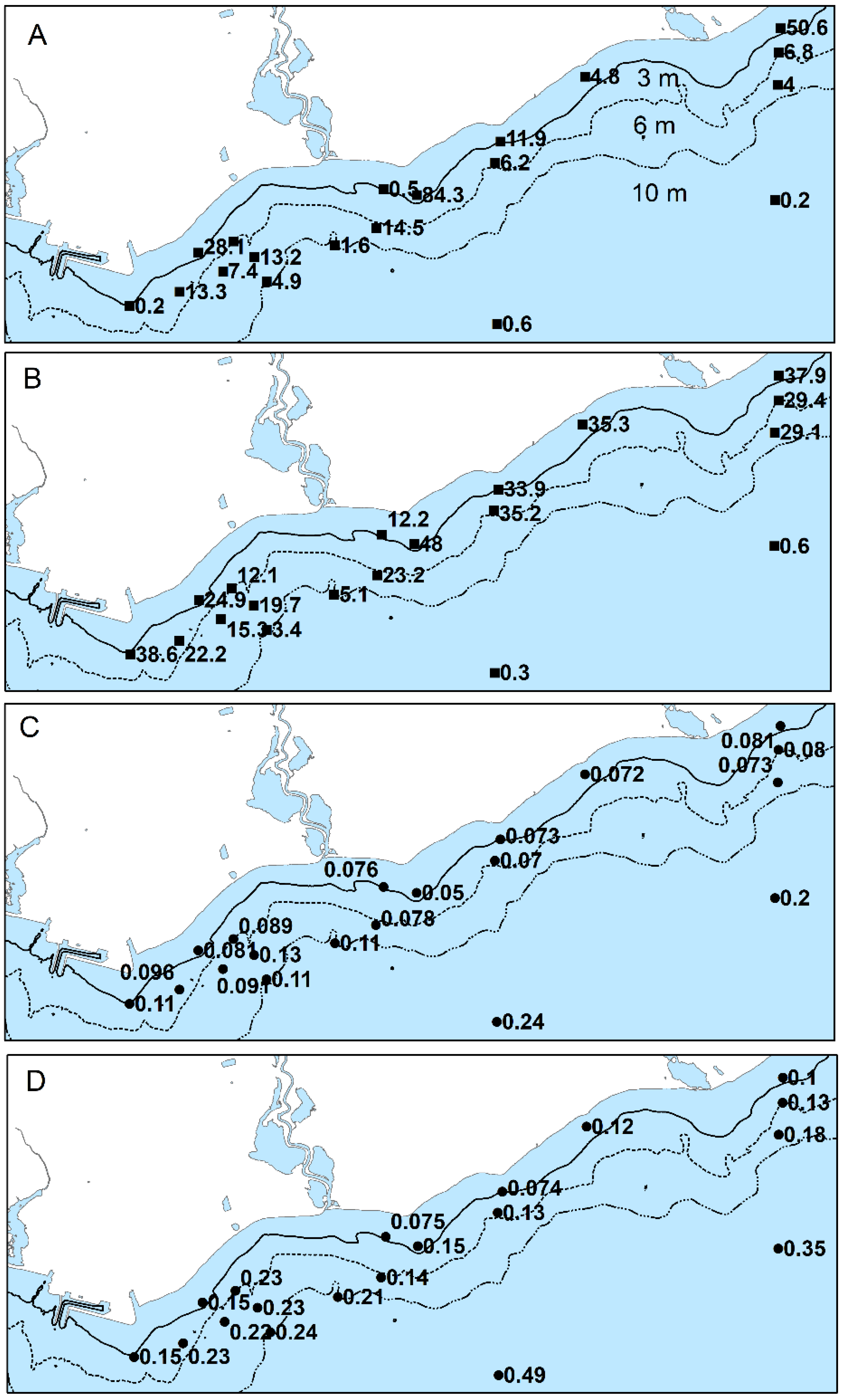
49]. In June 2012, Qp at sites east of the mouth of Duffins Creek were <0.082% compared with 0.081 to 0.13% at sites west of Duffins Creek and closer to the WPCP diffuser, with the exception of the 20 m sites (
Figure 9). In August, the levels were higher overall and again with the same east to west pattern. Notable are the higher Qp values (mostly >0.1%) oriented to the WSW away from the WPCP diffuser (
Figure 9).
