Effects of the Freezing–Thawing Cycle Mode on Alpine Vegetation in the Nagqu River Basin of the Qinghai–Tibet Plateau


Abstract
:1. Introduction
2. Materials and Methods
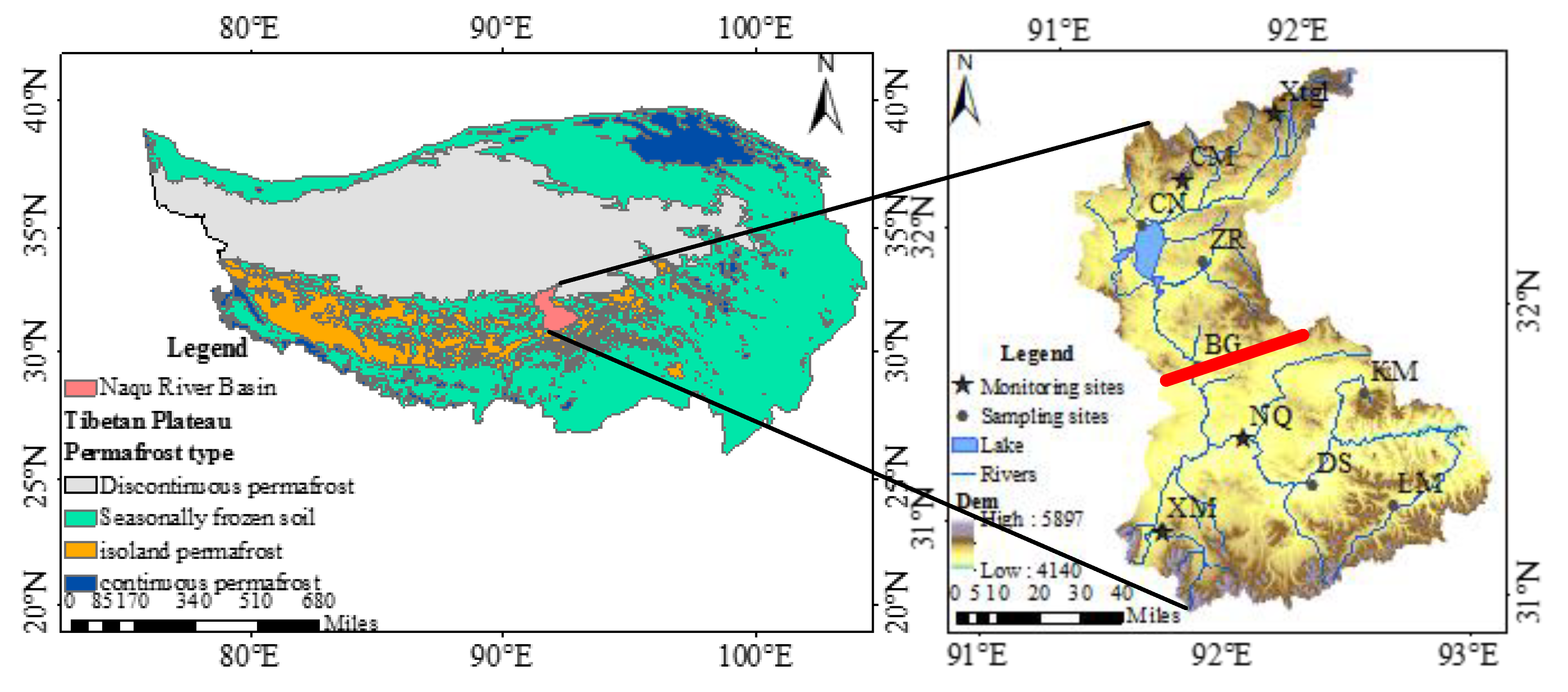
2.1. Study Area
2.2. Data Collecting and Processing
2.2.1. Soil Temperature and Volumetric Water Content Data
2.2.2. Vegetation Biomass Data
