Microbial Function and Hydrochemistry within a Stratified Anchialine Sinkhole: A Window into Coastal Aquifer Interactions


Abstract
:1. Introduction
2. Materials and Methods
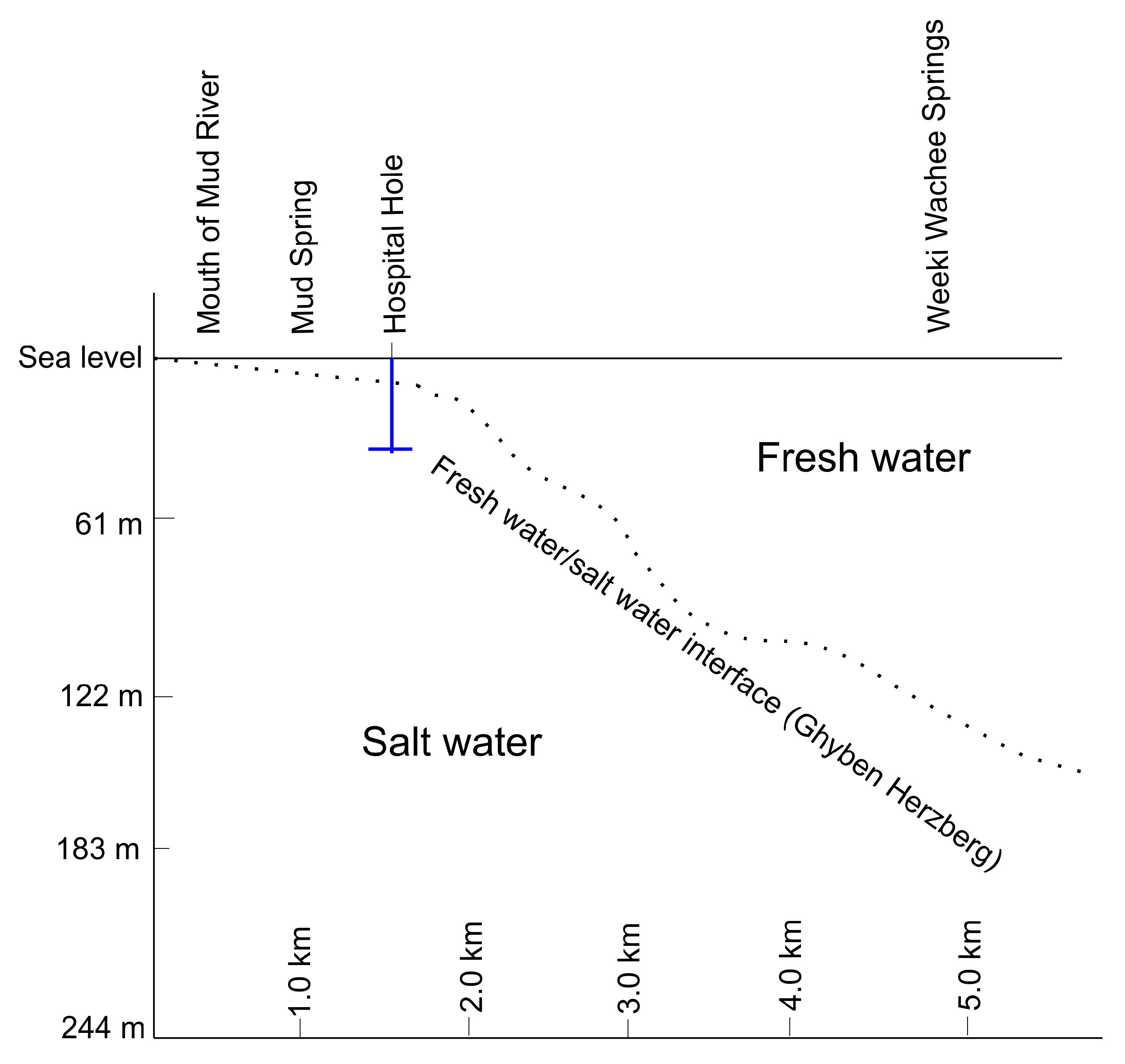
2.1. Site Description
2.2. Sampling Strategies
2.3. Hydrochemical Analysis
2.4. Biological Analysis
2.5. Statistical Analysis
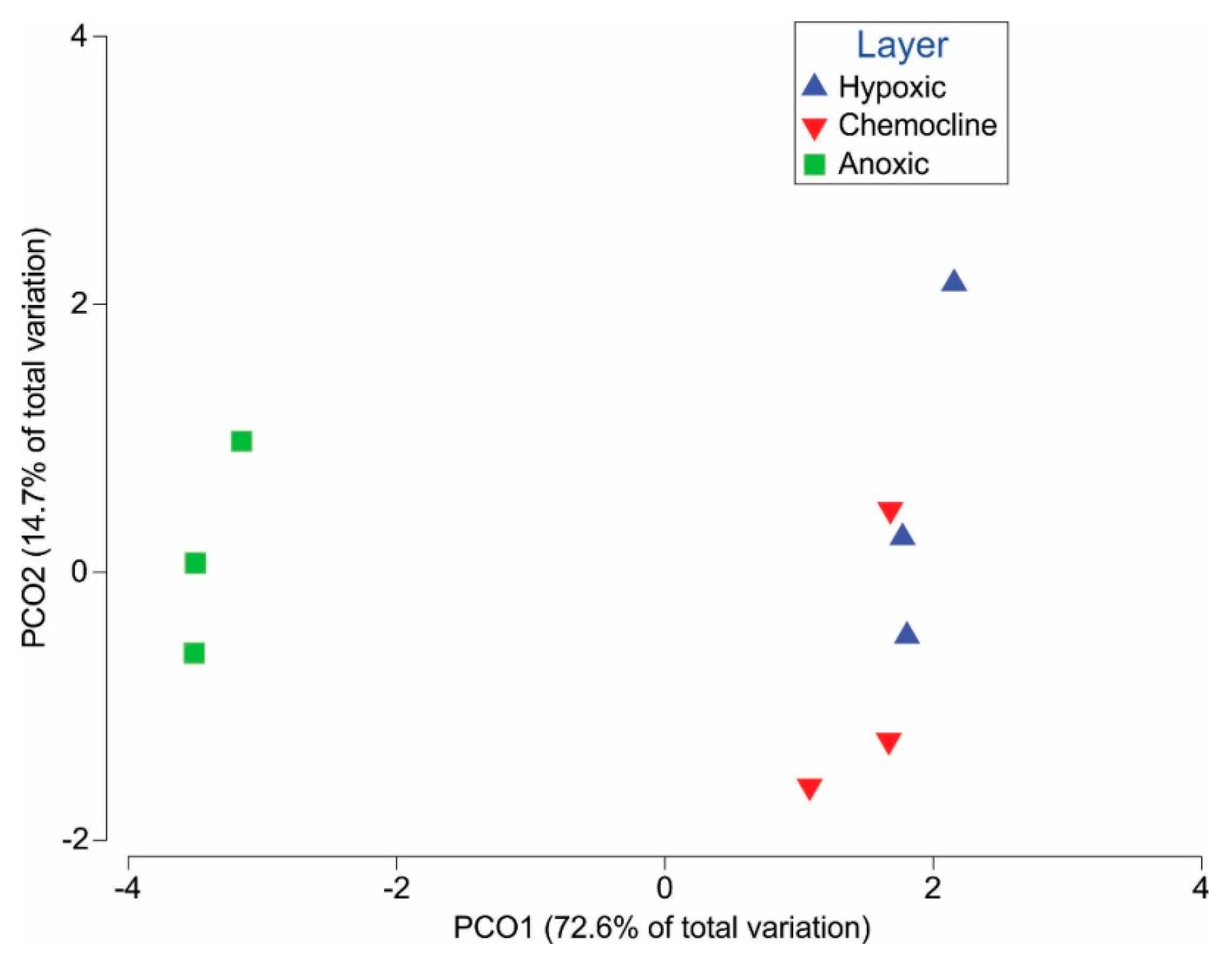
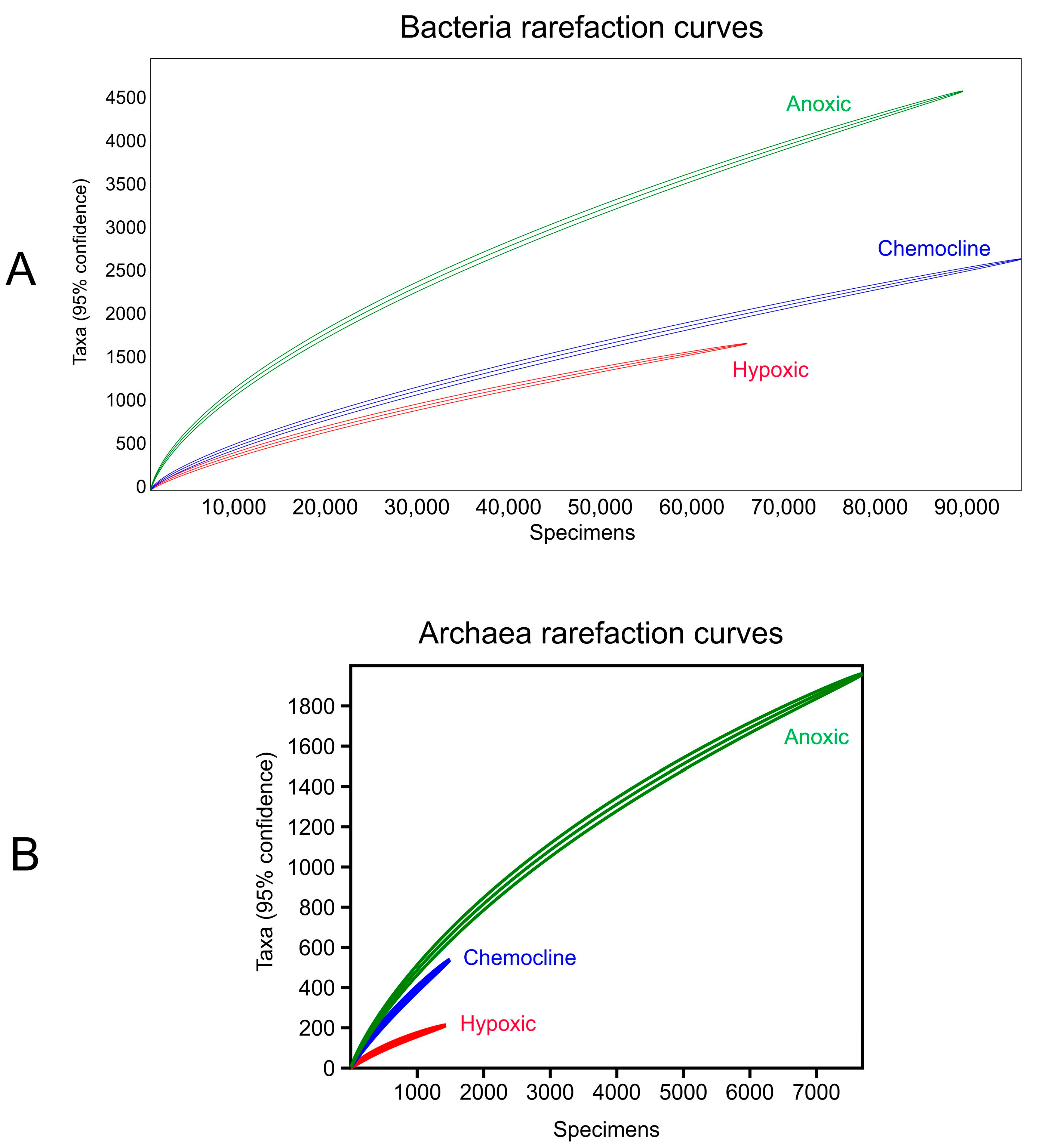
3. Results
4. Discussion
