3.1. Experimental Results and Estimation of Steric Effect
Experimental tests were performed to understand the physical phenomena that act on the mass transfer mechanisms in a porous media. For this purpose, the monitoring of hydraulic and selectivity properties was required. Three series of tests were performed with the same membrane. Between each series of tests, the membrane was regenerated according to the protocol previously described in the Materials and Methods section. This regeneration aimed to recover the initial membrane properties.
Table 2 provides experimental results obtained for three series in terms of maximal rejection rate (selectivity property) and hydraulic permeability (hydraulic performance). For each series, the results are in chronological order. These series were chosen as the first molecule filtrated was different (
L-phenylalanine for series 1, vitamin B12 and Lysozyme for series 2, and
L-Tyrosine for the last).
The first experiments (left-hand column) were performed with a new membrane. After conditioning, the hydraulic permeability was 6.7 × 10−14 m3 m−2. The four studied molecules were filtrated one after another, from the smallest to the largest. The rejection rate increased as filtration tests were performed and, at the same time, the hydraulic permeability significantly decreased, suggesting that a part of the protein was adsorbed at the membrane surface (or in the pore), reducing mass transfer. Nevertheless, filtration of VB12 does not seem to modify the membrane hydraulic performances, and, thus, was chosen as the model solute to follow the membrane selectivity properties. The solute rejection rates obtained with the membrane still having its initial properties (i.e., membrane that had only filtrated water and vitamin B12 after regeneration) were 5% for L-phenylalanine (test 1 series 1), 5% for L-Tyrosine (test 1—series 3), 20% for Vitamin B12 (test 1 series 2), and 65% for Lysozyme (test 2—series 2). These rejection rates are in good agreement with the size of the studied molecules, but much lower than expected for a membrane with a cut-off of 1 kDa.
After these first experimental tests, the membrane permeability and the solute transmission decreased. This behavior was illustrated by the rejection rate of vitamin B12, which increased as protein or amino acid solutions were filtrated. For example, it increased from 20% to 90% after several filtration tests of lysozyme solutions (series 2), indicating that steric effect increased significantly. This can be explained by protein or amino acid adsorption at the membrane surface and or in the pore, thereby restricting the transfer of solute through the porous medium. These adsorption phenomena are not completely irreversible, since after the regeneration treatment the membrane recovered its initial properties.
The estimated mean pore radii are given in
Table 2 for series 1 (from Equation (3)). The results (calculated average pore radii) corroborate previous observations. Indeed, the average pore radius decreased as protein filtration was performed. The results obtained with the different solutes show that this current model (model A) remains unsatisfactory for estimating steric effect because the results are dependent on the studied molecule. The average pore radius estimated with vitamin B12 was systematically lower than for lysozyme and
L-phenylalanine. Bowen et al. [
14] obtained the same results for the filtration of four uncharged solutes (vitamin B12, raffinose, sucrose, and glucose). Indeed, the estimated mean pore radius can vary by a ratio of 2:1. They also observed a modification of water permeability after filtration of each solute.
Additional tests were performed with mixtures of uncharged solutes (vitamin B12 and lysozyme) in water or salted water. These tests were performed following the tests of series 1.
The results (
Table 3) show that the rejection rate of the solute was not modified by any another solute in the solution or by salt (NaCl—5 mM) in dilute solution. In the present case, it seems that the solutes in solution did not interact between them. However, in the literature, several studies [
33,
34,
35] showed an increase in neutral solute transmission in saline solutions. This phenomenon is explained by a partial dehydratation of the molecule in the membrane pore, facilitating the transport through the membrane active layer. This behavior is specifically observed in nanofiltration. In the present study, the filtrated solutes, the porous material (membrane active layer), and the pore size were different, which could explain the observed differences.
Moreover, the difference in estimated mean pore radius according to the studied molecule was confirmed. There are two possible explanations. First, the one size cylindrical pore hypothesis is a poor and inadequate representation of the porous medium. Second, the solute interacts with the membrane surface according to the chemical groups of both the solute and the active layer (van der Waals forces or acid-base interactions). To investigate the first possibility, a pore size distribution was studied. To this end, nitrogen adsorption/desorption experiments were used to measure the porous volume according to the pore size.