2.2.3. Soil Chemical Properties
2.2.4. Soil Microbial Data
2.3. Freezing–Thawing Cycle Index Calculation
3. Results
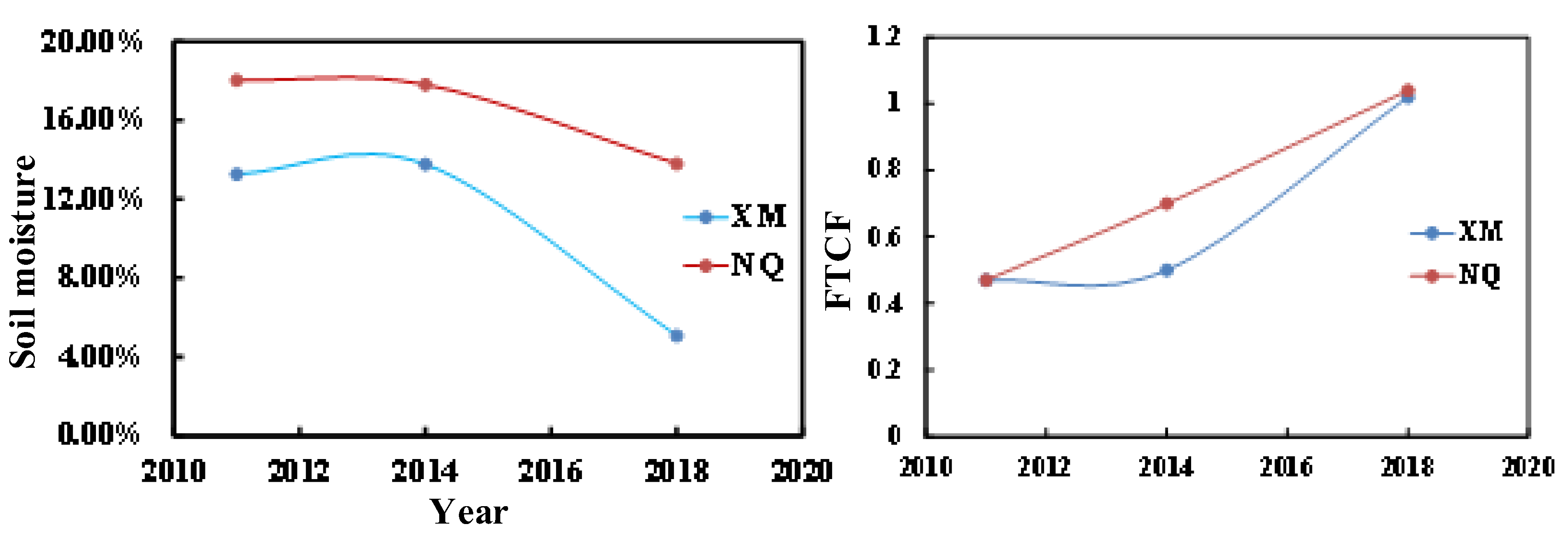
3.1. Freezing–Thawing Cycle Characteristics
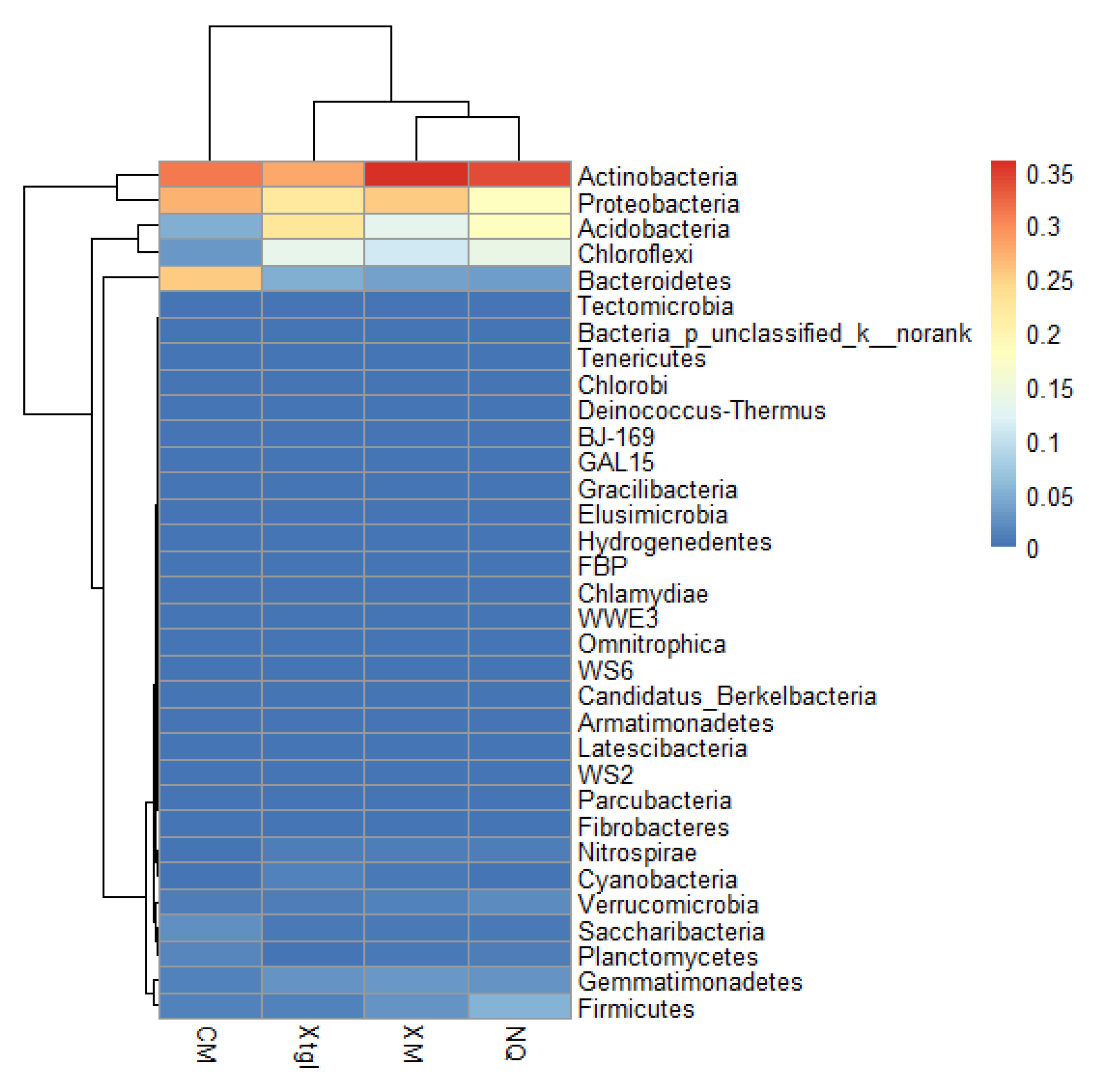
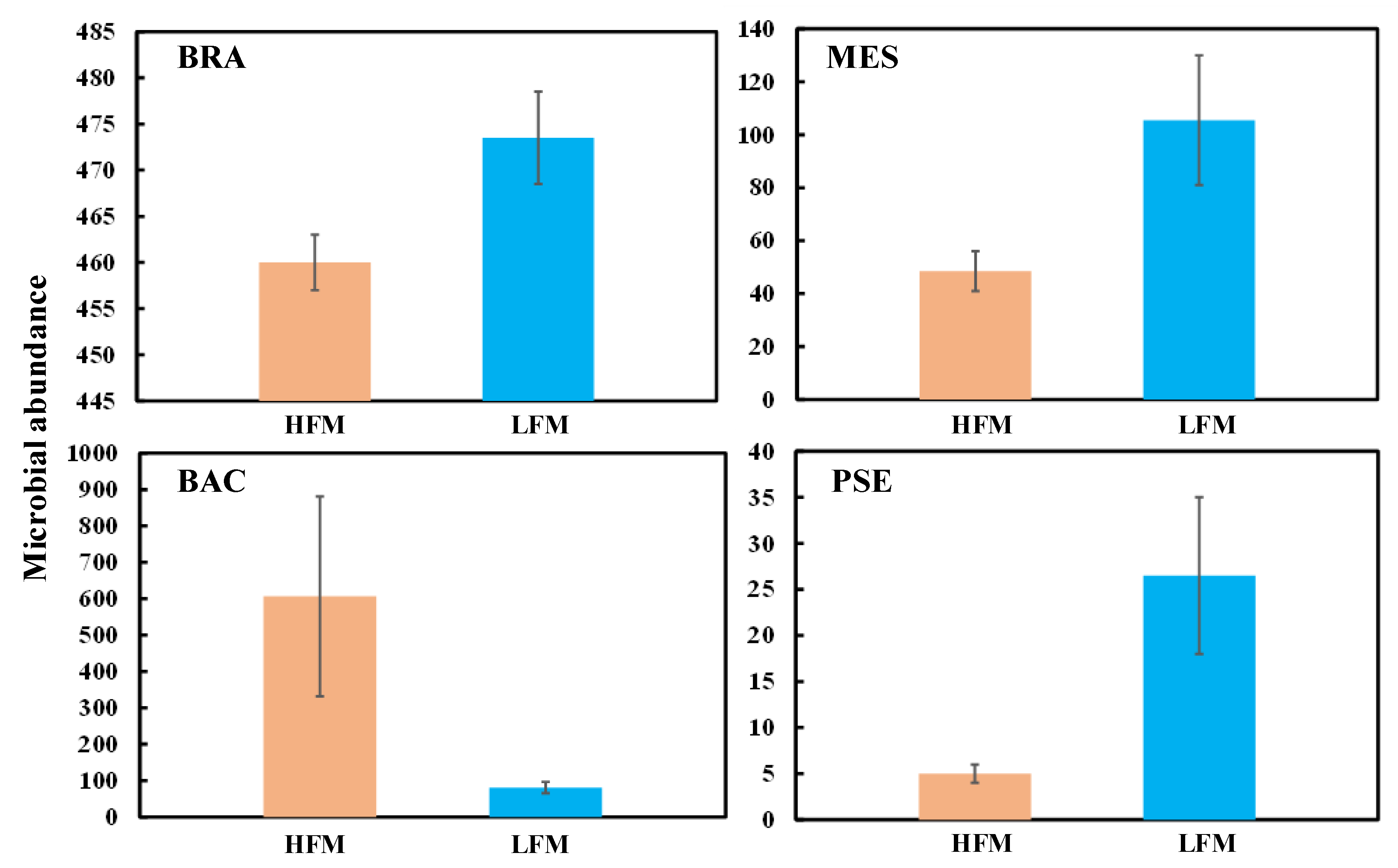
3.2. Microbial Characteristics
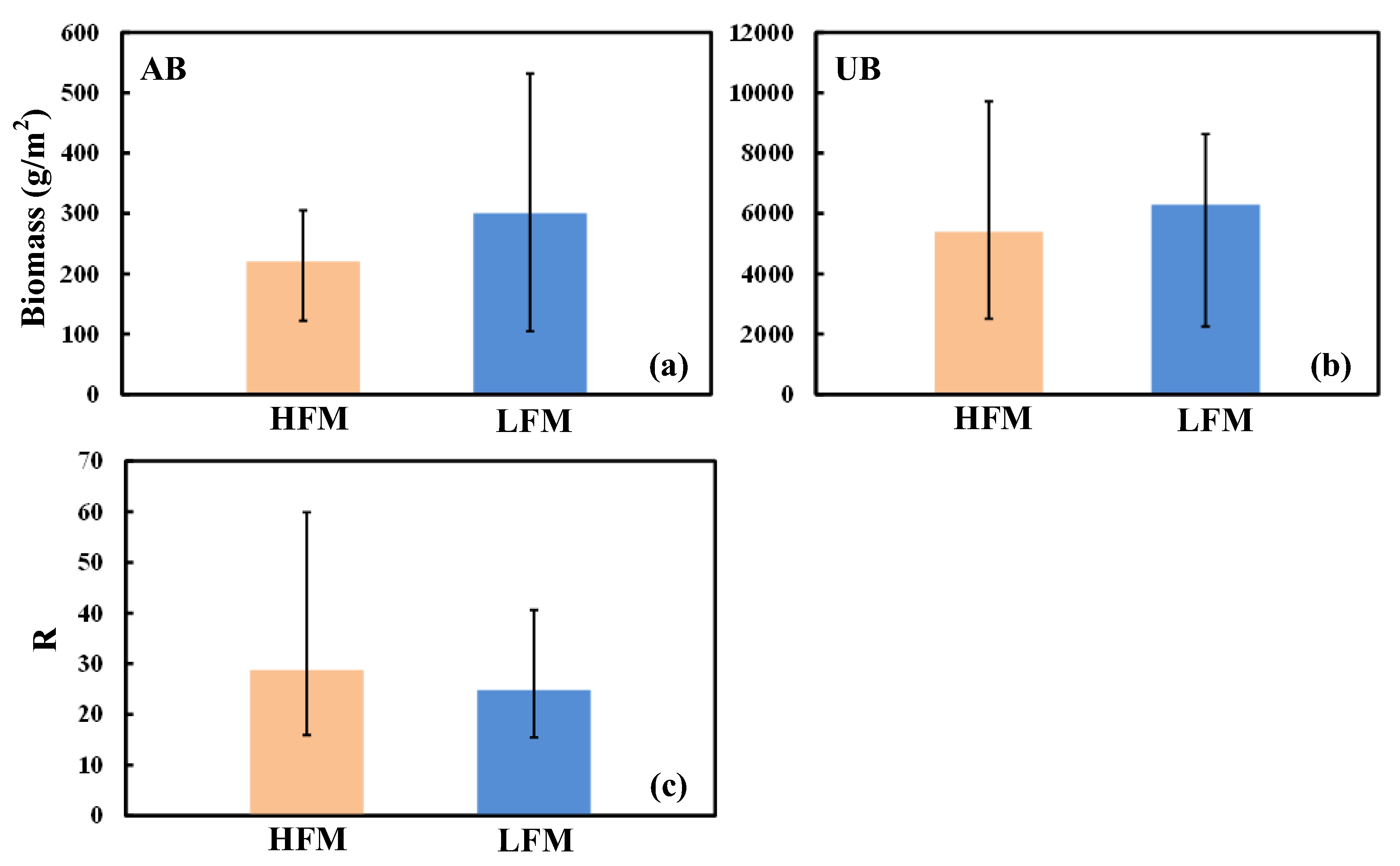
3.3. Aboveground and Underground Biomass