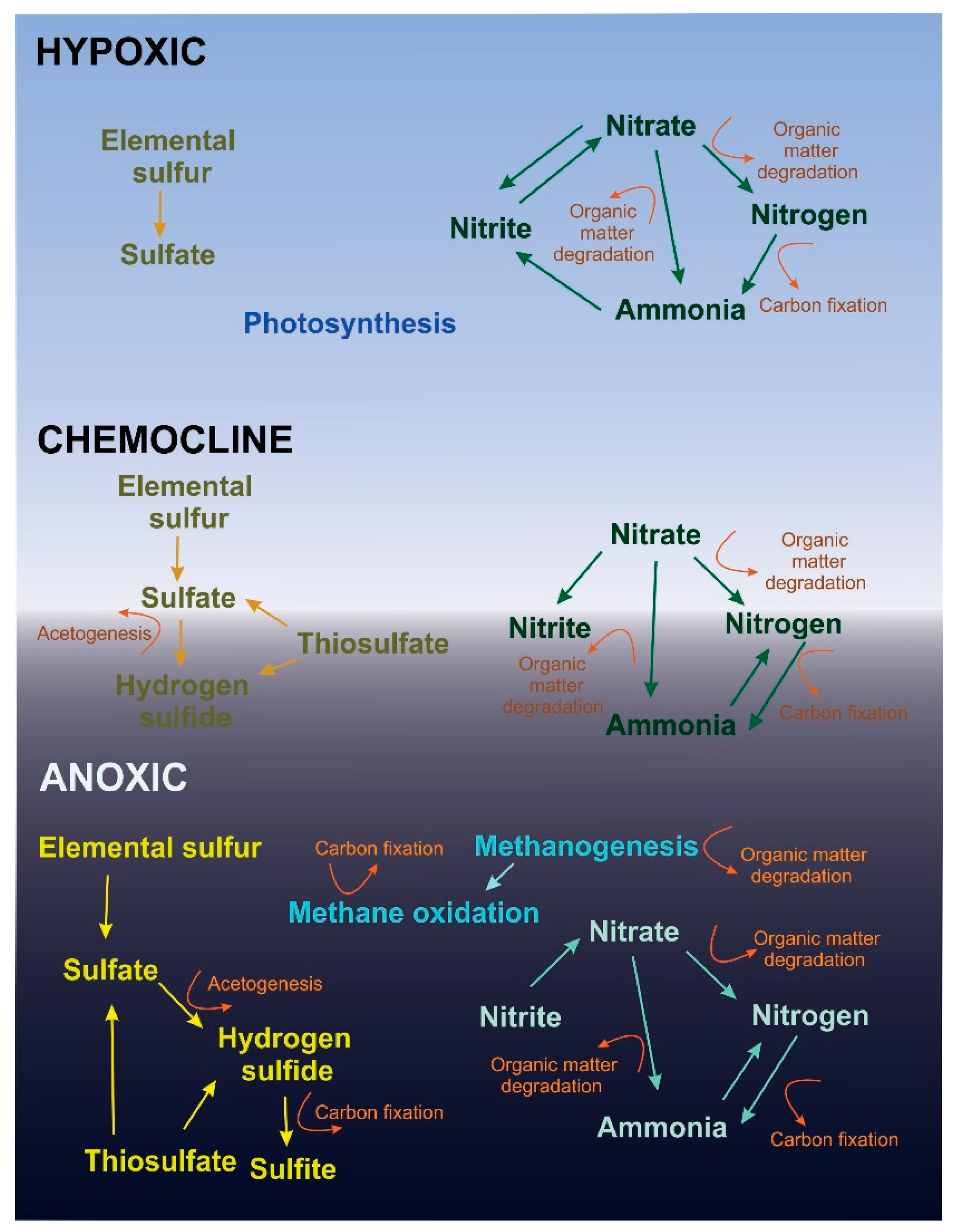
4.1. The Hypoxic Layer
4.2. The Anoxic Layer
4.3. The Chemocline
4.4. Hospital Hole: The Ecosystem
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ford, D.; Williams, P. Speleogenesis: The Development of Cave Systems. In Karst Hydrogeology and Geomorphology; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2007; pp. 209–270. ISBN 978-0-470-84996-5. [Google Scholar]
- Sprinkle, C.L. Hydrochemistry of the Floridan Aquifer System in Florida and in Parts of Georgia, South Carolina, and Alabama; Report No.: 1403I; United States Government Printing Office: Washington, DC, USA, 1989.
- Burgin, A.J.; Hamilton, S.K. Have we overemphasized the role of denitrification in aquatic ecosystems? A review of nitrate removal pathways. Front. Ecol. Environ. 2007, 5, 89–96. [Google Scholar] [CrossRef] [Green Version]
- Amon, R.M.; Benner, R. Bacterial utilization of different size classes of dissolved organic matter. Limnol. Oceanogr. 1996, 41, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Lovley, D.R.; Chapelle, F.H. Deep subsurface microbial processes. Rev. Geophys. 1995, 33, 365–381. [Google Scholar] [CrossRef]
- Sinclair, W.C. Preliminary Evaluation of the Water-Supply Potential of the Spring-River System in the Weeki Wachee Area and the Lower Withlacoochee River, West-Central Florida; U.S. Geological Survey, Water Resources Division: Reston, WV, USA, 1978.