The seeming contradiction between the distributions of
Cladophora biomass and Qp is potentially explained by the nuances of the ecology of
Cladophora that make the interpretation of field studies challenging. Unpredictable loss of attached biomass of
Cladophora due to physical disturbance by wave action and physiological deterioration of the alga over the growing season [
50] may confound inferences on overall biomass production among areas. The similar lake current regimes among sensors over the study area suggest that physical disturbance should be similar at sites of comparable depth. However, the effects of varying nutrient and temperature regimes on senescence of algae among locations cannot be discounted. The Pickering Nuclear Generating Plant discharges cooling water to the shoreline at the west end of the study area, affecting water temperature which may alter the timing of growth among locations.
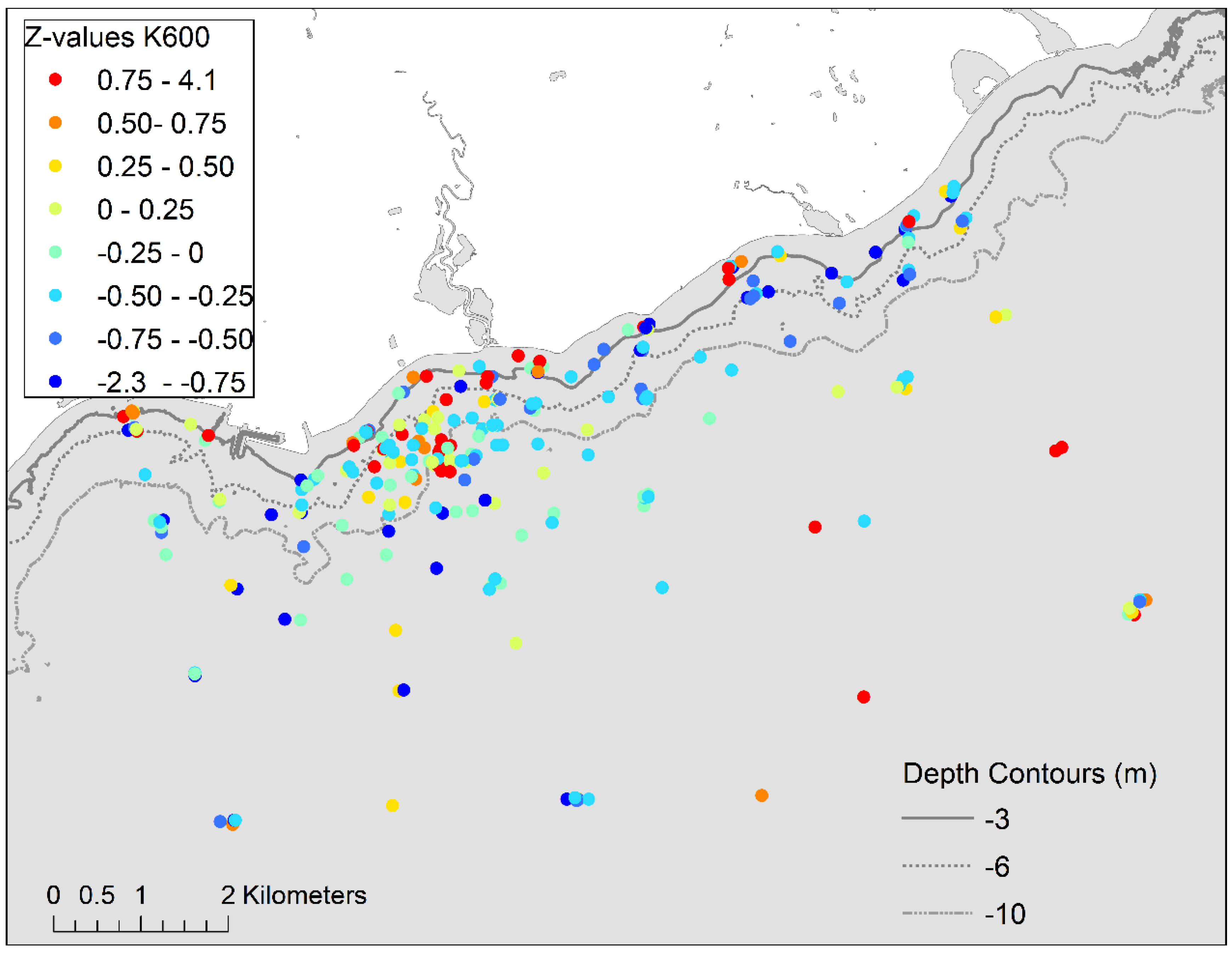
The interaction of phosphorus and light limitation on growth can also make the interpretation of Qp as an indicator of phosphorus enrichment challenging. At 20-m sites, Qp ranged from 0.20 to 0.24% and 0.35 to 0.49% in June and August, respectively, indicating phosphorus sufficiency due to light limitation of growth as evident from the low biomass. Discerning the effects of light limitation on Qp is more difficult when PAR flux is suboptimal for growth but not fully limiting [
51]. Light transmission at depths of
Cladophora growth was variable as inferred from beam attenuation (K
660) measurements in the upper water column which suggested that transmission is higher at times over the eastern section of the shoreline than the central-west section. The two largest discharges to the study area, the Duffins Creek WPCP diffuser and the mouth of Duffins Creek, are in the central-west section of the shoreline. Characterization of the PAR regime and
Cladophora growth modeling (e.g., [
48]) are required to distinguish light effects on Qp levels and would be useful in more fully resolving the reasons for the variability in Qp among areas.