3.4. Correlation Analysis
3.4.1. Short-Term Fluctuation Analysis
3.4.2. Long-Term Fluctuation Analysis
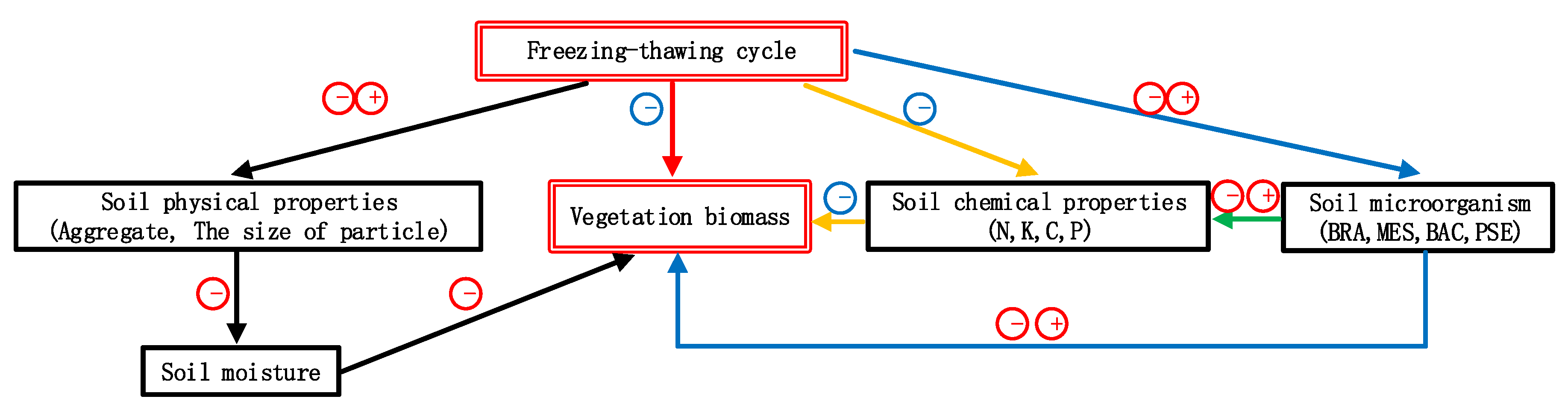
4. Discussion
5. Conclusions
- (1)
- The freezing–thawing cycle modes in the Nagqu River Basin are the HFM and the LFM. With the influence of climate change, the LFM is gradually shifting to the HFM.