- Gray, C.J.; Engel, A.S. Microbial diversity and impact on carbonate hydrochemistry across a changing hydrochemical gradient in a karst aquifer. ISME J. Multidiscip. J. Microb. Ecol. 2013, 7, 325–337. [Google Scholar]
- Moss, J.A.; Nocker, A.; Snyder, R.A. Microbial Characteristics of a Submerged Karst Cave System in Northern Florida. Geomicrobiol. J. 2011, 28, 719–731. [Google Scholar] [CrossRef]
- Coke, J.G.; Stoessell, R.K. An explanation for the lack of a dilute freshwater lens in unconfined tropical coastal aquifers: Yucatan example. Gulf Coast Assoc. Geol. Soc. Trans. 2006, 56, 785–792. [Google Scholar]
- Garman, M.K.; Rubelmann, H.; Karlen, D.; Wu, T.; Garey, J.R. Comparison of an inactive submarine spring with an active nearshore anchialine spring in Florida. Hydrobiologia 2011, 677, 65–87. [Google Scholar] [CrossRef]
- Takahashi, S.; Tomita, J.; Nishioka, K.; Hisada, T.; Nishijima, M. Development of a Prokaryotic Universal Primer for Simultaneous Analysis of Bacteria and Archaea Using Next-Generation Sequencing. PLoS ONE 2014, 9, e105592. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 7537–7541. [Google Scholar] [CrossRef] [PubMed]
- Menning, D.M.; Carraher-Stross, W.A.; Graham, E.D.; Thomas, D.N.; Phillips, A.R.; Scharping, R.J.; Garey, J.R. Aquifer Discharge Drives Microbial Community Change in Karst Estuaries. Estuaries Coasts 2017. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Cowell, B.C.; Botts, P.S. Factors influencing the distribution, abundance and growth of Lyngbya wollei in central Florida. Aquat. Bot. 1994, 49, 1–17. [Google Scholar] [CrossRef]
- Garman, M.K. The Biohydrochemistry of Submerged Coastal Karst Features in West Central Florida. Ph.D. Thesis, University of South Florida, Tampa, FL, USA, 2010. [Google Scholar]
- Bottrell, S.H.; Smart, P.L.; Whitaker, F.; Raiswell, R. Hydrochemistry and isotope systematics of sulphur in the mixing zone of Bahamian blue holes. Appl. Hydrochem. 1991, 6, 97–103. [Google Scholar]
- Gonzalez, B.; Iliffe, T.; Macalady, J.; Schaperdoth, I.; Kakuk, B. Microbial hotspots in anchialine blue holes: Initial discoveries from the Bahamas. Hydrobiologia 2011, 677, 149–156. [Google Scholar] [CrossRef]
- Garman, M.K.; Garey, J.R. The transition of a freshwater karst aquifer to an anoxic marine system. Estuaries 2005, 28, 686–693. [Google Scholar] [CrossRef]
- Kuenen, J.G. Anammox Bacteria: From discovery to application. Nat. Rev. Microbiol. 2007, 6, 320–321. [Google Scholar] [CrossRef] [PubMed]
- Oremland, R.S.; Polcin, S. Methanogenesis and Sulfate Reduction: Competitive and Noncompetitive Substrates in Estuarine Sediments. App. Environ. Microbiol. 1982, 44, 1270–1276. [Google Scholar]
- Fry, N.K.; Fredrickson, J.K.; Fishbain, S.; Wagner, M.; Stahl, D.A. Population structure of microbial communities associated with two deep, anaerobic, alkaline aquifers. Appl. Environ. Microbiol. 1997, 63, 1498–1504. [Google Scholar] [PubMed]
- Boetius, A.; Ravenschlag, K.; Schubert, C.J.; Rickert, D.; Widdel, F.; Gieseke, A.; Amann, A.; Jørgensen, B.B.; Witte, U.; Pfannkuche, O. A marine microbial consortium apparently mediating anaerobic oxidation of methane. Nature 2000, 407, 623–626. [Google Scholar] [CrossRef] [PubMed]
- Thamdrup, B.; Dalsgaard, T. Production of N2 through Anaerobic Ammonium Oxidation Coupled to Nitrate Reduction in Marine Sediments. Appl. Environ. Microbiol. 2002, 68, 1312–1318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kellermann, M.Y.; Wegener, G.; Elvert, M.; Yoshinaga, M.Y.; Lin, Y.S.; Holler, T.; Mollar, X.P.; Knittel, K.; Hinrichs, K.U. Autotrophy as a predominant mode of carbon fixation in anaerobic methane-oxidizing microbial communities. Proc. Natl. Acad. Sci. USA 2012, 109, 19321–19326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leschine, S.B. Cellulose degradation in anaerobic environments. Ann. Rev. Microbiol. 1995, 49, 399–426. [Google Scholar] [CrossRef] [PubMed]
- Harrington, D.; Maddox, G.; Hicks, R. Florida Springs Initiative Monitoring Network Report and Recognized Sources of Nitrate; Florida Department of Environmental Protection: Tallahassee, FL, USA, 2010.