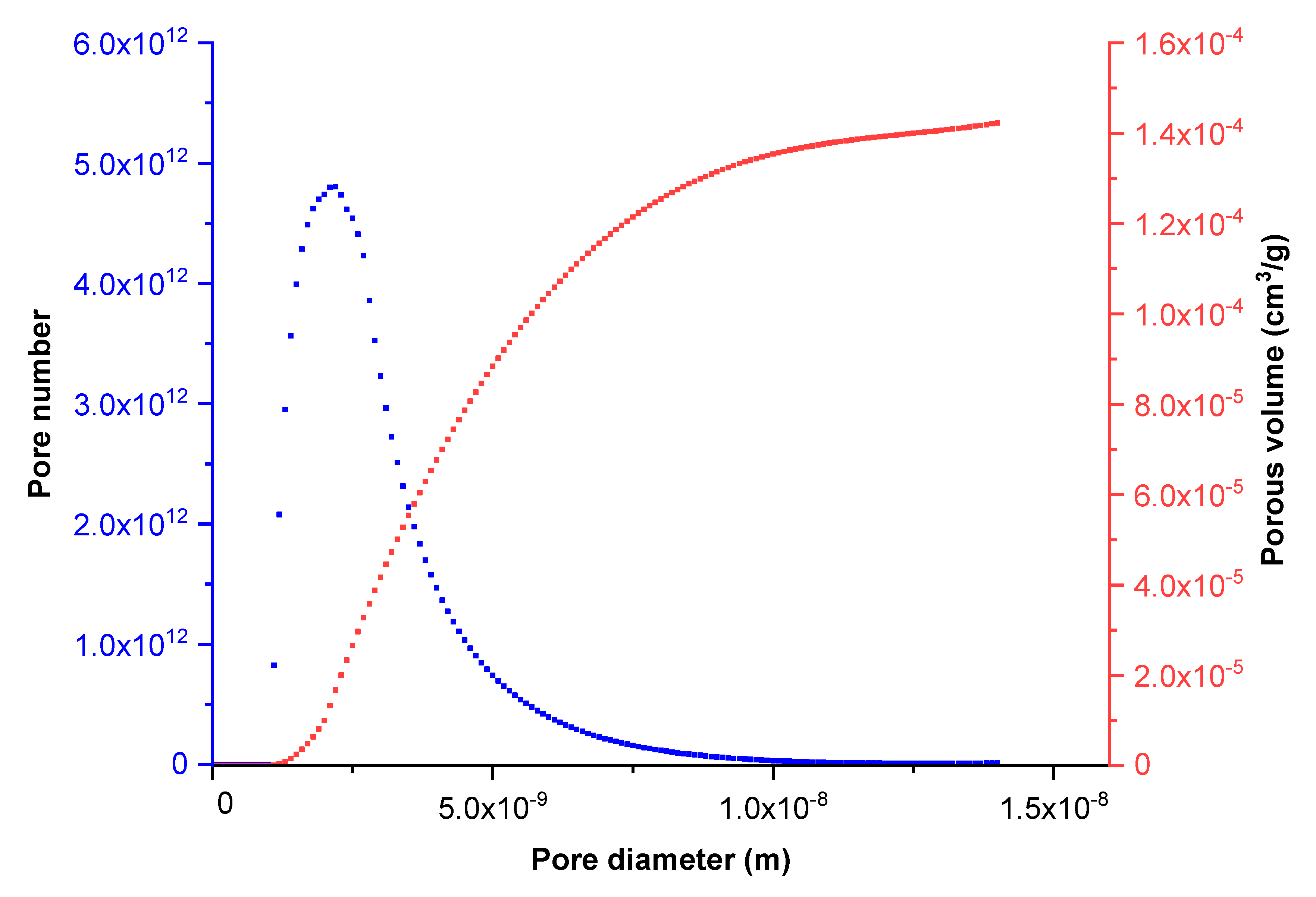
Figure 1 shows the cumulative porous volume obtained during the nitrogen adsorption step according to pore size. This curve can be suitably approximated by a log-normal distribution. These results were substantiated by Scanning Electron Microscopy (SEM) investigations. Indeed, the SEM images (
Figure 2c) show a top view of the active layer. The surface constituted sintered titania aggregates separated by nanometric cracks or channels. The larger ones, measuring about 4–10 nm, were of a lower amount than the smaller ones (less than 1 nm wide).