- (2)
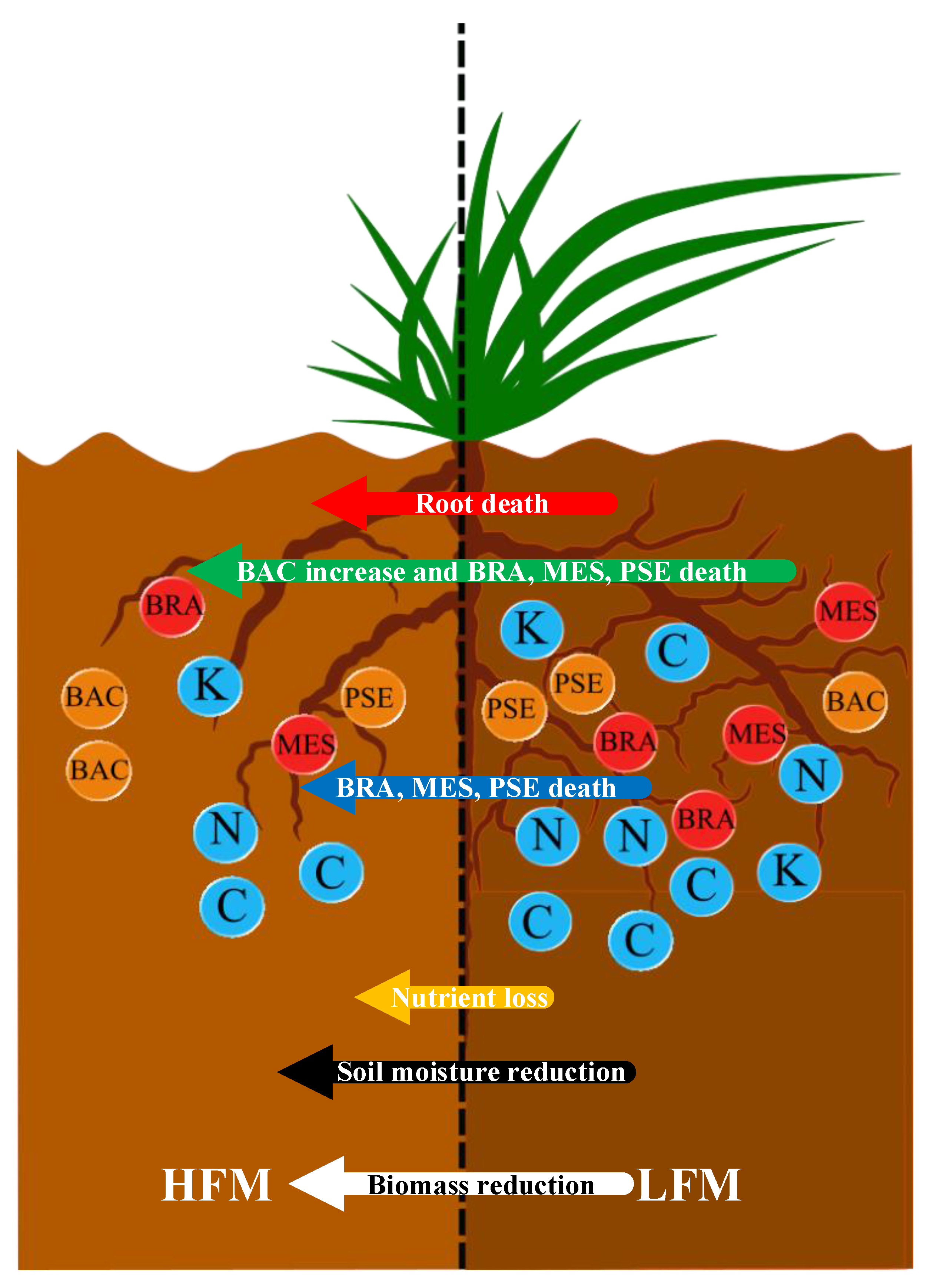
- The alpine vegetation biomass in the HFM is lower than that in the LFM. Frequent freezing–thawing cycles may lead to a reduction in root cell activity and even the death of root cells; on the other hand, high-frequency freezing–thawing cycles usually cause the death of certain microorganisms (BRA, MES, PSE) and weaken the roots’ nitrogen fixation ability and disease resistance. Moreover, the loss of soil nutrients is accelerated, the soil water holding capacity and soil moisture are reduced, and vegetation growth is inhibited.