- Boyer, D.G.; Pasquarell, G.C. Agricultural Land Use Impacts on Bacterial Water Quality in a Karst Groundwater Aqifer. JAWRA J. Am. Water Resour. Assoc. 1999, 35, 291–300. [Google Scholar] [CrossRef]
- Griebler, C.; Lueders, T. Microbial biodiversity in groundwater ecosystems. Freshw. Biol. 2009, 54, 649–677. [Google Scholar] [CrossRef] [Green Version]
- Blaschke, A.P.; Derx, J.; Zessner, M.; Kirnbauer, R.; Kavka, G.; Strelec, H.; Farnleitner, A.H.; Pang, L. Setback distances between small biological wastewater treatment systems and drinking water wells against virus contamination in alluvial aquifers. Sci. Total Environ. 2016, 573, 278–289. [Google Scholar] [CrossRef] [PubMed]
- Farnleitner, A.H.; Wilhartitz, I.; Ryzinska, G.; Kirschner, A.K.T.; Stadler, H.; Burtscher, M.M.; Hornek, R.; Szewzyk, U.; Herndl, G.; Mach, R.L. Bacterial dynamics in spring water of alpine karst aquifers indicates the presence of stable autochthonous microbial endokarst communities. Environ. Microbiol. 2005, 7, 1248–1259. [Google Scholar] [CrossRef] [PubMed]
- Macalady, J.L.; Lyon, E.H.; Koffman, B.; Albertson, L.K.; Meyer, K.; Galdenzi, S.; Mariani, S. Dominant Microbial Populations in Limestone-Corroding Stream Biofilms, Frasassi Cave System, Italy. Appl. Environ. Microbiol. 2006, 72, 5596–5609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pronk, M.; Goldscheider, N.; Zopfi, J. Microbial communities in karst groundwater and their potential use for biomonitoring. Hydrogeol. J. 2009, 17, 37–48. [Google Scholar] [CrossRef]
- Sacks, L.A.; Tihansky, A.B. Geochemical and Isotopic Composition of Ground Water with Emphasis on Sources of Sulfate in the Upper Floridan Aquifer and Intermediate Aquifer System in Southwest Florida; U.S. Geological Survey Water Resource Investigations Report 96-4146; US Department of the Interior: Washington, DC USA, 1996.







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Alkalinity (mg/L) | Ammonia (mg/L) | Nitrate (mg/L) | Nitrite (mg/L) | Sulfate (mg/L) | Sulfide (µg/L) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Surface | 145 | ±30 | 0.601 | ±0.008 | 0.440 | ±0.11 | 0.005 | ±0.001 | 0 | ±0 | 7 | ±1 |
| Hypoxic | 192 | ±38 | 0.434 | ±0.079 | 0.640 | ±0.15 | 0.000 | ±0.001 | 800 | ±158 | 1 | ±1 |
| Chemocline | 153 | ±28 | 0.356 | ±0.038 | 0.800 | ±0 | 0.197 | ±0.048 | 1000 | ±58 | 200 | ±6 |
| Anoxic | 206 | ±23 | 0.659 | ±032 | 5.080 | ±0.15 | 0.000 | ±0.001 | 1000 | ±55 | 30,000 | ±7972 |
| pH | Salinity (ppt) | Temperature (°C) | Dissolved oxygen (mg/L) | Phosphorus (mg/L) | ||||||||
| Surface | 7.91 | ±0.02 | 0.44 | ±0.05 | 23.93 | ±0.00 | 6.25 | ±0.02 | 0.50 | ±0.16 | ||
| Hypoxic | 7.47 | ±0.16 | 10.09 | ±3.89 | 24.15 | ±0.10 | 0.24 | ±0.53 | 0.16 | ±0.0 | ||
| Chemocline | 7.42 | *** | 11.75 | *** | 24.18 | *** | 0.01 | *** | 0.15 | ±0.20 | ||
| Anoxic | 7.27 | ±0.55 | 14.90 | ±0.7 | 20.5 | ±1.04 | 0 | ±0.00 | 0.28 | ±0.00 | ||
| Bacteria | |||||
| Name | Hypoxic | Name | Chemocline | Name | Anoxic |
| Halioglobus | 15,033 | Sulfurimonas | 10,604 | Sulfurimonas | 12,659 |
| Escherichia | 2994 | Halioglobus | 10,264 | Escherichia | 2433 |
| Arcobacter | 782 | Arcobacter | 5757 | Unknown | 1434 |
| Sulfurimonas | 703 | Alkalilimnicola | 1023 | Desulfocapsa | 1144 |