From these experimental results, we assumed that the porous medium was constituted of cylindrical and unidirectional pores according to the previous size distribution (
Figure 1) and calculated the pore number as a function of the pore size.
Assuming that the flow rate in each pore can be described by the Hagen–Poiseuille law, the permeation flux can be estimated with Equation (4). From these results, it is possible to estimate the active layer thickness by equaling the observed permeation flux with the flow rate calculated by Equation (4). For this purpose, the studied range of pore diameter (0–14 nm) was divided into sub-ranges of pore size from
i = 0 to 1400. The active layer thickness was around 2.9 µm. Previous observations [
28] performed by microscopy (SEM) showed that the active layer was about 1–2 µm thick.
Figure 2a shows a cross section of the membrane obtained after breaking it. The different alumina layers (different porosities) are visible below the titania active layer (full grey layer). The apparent thickness (
Figure 2c) of this layer is in agreement with previous investigation.
Calculations were performed for modelling selectivity performances of the new or regenerated membranes for the four studied molecules (i.e., for membranes, which have only been in contact with water). These calculated results were compared with the experimental observed rejection rates of the four studied solutes (
Table 4). The results are very close to the experimental ones for the four studied molecules. This comparison indicates that taking into account pore size distribution provides a good way to estimate rejection rates of neutral solutes.
3.2. Hydraulic Performance Loss and Adsorption in the Pore
The hydraulic performances of the membranes decreased as filtration tests of neutral solutes were performed (except for Vitamin B12). At the same time, selectivity performances increased, indicating an increase in the steric effect limiting the mass transfer. As shown in