The abundance of
Cladophora in 2012 appears lower than in 2008 [
9]. Average biomass at 3-m sites in the Ajax area in June and July of 2008 was 130 and 78 g/m
2, respectively, falling to 44- and 29- at 6-m sites. This compares with the highest average biomass of 84 and 34 g/m
2 at any 3- and 6-m site, respectively, in 2012 (
Figure 9). The growth of
Cladophora may vary from year to year because of differences in weather, lake conditions and possibly differences in nutrient loading. Biomass levels have varied widely where monitored annually [
12].
Earlier studies examining why
Cladophora is abundant in Lake Ontario [
9,
10,
11,
13,
15,
16,
18], an oligotrophic lake [
14], have made appreciable progress yet questions remain that arguably stand in the way of a strategy to abate overabundance of Cladophora. Ecological changes in the lake after the invasion of dreissenid mussels account for a general increase in
Cladophora. A key factor has been a lake-wide increase in water clarity, with more light reaching the lakebed, resulting in better growing conditions and more lakebed habitat for
Cladophora [
16,
18].
Less certain is the understanding of the pathways by which phosphorus loaded to the lake fuels the widespread abundance of
Cladophora in the lake. Nutrient loading in urban areas has been identified as a contributing factor to differences in
Cladophora abundance among areas in Lake Ontario [
9]. This reasonably suggests that the lake-wide regional supply of phosphorus when augmented by local-area sources results in the increased growth of
Cladophora with the possible exceptions of when either (i) the lake-wide supply exceeds the local-area supply, or, (ii) when physical conditions such as light also limit algae growth. More speculative is the hypothesized beneficial effects of increased nutrient focusing at the lakebed due to the heightened biological activity associated with a high concentration of mussel biomass. There is a growing case that a
Dreissena-colonized lakebed leaks dissolved phosphorus into the base of the water column as envisaged by Hecky et al. [
20] and observed by Ozersky et al. [
44] and Waples et al. [
25] using incubation chamber experiments.