Author Contributions
Funding
Conflicts of Interest
References
- Gill, R.A.; Jackson, R.B. Global patterns of root turnover for terrestrial ecosystems. New Phytol. 2000, 147, 13–31. [Google Scholar] [CrossRef]
- Morgan, R.P.C. Soil Erosion and Conservation; Blackwell Publishing: Oxford, UK, 2005. [Google Scholar]
- Luke McCormack, M.; Eissenstat, D.M.; Prasad, A.M.; Smithwick, E.A.H. Regional scale patterns of fine root lifespan and turnover under current and future climate. Glob. Chang. Biol. 2013, 19, 1697–1708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.; Tang, S.; Wang, C.; Zhou, P.; Tenuta, M.; Han, G.; Huang, D. Contribution of urine and dung patches from grazing sheep to methane and carbon dioxide fluxes in an Inner Mongolian desert grassland. Asian-Australas. J. Anim. Sci. 2012, 25, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Li, J.; Wei, Q.; Shan, Z.; Li, B.; Lang, D. Effects of vegetation construction on soil and water conservation in small watershed of purplish soil region, northern Sichuan. Trans. Chin. Soc. Agric. Eng. 2017, 33, 141–147. [Google Scholar]
- Wen, L.; Jinlan, W.; Xiaojiao, Z.; Shangli, S.; Wenxia, C. Effect of degradation and rebuilding of artificial grasslands on soil respiration and carbon and nitrogen pools on an alpine meadow of the Qinghai-Tibetan Plateau. Ecol. Eng. 2018, 111, 134–142. [Google Scholar] [CrossRef]
- Wu, Q.B.; Shi, B.; Liu, Y.Z. Study on the interaction between permafrost and highway along the Qinghai-Tibet Highway. Sci. Sin. D 2002, 32, 514–520. [Google Scholar]
- Walker, D.A.; Jia, G.J.; Epstein, H.E.; Raynolds, M.K.; Chapin, F.S., III; Copass, C.; Hinzman, L.D.; Knudson, J.A.; Maier, H.A.; Michaelson, G.J.; et al. Vegetation-soilthaw-depth relationships along a low-arctic bioclimate gradient, Alaska: Synthesis of information from the ATLAS studies. Permafr. Periglac. Process. 2003, 14, 103–123. [Google Scholar] [CrossRef]
- Shi, S.B.; Chen, G.C.; Yue, X.G.; Wang, X.Y.; Li, H.M.; Xu, W.H. Comparative studies of photosynthetic characteristics in typical alpine plants of the Qinghai-Tibet Plateau. J. Plant Ecol. 2006, 30, 40–46. [Google Scholar]
- Li, Y.N.; Zhao, L.; Zhao, Q.X.; Zhou, H.K. Effects of a 5-years mimic Temperature Increase to the structure and productivity of kobresia humilis meadow. Acta Agrestia Sin. 2004, 12, 236–239. [Google Scholar]
- Wang, Q.; Zhang, Q.P.; Zhou, W. Grassland coverage changes and analysis of the driving forces in Maqu County. Phys. Procedia 2012, 33, 1292–1297. [Google Scholar] [CrossRef]
- Brooker, R.W. Plant-plant interactions and environmental change. New Phytol. 2006, 171, 271–284. [Google Scholar] [CrossRef] [PubMed]
- Danby, R.K.; Hik, D.S. Responses of white spruce (Picea glauca) to experimental warming at a subarctic alpine treeline. Glob. Chang. Biol. 2007, 13, 437–451. [Google Scholar] [CrossRef]
- Bajguz, A.; Hayat, S. Effects of brassinosteroids on the plant responses to environmental stresses. Plant Physiol. Biochem. 2009, 47, 1–8. [Google Scholar] [CrossRef]
- Wu, X.; Shen, Z.Y. Effects of freezing-thawing cycle on greenhouse gases production and emission from soil. Chin. J. Ecol. 2010, 29, 1432–1439. [Google Scholar]
- Henry, H.A.L. Soil freeze-thaw cycle experiments: Trends, methodological weaknesses and suggested improvements. Soil Biol. Biochem. 2007, 39, 977–986. [Google Scholar] [CrossRef]
- Gong, J.D.; Qi, X.S.; Xie, Z.K.; Wang, Y.J. Effect of seasonal freezing on soil moisture and its significance for agriculture. J. Glaciol. Geocryol. 1997, 19, 328–333. [Google Scholar]
- Kreyling, J.; Peršoh, D.; Werner, S.; Benzenberg, M.; Wöllecke, J. Short-term impacts of soil freeze-thaw cycles on roots and root-associated fungi of Holcus lanatus and Calluna vulgaris. Plant Soil 2011, 353, 19–31. [Google Scholar] [CrossRef]
- Fitzhugh, R.D.; Driscoll, C.T.; Groffman, P.M.; Tierney, G.L.; Hardy, F.J.P. Effects of soil freezing disturbance on soil solution nitrogen, phosphorus, and carbon chemistry in a northern hardwood ecosystem. Biogeochemistry 2001, 56, 215–238. [Google Scholar] [CrossRef]
- Jarvis, S.C.; Stockdale, E.A.; Shepherd, M.A.; Powlson, D.S. Nitrogen mineralization in temperate agricultural soils: Processes and measurement. Adv. Agron. 1996, 57, 187–235. [Google Scholar]
- Christopher, S.F.; Shibata, H.; Ozawa, M.; Nakagawa, Y.; Mitchell, M.J. The effect of soil freezing on N cycling: Comparison of two headwater subcatchments with different vegetation and snowpack conditions in the northern Hokkaido island of Japan. Biogeochemistry 2008, 88, 15–30. [Google Scholar] [CrossRef]
- Zhang, H.O.; Xie, J.C.; Nan, H.P.; Han, Q.C.; Wang, N.; Zhang, Y. The interaction of freezing-thawing on soil aggregates and organic matter of pisha sandstone and sand compound soil. J. Soil Water Conserv. 2016, 30, 273–278. [Google Scholar]
- Starkloff, T.; Ritsema, C.; Stolte, J.; Larsbo, M.; Hessel, R. Quantifying the impact of a succession of freezing-thawing cycles on the pore network of a silty clay loam and a loamy sand topsoil using X-ray tomography. Catena 2017, 156, 365–374. [Google Scholar] [CrossRef]
- Six, J.; Bossuyt, H.; Degryze, S.; Denef, K. A history of research on the link between (micro) aggregates, soil biota, and soil organic matter dynamics. Soil Tillage Res. 2004, 79, 7–31. [Google Scholar] [CrossRef]
- Sahin, U.; Angin, I.K.; Iziloglu, F.M. Effect of freezing and thawing processes on some physical properties of saline–sodic soils mixed with sewage sludge or fly ash. Soil Tillage Res. 2008, 99, 254–260. [Google Scholar] [CrossRef]
- Han, L.; Wan, Z.M.; Sun, H.Y. Research Progress on the Effects of Freezing and Thawing on Soil Physical, Chemical and Biological Properties. Chin. J. Soil Sci. 2018, 49, 736–742. [Google Scholar]
- Finegold, L. Molecular and biophysical aspects of adaptation of life to temperatures below the freezing point. Adv. Space Res. 1996, 18, 87–95. [Google Scholar] [CrossRef]
- Liebner, S.; Rublack, K.; Stuehrmann, T.; Wagner, D. Diversity of aerobic methanotrophic bacteria in a permafrost active layer soil of the lena delta, siberia. Microb. Ecol. 2009, 57, 25–35. [Google Scholar] [CrossRef]
- Deng, X.M.; Wang, J.; Zhu, W.S.; Chen, R.S.; Liu, L.P. Effects of frost action on soil physical properties of plough pan. Chin. Sci. Bull. 1999, 43, 2583–2587. [Google Scholar] [CrossRef]