Table 2 for the first series, the average pore radius (calculation relative to each filtrated molecule) decreased, step by step, until a minimum value. Four clogging/blocking mechanisms are classically reported in the literature: complete blocking (the filtrated molecule blocks the pore inlet), standard blocking (adsorption of the molecules in the pore), intermediate blocking (formation of a non-continuous layer, blocking partially the flow inlet), and a cake filtration. This last one often occurs in dead-end filtration. In our case, it can be ruled out owing to the tangential flow and associated shear stress. The intermediate blocking cannot explain both permeate flux decrease and selectivity increase. Indeed, to that end, non-continuous layers should preferentially grow, clogging the larger pore inlet only. The first mechanism, i.e., pore blocking, cannot be a plausible hypothesis, because during filtration of amino acids, the pores that clog should be the smallest, which would result in a decrease in rejection rate. So, the only plausible explanation is protein or amino acid adsorption in the larger pores, reducing the flow rate and increasing the selectivity. F. Wand et al. [
36] observed that standard blocking occurred first during ultrafiltration of colloid–water solutions (dead-end filtration experiments). K. Katsoufidou et al. [
37] observed a rapid irreversible membrane fouling during ultrafiltration of humic acid due to internal pore adsorption.
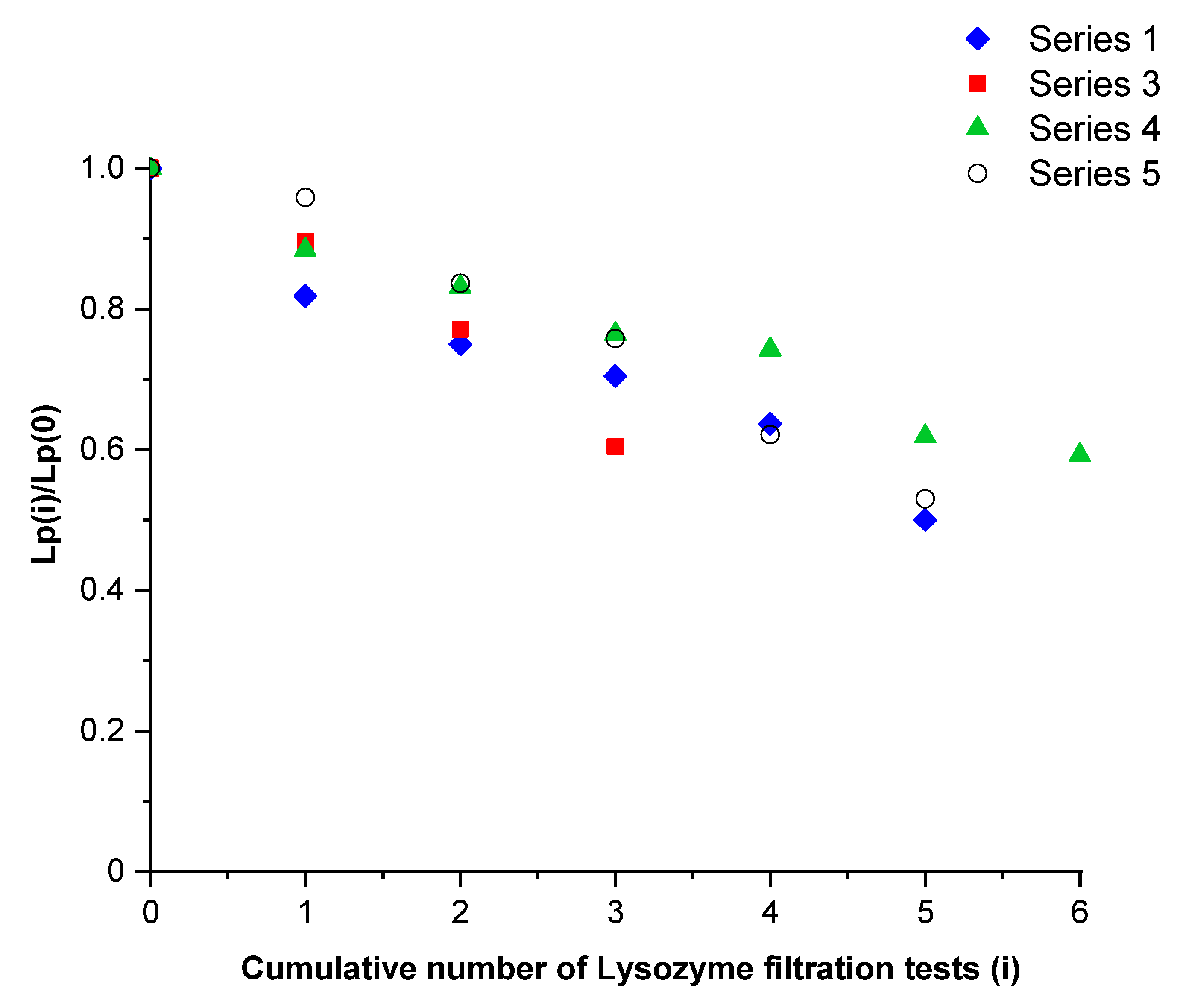
Figure 3 shows the hydraulic permeability of the membrane divided by the one obtained just before the first filtration test of lysozyme for series 1 and 3 (presented in
Table 2). These results were compared with two test series performed in the same operating conditions with another membrane (series 4 and 5). The hydraulic permeability decreased as filtration tests were carried out (quasi linear behavior), indicating the same adsorption kinetics, regardless of the studied membrane and its past experiments. Assuming one size pore distribution, the hydraulic permeability was proportional to the fourth power of the pore radius (Poiseuille flow). In these conditions, the pore radius estimated by the filtration tests of a neutral solute should vary according to the fourth-root dependence of the number of tests.