4.4. Dreissenid Mussels as Modifiers of the Lakebed Environment
The abundance and wide distribution of
Dreissena shapes the physical and biological character of the lakebed of the study area and, Lake Ontario in general [
21,
22], aspects of which may affect
Cladophora. Mussel abundance was variable within sites and sub-areas suggesting that any associations between mussels, lakebed nutrients and
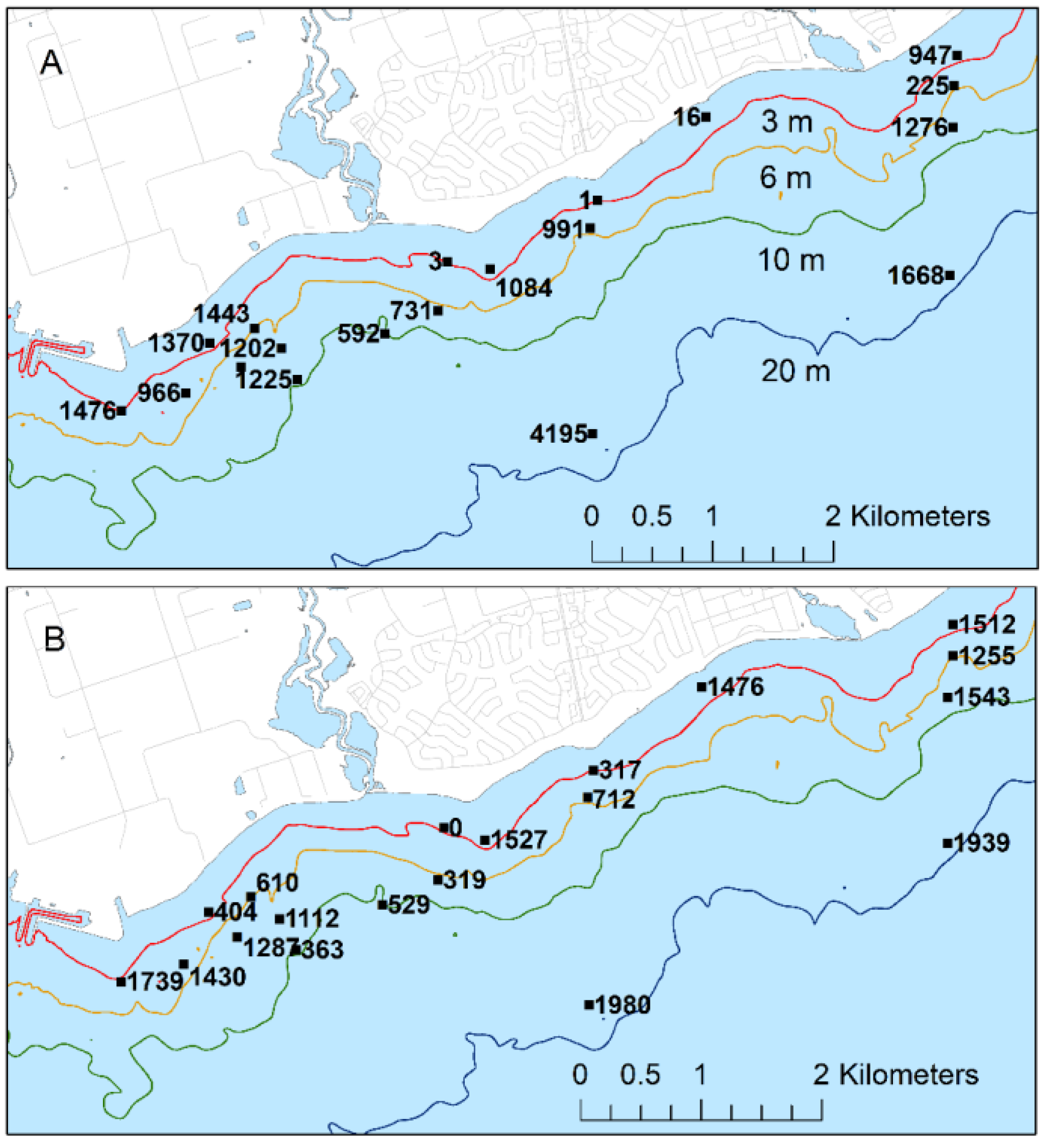
Cladophora would likely be difficult to detect and likely be most evident at the scale of patches (i.e., among sampling quadrats). Mussel biomass appeared higher west of the WPCP diffuser and on the eastern margins of the study area contrasting with lower abundance east of the WPCP diffuser and east of the mouth of Duffins Creek, an area more strongly influenced by sedimentation from Duffins Creek [
26]. Biomass was consistently high at sites WSW of the WPCP outfall, suggesting a positive effect of the proximity to the outfall. Abundance was unrelated to depth other than that it was consistently high at the two deepest sites, suggesting that physical disturbance from waves had a negative effect on abundance.