- Yang, S.Z.; Jin, H.J. Physiological and ecological effects of freezing and thawing processes on microorganisms in season-ally-froze ground and in permafrost. Acta Ecol. Sin. 2008, 28, 5056–5074. [Google Scholar]
- Anisimov, O.A. Potential feedback of thawing permafrost to the global climate system through methane emission. Environ. Res. Lett. 2017, 2, 045016. [Google Scholar] [CrossRef]
- Goldberg, S.D.; Borken, W.; Gebauer, G. N2O emission in a Norway spruce forest due to soil frost: Concentration and isotope profiles shed a new light on an old story. Biogeochemistry 2010, 97, 21–30. [Google Scholar] [CrossRef]
- Koven, C.D.; Riley, W.J.; Stern, A. Analysis of permafrost thermal dynamics and response to climate change in the CMIP5 Earth System Models. J. Clim. 2013, 26, 1877–1900. [Google Scholar] [CrossRef]
- Luo, D.; Wu, Q.; Jin, H. Recent changes in the active layer hickness across the northern hemisphere. Environ. Earth Sci. 2016, 75, 1–15. [Google Scholar] [CrossRef]
- Shiklomanov, N.I.; Streletskiy, D.A.; Nelson, F.E. Northern hemisphere component of the global circumpolar active layer monitoring (CALM) program. In Proceedings of the 10th International Conferences on Permafrost, Salekhard, Russia, 25–29 June 2012; pp. 377–382. [Google Scholar]
- Romanovsky, V.E.; Drozdov, D.S.; Oberman, N.G.; Malkova, G.V.; Kholodov, A.L.; Marchenko, S.S. Thermal state of permafrost in russia. Permafr. Periglac. Process. 2010, 21, 136–155. [Google Scholar] [CrossRef]
- Luo, B.; Zhi, H.; Duo, D. Analysis of Freezing and Thawing Processes on Typical Underlying Surface in Permafrost Area of Tibet Plateau. Plateau Mt. Meteorol. Res. 2018, 38, 13–18. [Google Scholar]
- McGuire, A.D.; Wirth, C.; Apps, M.; Beringer, J.; Clein, J.; Epstein, H.; Kicklighter, D.W.; Bhatti, J.; Chapin, F.S., III; de Groot, B.; et al. Environmental variation, vegetation distribution, carbon dynamics and water/energy exchange at high latitudes. J. Veg. Sci. 2002, 13, 301–314. [Google Scholar] [CrossRef]
- Christensen, T.R. Thawing sub-arctic permafrost: Effects on vegetation and methane emissions. Geophys. Res. Lett. 2004, 31, L04501. [Google Scholar] [CrossRef]
- Schuur, E.A.G.; Abbott, B. Climate change: High risk of permafrost thaw. Nature 2011, 480, 32–33. [Google Scholar] [CrossRef]
- Vapaavuori, E.M.; Rikala, R.; Ryyppo, A. Effects of root temperature on growth and photosynthesis in conifer seedlings during shoot elongation. Tree Physiol. 1992, 10, 217–230. [Google Scholar] [CrossRef]
- Levitt, J.; Siminovittch, D. The relation between frost resistance and the physical state of protoplasm. Can. J. Res. 1940, 18, 550–561. [Google Scholar] [CrossRef]
- Kennedy, A.D. Photosynthetic response of the Antarctic moss Polytrichum alpestre Hoppe to low temperatures and freeze-thaw stress. Polar Biol. 1993, 13, 271–279. [Google Scholar] [CrossRef]
- Gilichinsky, D. Microbial life in permafrost: A historical review. Permafr. Periglac. Process. 1995, 6, 243–250. [Google Scholar] [CrossRef]
- Zeng, Y.X.; Yu, Y.; Cai, M.H.; He, J.F.; Chen, B. A survey on cold-adapted microorganisms and their enzymes. J. Microbiol. 2004, 24, 83–88. [Google Scholar]
- Wang, J.Y.; Song, C.C.; Wang, X.W.; Wang, L.L. Progress in the study of effect of freeze-thaw processes on the organic carbon pool and microorganisms in soils. J. Glaciol. Geocryol. 2011, 33, 442–452. [Google Scholar]
- Jarvis, B.D.W.; Van Berkum, P.; Chen, W.X.; Nour, S.M.; Fernandez, M.P.; Cleyetmarel, J.C. Transfer of Rhizobium leti, Rhizobium huzkuii, Rhizobium ciceri, Rhizobium mediterraneum, and Rhizobium tianshanense to Mesorhizobium gene. nov. Int. J. Syst. Evol. Microbiol. 1997, 47, 895–898. [Google Scholar]
- Shi, X.X.; Shi, S.L.; Yang, J.; Wang, Z.F. Research advancement in taxonomy of Rhizobium leguminosarum. Grassl. Turf 2006, 1, 12–17. [Google Scholar] [CrossRef]
- China General Microbiological Culture Collection Center. Chinese Strain Catalogue; China Light Industry Press: Beijing, China, 1983; p. 57. [Google Scholar]
- Coy, R.M.; Held, D.W.; Kloepper, J.W. Rhizobacterial colonization of bermudagrass by Bacillus spp. in a Marvyn loamy sand soil. Appl. Soil Ecol. 2019, 141, 10–17. [Google Scholar] [CrossRef]
- Zhang, H.Y.; Li, Z.G. Sustainable use of Bacillus licheniformis and its resources. Soil 2001, 33, 92–97. [Google Scholar] [CrossRef]
- O’Sullivan, D.J.; O’Gara, F. Traits of fluorescent Pseudomonas spp. involved in suppression of plant root pathogens. Microbiol. Rev. 1992, 56, 662–676. [Google Scholar]
- Liu, J.; Fan, H.M.; Zhou, L.L.; Wu, M.; Chai, Y.; Liu, Y.H. Study on effects of freeze-thaw cycle on bulk density and porosity of black soil. J. Soil Water Conserv. 2009, 23, 186–189. [Google Scholar]
- Du, Z.Y.; Cai, Y.J.; Wang, X.D.; Yan, Y.; Lu, X.Y.; Liu, S.Z. Research progress on the effects of soil freeze-thaw on plant physiology and ecology. Chin. J. Eco-Agric. 2014, 22, 1–9. [Google Scholar] [CrossRef]
- Lu, Y.J. Interaction and Joint Regulation between Water and Soil Resources in the Alpine Region: A Case Study in the Naqu River Basin of the Tibetan Plateau; China Institute of Water Resources and Hydropower Research: Beijing, China, 2017. [Google Scholar]
- Yang, K.; Qin, J.; Zhao, L.; Chen, Y.Y.; Tang, W.J.; Han, M.L.; Chen, Z.Q.; Lv, N.; Ding, B.H.; Wu, H.; et al. A Multi-Scale Soil Moisture and Freeze-Thaw Monitoring Network on the Third Pole. Bull. Am. Meteorol. Soc. 2013, 94, 1907–1916. [Google Scholar] [CrossRef]
- Zhao, L.; Yang, K.; Qin, J.; Chen, Y.Y.; Tang, W.J.; Montzka, C.; Wu, H.; Lin, C.G.; Han, M.L.; Vereecken, H. Spatiotemporal analysis of soil moisture observations within a Tibetan mesoscale area and its implication to regional soil moisture measurements. J. Hydrol. 2013, 482, 92–104. [Google Scholar] [CrossRef]
- Qin, J.; Yang, K.; Lu, N.; Chen, Y.Y.; Zhao, L.; Han, M.L. Spatial upscaling of in-situ soil moisture measurements based on MODIS-derived apparent thermal inertia. Remote Sens. Environ. 2013, 138, 1–9. [Google Scholar] [CrossRef]
- Chen, Y.; Yang, K.; Qin, J.; Zhao, L.; Tang, W.; Han, M. Evaluation of AMSR-E retrievals and GLDAS simulations against observations of a soil moisture network on the central Tibetan plateau. J. Geophys. Res. Atmos. 2013, 118, 4466–4475. [Google Scholar] [CrossRef]
- China Soil Data. Available online: http://vdb3.soil.csdb.cn/ (accessed on 23 May 2019).