| Prochlorococcus | 182 | Emcibacter | 366 | Christensenella | 745 |
| Oceanicola | 148 | Unknown | 344 | Lentimicrobium | 624 |
| Emcibacter | 136 | Desulfocapsa | 275 | Arcobacter | 565 |
| Alkalilimnicola | 129 | Escherichia | 204 | Erysipelothrix | 469 |
| Rhodobacter | 109 | Lentimicrobium | 133 | Desulfosarcina | 354 |
| Citreicella | 87 | Sulfurovum | 129 | Mariniphaga | 304 |
| Thalassobius | 84 | Christensenella | 76 | Candidatus Aquiluna | 276 |
| Unknown | 56 | Desulfosarcina | 71 | Anammoximicrobium | 245 |
| Clostridium | 46 | Thalassobius | 65 | Ornatilinea | 168 |
| Citreimonas | 41 | Prochlorococcus | 51 | Claocimonetes | 158 |
| Methylotenera | 32 | Erysipelothrix | 48 | Sulfurovum | 156 |
| Desulfocapsa | 30 | Mariniphaga | 48 | Desulfotalea | 155 |
| Hypnocyclicus | 28 | Candidatus Aquiluna | 42 | Thermomarinilinea | 140 |
| Pelobacter | 26 | Dissulfuribacter | 40 | Sphingobacteriia | 139 |
| Lentimicrobium | 25 | Citreicella | 40 | Nibribacter | 129 |
| Streptococcus | 22 | Draconibacterium | 34 | Dissulfuribacter | 126 |
| Dechloromonas | 22 | Anammoximicrobium | 33 | Cryobacterium | 117 |
| Nitrospina | 18 | Cytophaga | 29 | Pelolinea | 112 |
| Psychrobacter | 17 | Lutibacter | 28 | Draconibacterium | 104 |
| Thiobacillus | 13 | Citreimonas | 28 | Cytophaga | 100 |
| TOTAL | 20,764 | TOTAL | 29,731 | TOTAL | 22,855 |
| Archaea | |||||
| Name | Hypoxic | Name | Chemocline | Name | Anoxic |
| Halarchaeum | 0 | Halarchaeum | 9 | Halarchaeum | 54 |
| Haloparvum | 2 | Haloparvum | 0 | Haloparvum | 0 |
| Halorubrum | 1 | Halorubrum | 3 | Halorubrum | 16 |
| Methanobacterium | 5 | Methanobacterium | 5 | Methanobacterium | 66 |
| Methanobrevibacter | 3 | Methanobrevibacter | 4 | Methanobrevibacter | 69 |
| Methanocalculus | 1 | Methanocalculus | 12 | Methanocalculus | 115 |
| Methanocaldococcus | 1 | Methanocaldococcus | 5 | Methanocaldococcus | 55 |
| Methanococcus | 6 | Methanococcus | 72 | Methanococcus | 599 |
| Methanoculleus | 0 | Methanoculleus | 4 | Methanoculleus | 20 |
| Methanomicrobium | 0 | Methanomicrobium | 2 | Methanomicrobium | 31 |
| Methanoplanus | 2 | Methanoplanus | 0 | Methanoplanus | 0 |
| Methanosaeta | 0 | Methanosaeta | 2 | Methanosaeta | 36 |
| Methanosaeta | 6 | Methanosaeta | 3 | Methanosaeta | 1 |
| Methanosphaera | 1 | Methanosphaera | 1 | Methanosphaera | 1 |
| Methanospirillum | 4 | Methanospirillum | 2 | Methanospirillum | 0 |
| Methanothermococcus | 3 | Methanothermococcus | 5 | Methanothermococcus | 13 |
| Nitrosopulmilus | 365 | Nitrosopulmilus | 164 | Nitrosopulmilus | 6 |
| Bacteria | Total per Layer | Fecal/ WTP/ pathogen | Anaerobes | Aerobes | Facultative Aerobe/ Anaerobe | Halotolerant | Photosynthetic | Methane Oxidizers | ||
| Hypoxic | 20,947 | Total | 3956 | 931 | 651 | 19,037 | 16,715 | 299 | 39 | |
| Percent | 18.9% | 4.4% | 3.1% | 90.9% | 79.8% | 1.4% | 0.2% | |||
| Chemocline | 30,301 | Total | 6667 | 6576 | 725 | 22,396 | 23,111 | 71 | 277 | |
| Percent | 22.0% | 21.7% | 2.4% | 73.9% | 76.3% | 0.2% | 0.9% | |||
| Anoxic | 24,167 | Total | 5633 | 4937 | 666 | 16,363 | 15,033 | 6 | 1143 | |
| Percent | 23.3% | 20.4% | 2.8% | 67.7% | 62.2% | 0.0% | 4.7% | |||