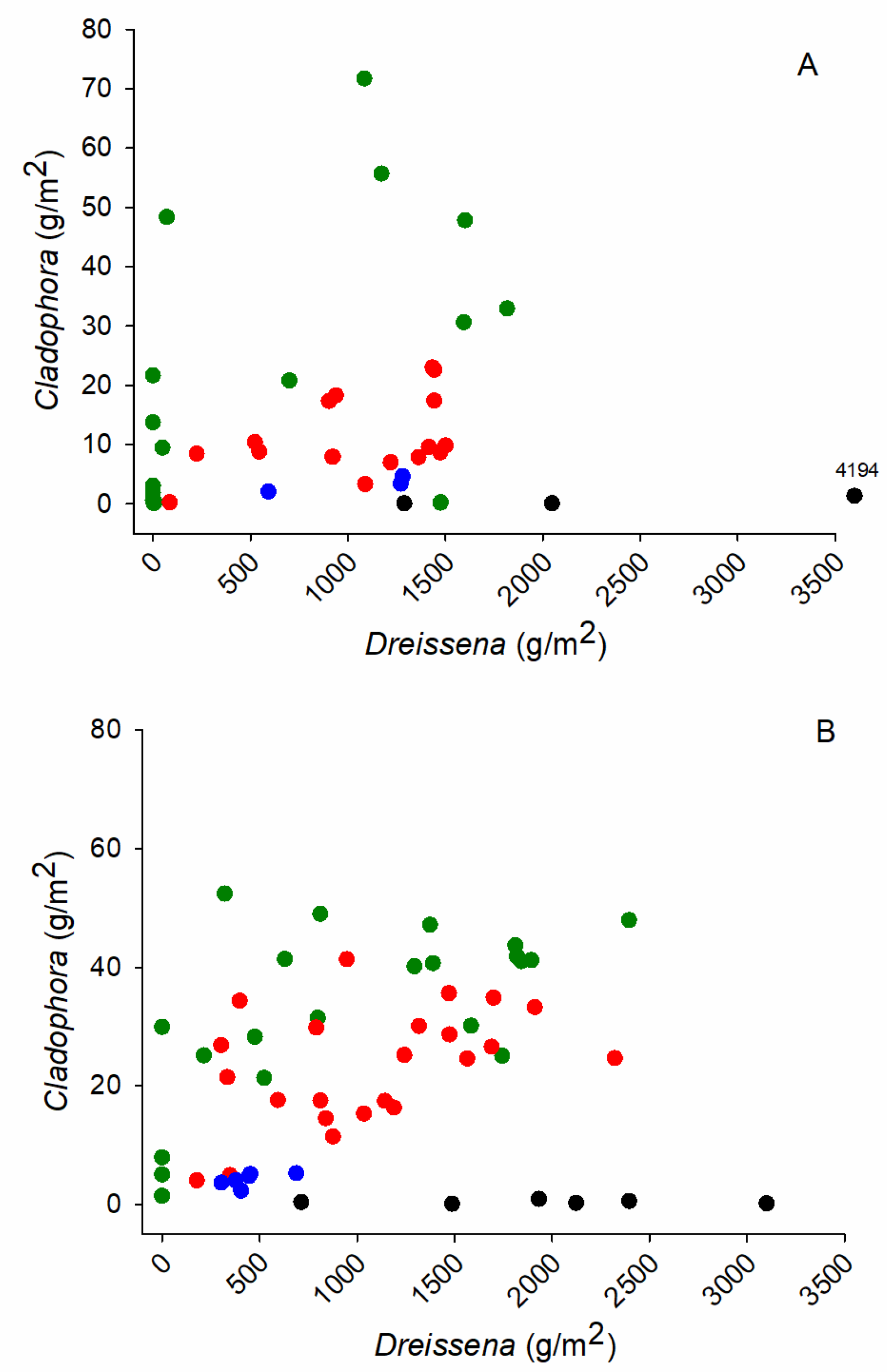
The higher biomass of
Cladophora seen in some quadrats with high mussel abundance is suggestive of a beneficial association, however, the correlation between
Cladophora and
Dreissena biomass at sites <8 m, the depth range of strong
Cladophora growth, was weak. A stronger positive association between
Cladophora and
Dreissena was detected in the regional study using a larger dataset, which also including the data here, over a broader geographic area [
10].
The possibility that internal phosphorus loading facilitated by Dreissena adds to, and obscures, local-area phosphorus loading, contributing to the growth of Cladophora, is neither supported nor well resolved with the data here. The weak association between Cladophora and Dreissena biomass, the absence of spatial association between Cladophora biomass and areas of nutrient enrichment, and in consideration of the sheer abundance of Dreissena and a growing literature that indicates that Dreissena affects lake nutrient pathways suggests that efforts to manage Cladophora in areas with nutrient pollution be mindful of possible interactions with dreissenid mussels.