- Yan, Y.; Zhu, J.J.; Zhang, B.; Zhang, Y.J.; Lu, S.B.; Pan, Q.M. A review of belowground biomass allocation and its response to global climatic change in grassland ecosystems. Chin. J. Plant Ecol. 2017, 41, 585–596. [Google Scholar] [Green Version]
- Herrmann, A.; Witter, E. Sources of C and N contributing to the flush in mineralization upon freeze-thaw cycles in soils. Soil Biol. Biochem. 2002, 34, 1495–1505. [Google Scholar] [CrossRef]
- Fan, Z.P.; Li, S.G.; Li, F.Y.; Gao, H.C.; Yan, J.L. Effect of freezing-thawing on soil dissolved inorganic nitrogen and soil microbial biomass nitrogen in riparian zone. J. Meteorol. Environ. 2013, 29, 106–111. [Google Scholar]
- Wang, D.Y.; Ma, W.; Chang, X.X.; Sun, Z.Z.; Feng, W.J.; Zhang, J.W. Physico-mechanical properties changes of Qinghai-Tibet clay due to cyclicfreezeing and thawing. Chin. J. Rock Mech. Eng. 2005, 24, 4313–4319. [Google Scholar]
- Ai, K.M.; Zhou, K.P.; Hu, J.H.; Li, J.L.; Liu, F.P. The mechanical properties of tailings in response to environmental test. Min. Metall. Eng. 2014, 34, 4–8. [Google Scholar]
- Jiao, Y.L.; Li, R.; Zhao, L.; Wu, T.H.; Xiao, Y.; Hu, G.J.; Qiao, Y.P. Processes of soil thawing-freezing and features of soil moisture migration in the permafrost active layer. J. Glaciol. Geocryol. 2014, 36, 237–247. [Google Scholar]
- Luo, D.L.; Jin, H.J.; Lv, L.Z.; Wu, Q.B. Temporal and Spatial Characteristics of Permafrost Active Layer and Seasonal Frozen Soil Freezing and Thawing Process in the Source Region of the Yellow River. Chin. Sci. Bull. 2014, 59, 1327–1336. [Google Scholar] [CrossRef]
- Yang, S.H.; Wu, T.H.; Li, R.; Zhu, X.F.; Wang, W.H.; Yu, W.J.; Qin, Y.H.; Hao, J.M. Spatial-temporal Changes of the Near-surface Soil Freeze-thaw Status over the Qinghai-Tibetan Plateau. Plateau Meteorol. 2018, 37, 43–53. [Google Scholar]
- Luo, D.L.; Jin, H.J.; He, R.X.; Yang, S.Z. Responses of surface vegetation on soil temperature and moisture of the active layer in the source area of the yellow river. Earth Sci. 2014, 39, 421–430. [Google Scholar]
- Zhou, Y.W.; Guo, D.X.; Qiu, G.Q.; Wang, Y.T. Geocryology in China; Science Press: Beijing, China, 2000; pp. 92–360. [Google Scholar]
- Wang, S.L.; Zhao, X.F.; Guo, D.X.; Huang, Y.Z. Response of permafrost to climate change in the Qinghai-Xizang Plateau. J. Glaciol. Geocryol. 1996, 18, 157–165. [Google Scholar]
- Anna, H.; Kerri, C. Interactions between plants and Soil Microbes may alter the Relative Importance of Intraspecific and Interspecific Plant Competition in a Changing Climate. AoB Plants 2018, 10, ply039. [Google Scholar]
- Bardgett, R.D.; Leemans, D.K.; Cook, R.; Hobbs, P.J. Seasonality of soil biota of grazed and ungrazed hill grasslands. Soil Biol. Biochem. 1997, 29, 1285–1294. [Google Scholar] [CrossRef]
- Yang, A.C.; Lv, J.; Lu, J.J. Bacteria Isolated from Soil Samples of Hoh Xil-Tanggula Mountains. J. Shihezi Univ. (Nat. Sci.) 2012, 30, 545–550. [Google Scholar]
- Zhang, B.G.; Zhang, W.; Liu, G.X.; Chen, T.; Wang, L.; Zhang, G.S.; Wu, X.K.; Tan, X.S.; Long, H.R.; Mao, W.L. Effect of freeze-thaw cycles on the soil bacterial communities in different ecosystem soils in the Tibetan Plateau. J. Glaciol. Geocryol. 2012, 34, 1499–1507. [Google Scholar]
- Lipson, D.A.; Schmidt, S.K. Seasonal changes in an alpine soil bacterial community in the Colorado Rocky Mountains. Appl. Environ. Microbiol. 2004, 70, 2867–2879. [Google Scholar] [CrossRef]