| Total per Layer | Sulfur Reducers | Sulfur Oxidizers | Nitrogen Reducers | Nitrogen Oxidizers | Iron Reducers | Manganese Oxidizers | Unknown Organism | |||
| Hypoxic | 20,947 | Total | 66 | 1847 | 17,104 | 7 | 8 | 0 | 59 | |
| Percent | 0.3% | 8.8% | 81.7% | 0.0% | 0.0% | 0.0% | 0.3% | |||
| Chemocline | 30,301 | Total | 502 | 17,616 | 28,137 | 35 | 42 | 0 | 407 | |
| Percent | 1.7% | 58.1% | 92.9% | 0.1% | 0.1% | 0.0% | 1.3% | |||
| Anoxic | 24,167 | Total | 2289 | 13,466 | 14,711 | 246 | 152 | 0 | 1882 | |
| Percent | 9.5% | 55.7% | 60.9% | 1.0% | 0.6% | 0.0% | 7.8% | |||
| Archaea | Total per Layer | Fecal/ WTP/ Pathogen | Halotolerant | Aerobe | Anaerobe | Facultative | Methanogen | Sulfate Reducers | ||
| Hypoxic | 402 | Total | 19 | 373 | 368 | 32 | 0 | 32 | 0 | |
| Percent | 4.7% | 92.8% | 91.5% | 8.0% | 0.0% | 8.0% | 0.0% | |||
| Chemocline | 295 | Total | 20 | 193 | 166 | 119 | 9 | 121 | 0 | |
| Percent | 6.8% | 65.4% | 56.3% | 40.3% | 3.1% | 41.0% | 0.0% | |||
| Anoxic | 1083 | Total | 204 | 204 | 22 | 1006 | 54 | 1006 | 0 | |
| Percent | 18.8% | 18.8% | 2.0% | 92.9% | 5.0% | 92.9% | 0.0% | |||
| Total per Layer | Nitrate Reduction | Nitrification | Hydrogen Sulfide Formation | Nitrogen Fixation | ||||||
| Hypoxic | 402 | Total | 3 | 365 | 4 | 10 | ||||
| Percent | 0.7% | 90.8% | 1.0% | 2.5% | ||||||
| Chemocline | 295 | Total | 12 | 163 | 17 | 77 | ||||
| Percent | 4.1% | 55.3% | 5.8% | 26.1% | ||||||
| Anoxic | 1083 | Total | 70 | 6 | 125 | 665 | ||||
| Percent | 6.5% | 0.6% | 11.5% | 61.4% |
| Well Identifier | Well Depth (m) | Casing Depth (m) | Distance to Coast (km) | Collection Date | Sulfate (mg/L) | Alkalinity (mg/L) | Specific Conductance (uS/cm) |
|---|---|---|---|---|---|---|---|
| 20891 | 29 | 13 | 1.2 | 11 February 2016 | 6.7 | 211.3 | 1999 |
| 20890 | 49 | 41 | 1.2 | 11 February 2016 | 26.6 | 300.1 | 2100 |
| 20690 | 92 | 84 | 2.4 | 24 March 2016 | 193 | 146.9 | 4643 |
| 20124 | 138 | 135 | 2.8 | 3 February 2016 | 618 | 140.8 | 358 |
| 20893 | 159 | 147 | 4.0 | 11 February 2016 | 1100 | 122 | 3851 |
| 20939 | 160 | 154 | 0.8 | 24 March 2016 | 2750 | 113 | 50,246 |
| 20735 | 171 | 46 | 8.0 | 31 January 2001 | 2569 | N/A | 12,600 |
| Hospital Hole hypoxic layer | 1.4 | 28 March 2016 | 800 | 145 | 20,000 | ||
| Hospital Hole anoxic layer | 1.4 | 28 March 2016 | 1000 | 200 | 25,000 | ||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Davis, M.C.; Garey, J.R. Microbial Function and Hydrochemistry within a Stratified Anchialine Sinkhole: A Window into Coastal Aquifer Interactions. Water 2018, 10, 972. https://doi.org/10.3390/w10080972
Davis MC, Garey JR. Microbial Function and Hydrochemistry within a Stratified Anchialine Sinkhole: A Window into Coastal Aquifer Interactions. Water. 2018; 10(8):972. https://doi.org/10.3390/w10080972
Chicago/Turabian StyleDavis, Madison C., and James R. Garey. 2018. "Microbial Function and Hydrochemistry within a Stratified Anchialine Sinkhole: A Window into Coastal Aquifer Interactions" Water 10, no. 8: 972. https://doi.org/10.3390/w10080972
APA StyleDavis, M. C., & Garey, J. R. (2018). Microbial Function and Hydrochemistry within a Stratified Anchialine Sinkhole: A Window into Coastal Aquifer Interactions. Water, 10(8), 972. https://doi.org/10.3390/w10080972