4.5. Summary
The lands draining to the shores of the towns of Ajax and Pickering and, also beyond the drainage area, affect water quality in the adjacent coastline of Lake Ontario. Within the water column, the discharge of treated sewage stands out because of the concentration gradients of phosphorus and nitrogen that occur over the mixing area. Dilution rapidly reduced levels to those approaching nearshore background concentrations with subtle gradients extending away from the outfall diffuser that are detectable for some distance at times. The prevailing alongshore flow of the coastal zone frequently distributes affected water parallel to the shoreline. Runoff into the lake also added to the phosphorus enrichment in the area as indicated by the elevated DP and TP in shallow water at the shoreline. In general, water quality along the shoreline was more perturbed than further into the lake, however, it was difficult to distinguish the individual influences of Duffins Creek and other runoff points due to mixing along the shoreline.
The growth of phytoplankton was not a concern, as inferred from chlorophyll a concentrations. With the possible exception of the immediate mixing zone near the WPCP diffuser, oligotrophic conditions, as inferred from chlorophyll a and TP, prevailed over the study area. In stark contrast, the green algae Cladophora, forming lawns growing on the lakebed, was abundant at depths <10 m. Biomass exceeded the criteria indicating nuisance levels in some samples.
The distribution of Cladophora biomass was not related to the locations of nutrient loading, or the patterns of water quality measured in the water column. Measurements of dissolved nutrients at the lakebed were used to detect the influence of loading sources on nutrient levels in proximity to growing Cladophora. At times, there was little spatial pattern in levels of dissolved nutrients, and overall, evidence that levels were affected by biological uptake. However, spatial patterns in concentrations indicated intermittent, and spatially varying, enrichment with SRP, DP and TIN near the WPCP diffuser and with DP along the shoreline.
Unlike Cladophora biomass which was seemingly unrelated to either water column or nearbed nutrient levels, the phosphorus levels in Cladophora (Qp), an indicator of nutrient sufficiency, were higher in the area of the WPCP diffuser. Measurements of light transmission through the water column also suggest that light limitation, presumably affecting the growth of Cladophora, should be considered as a modifying factor. While the possible impact of light limitation merits further investigation, the balance of the Qp data provides evidence of phosphorus enrichment.
The lakebed was widely colonized with Dreissena, suggesting that filter-feeding by mussels might augment the phosphorus supply to the nearshore lakebed and Cladophora during the frequent onshore circulation of offshore water. There was no correspondence between Dreissena biomass and dissolved nutrient levels on the lakebed, however, biomass of Cladophora was weakly correlated with the biomass of Dreissena at shallow depths.
The findings here describing the covariation between
Cladophora and water quality over a local-area scale of variability have similarities to those reported over a regional scale. Like the regional scale analysis [
10], there was little correspondence between water column nutrient levels and the abundance of
Cladophora. Similar to the regional analysis, Qp levels were higher in areas where water quality suggested exposure of algae to higher phosphorus levels, and in both cases there was an element of uncertainty in the interpretation Qp due to possibly co-varying light regimes and phosphorus levels. The Qp data indicated that
Cladophora growth was likely phosphorus limited over the regions in the geographically broader study as it was over much of the Pickering-Ajax area. The biomass of
Cladophora was weakly correlated with the abundance of
Dreissena at both the local and regional scales, suggesting a positive effect of the mussels on algae growth but with the strength of the evidence leaving uncertainty.
The seeming lack of correspondence between the abundance of Cladophora and varying levels of water column phosphorus within the Pickering-Ajax study area is perplexing and points to the need to resolve whether loading of phosphorus from the offshore to the nearshore, dependent on lake-wide loading, plays a role in the wide-spread abundance of Cladophora in Lake Ontario. The attempt here to parse out the local-area effects on Cladophora abundance provides reasons to expect that phosphorus gradients in the study area will affect growth. Field studies of benthic algae are also challenging and fraught with an element of uncertainty due the typically high levels of variability on the lakebed, and the logistical difficulty of accurately sampling on the lakebed.
Phosphorus inputs to Lake Ontario may increase in the coming years due to anticipated continued urban growth [
52]. Widespread phosphorus limitation of growth is evident from this and the regional study with the implication that additional phosphorus supply will likely result in more algal growth. Attaining the phosphorus concentration and loading targets required to manage overabundance of
Cladophora as envisaged by the US-Canada Great Lakes Water Quality Agreement and the Province of Ontario Great Lakes Strategy will require an “ecosystem approach” in which investigations will need to advance understanding of the balances between (i) area and lake-wide nutrient supply, and, (ii) benthic and water column nutrient flux.














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}