- Costello, E.K.; Schmidt, S.K. Microbial diversity in alpine tundra wet meadow soil: Novel Chloroflexi from a cold, water-satu-rated environment. Environ. Microbiol. 2006, 8, 1471–1486. [Google Scholar] [CrossRef] [PubMed]
- Kaper, J.B.; Nataro, J.P.; Mobley, H.L. Pat hogenic Escherichia coli. Nat. Rev. Microbiol. 2004, 2, 123–140. [Google Scholar] [CrossRef] [PubMed]
- Oliver, J.E. Intergovernmental Panel in Climate Change (IPCC). Encycl. Energy Nat. Resour. Environ. Econ. 2013, 26, 48–56. [Google Scholar]
- Gittel, A.; Bárta, J.; Kohoutová, I.; Mikutta, R.; Owens, S.; Gilbert, J.; Urich, T. Distinct microbial communities associated with buried soils in the Siberian tundra. ISME J. 2013, 8, 841–853. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
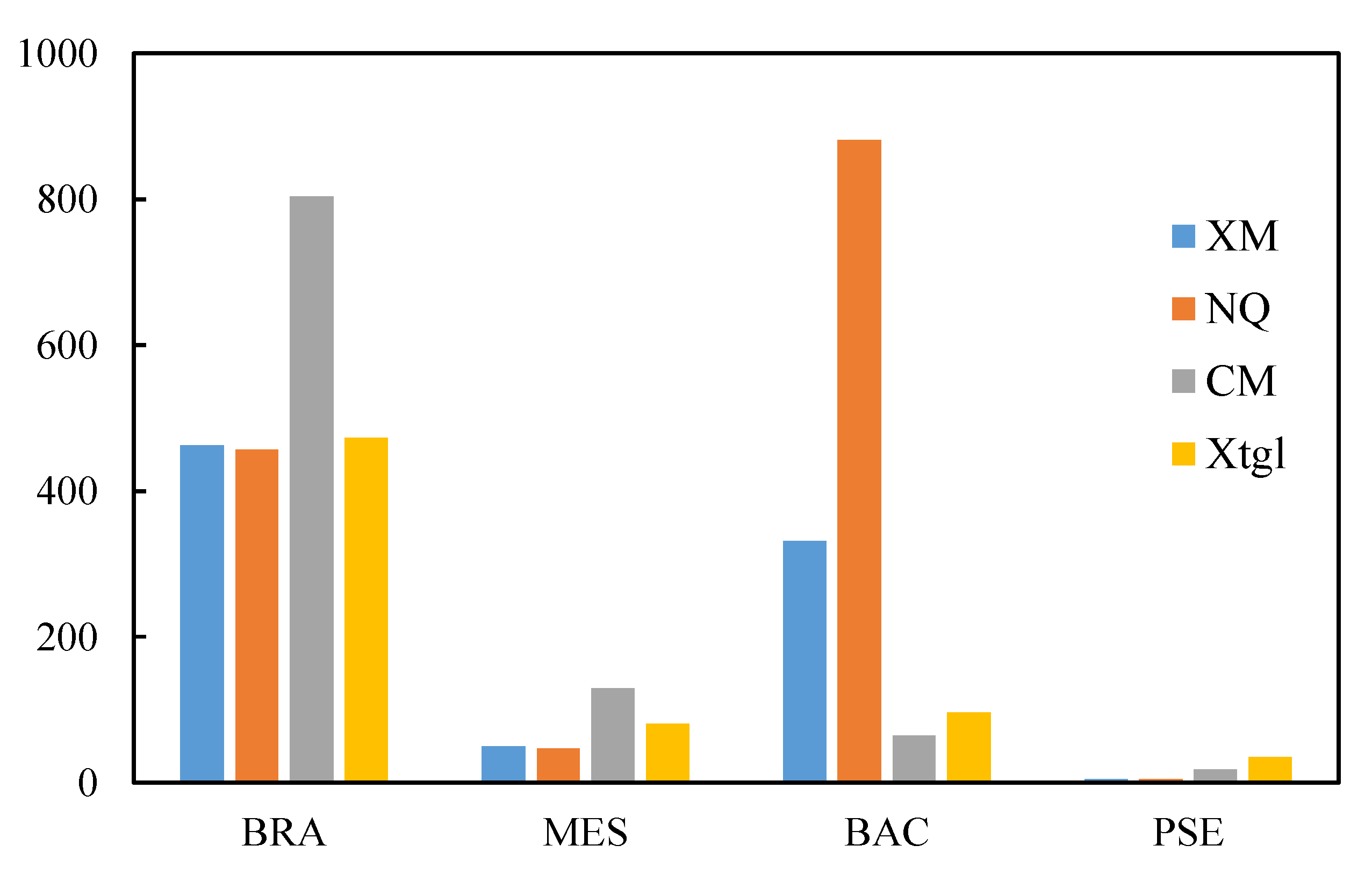{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sites | Altitude (m) | Vegetation Coverage (%) | Soil Bulk Density (g/cm3) | Soil Porosity (%) |
|---|---|---|---|---|
| Xtgl | 5050 | 10 | 1.34 | 67.93 |
| CM | 4760 | 70 | 1.26 | 79.13 |
| CN | 4580 | 90 | 1.30 | 72.63 |
| ZR | 4660 | 60 | 1.32 | 62.20 |
| BG | 4560 | 20 | 1.51 | 55.53 |
| NQ | 4560 | 35 | 1.29 | 51.67 |
| XM | 4730 | 60 | 1.29 | 71.77 |
| DS | 4550 | 80 | 1.38 | 46.40 |
| LM | 4640 | 90 | 1.49 | 63.60 |
| KM | 4710 | 80 | 1.37 | 43.50 |
| Monitoring Sites | Starting Time | Closing Time | NFTC | FTCD | FTCF |
|---|---|---|---|---|---|
| XM | 9 January 2018 | 28 March 2018 | 80 | 78 | 1.02 |
| NQ | 24 February 2018 | 20 March 2018 | 26 | 25 | 1.04 |
| CM | 8 February 2018 | 8 March 2018 | 12 | 28 | 0.43 |
| Xtgl | 28 March 2018 | 22 April 2018 | 20 | 26 | 0.77 |
| Monitoring Sites | Starting Time | Closing Time | NFTC | FTCD | FTCF |
|---|---|---|---|---|---|
| XM | 16 March 2011 | 17 April 2011 | 15 | 32 | 0.47 |
| 16 March 2014 | 4 April 2014 | 10 | 20 | 0.50 | |
| 9 January 2018 | 28 March 2018 | 80 | 78 | 1.02 | |
| NQ | 1 April 2011 | 17 April 2011 | 8 | 17 | 0.47 |
| 27 February 2014 | 25 March 2014 | 19 | 27 | 0.70 | |
| 24 February 2018 | 20 March 2018 | 26 | 25 | 1.04 |
| Site | Minimum Soil Temperature | Maximum Soil Temperature | Average Soil Temperature | FTCF |
|---|---|---|---|---|
| XM | −6.6 | 16.2 | 5.4 | 1.02 |
| NQ | −9.9 | 10.7 | −1.1 | 1.04 |
| CM | −6.6 | 16.4 | 5.5 | 0.43 |
| Xtgl | −15.9 | 16.0 | 0.4 | 0.77 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Man, Z.; Weng, B.; Yang, Y.; Gong, X.; Li, M.; Yu, Z. Effects of the Freezing–Thawing Cycle Mode on Alpine Vegetation in the Nagqu River Basin of the Qinghai–Tibet Plateau. Water 2019, 11, 2122. https://doi.org/10.3390/w11102122
Man Z, Weng B, Yang Y, Gong X, Li M, Yu Z. Effects of the Freezing–Thawing Cycle Mode on Alpine Vegetation in the Nagqu River Basin of the Qinghai–Tibet Plateau. Water. 2019; 11(10):2122. https://doi.org/10.3390/w11102122
Chicago/Turabian StyleMan, Zihao, Baisha Weng, Yuheng Yang, Xiaoyan Gong, Meng Li, and Zhilei Yu. 2019. "Effects of the Freezing–Thawing Cycle Mode on Alpine Vegetation in the Nagqu River Basin of the Qinghai–Tibet Plateau" Water 11, no. 10: 2122. https://doi.org/10.3390/w11102122
APA StyleMan, Z., Weng, B., Yang, Y., Gong, X., Li, M., & Yu, Z. (2019). Effects of the Freezing–Thawing Cycle Mode on Alpine Vegetation in the Nagqu River Basin of the Qinghai–Tibet Plateau. Water, 11(10), 2122. https://doi.org/10.3390/w11102122