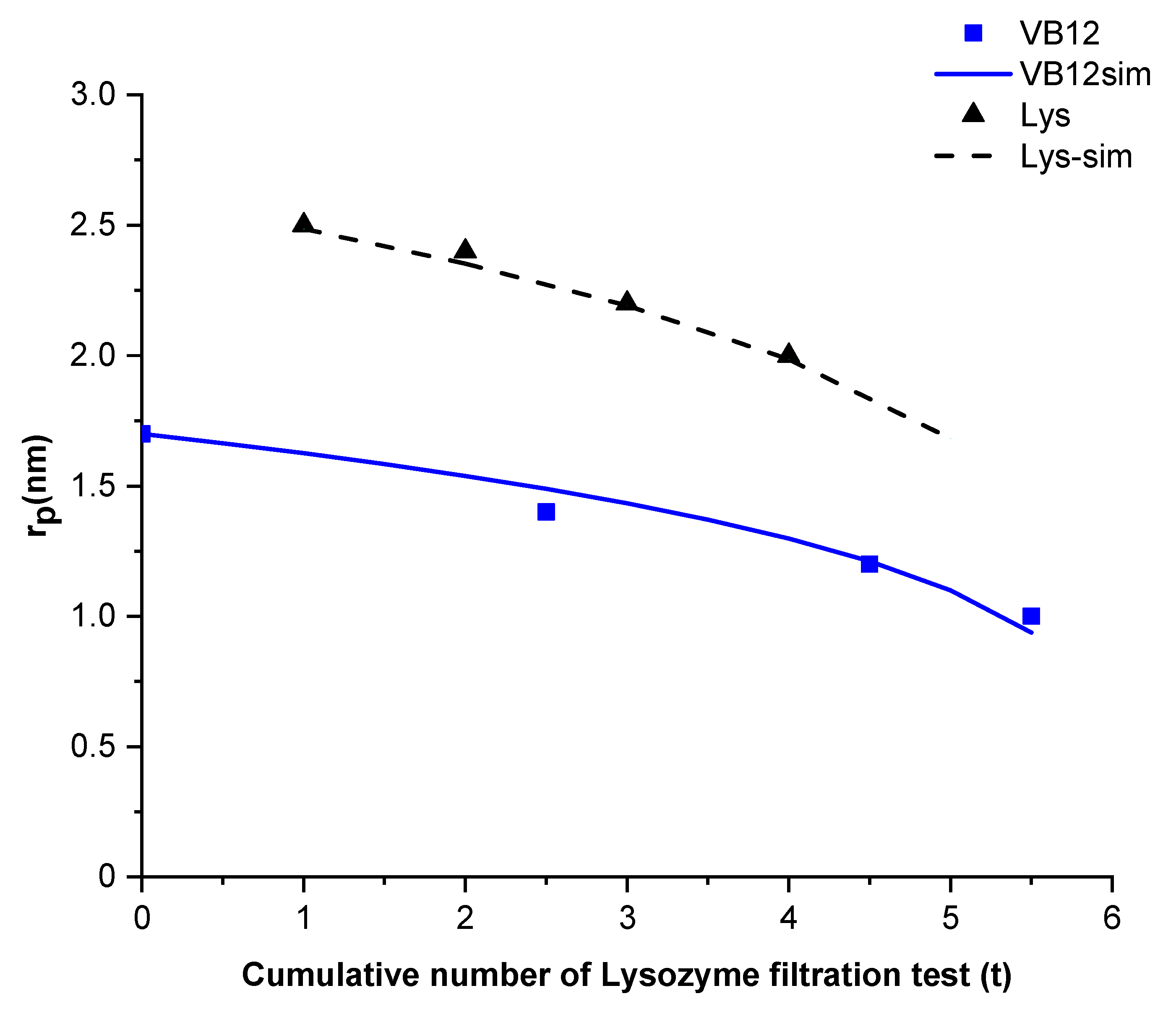
Figure 4 compares the estimated mean pore radius for vitamin B12 and Lysozyme filtration tests (series 1) with the mean pore radius calculated using a linear function of the fourth-root of the cumulative number of lysozyme test. For the two cases, the estimated pore radius can be approximated by
.
The same investigation with pore size distribution was not possible owing to the number of adjustable parameters (i.e., Np(i) and rp(i)).
Steric hindrance after protein adsorption is required in order to predict the selectivity performances of the membrane. Classical models of adsorption or adsorption kinetics do not provide information about flow restriction in pores.
In these conditions, to model these adsorption phenomena, we considered the adsorption of spherical molecules to be by uniform layers in the pore, reducing its radius of two stokes radius of the adsorbed molecule. This assumption is an arbitrary but required hypothesis for calculating the pore size distribution after molecule adsorption. The active layer thickness and the total number of pores were assumed to be unchanged (only their sizes can be modified). The calculation was carried out in all the pores with diameters twice larger than the molecule size. This procedure was numerically repeated until the calculated mass flow rate equaled the experimental permeation flux. When the equality of experimental and numerical hydraulic performances was reached, the model became predictive and capable of calculating the rejection rates of other filtrated molecules (i.e., Vitamin B12, L-Tyrosine, L-Phenylalanine, and Lysozyme).
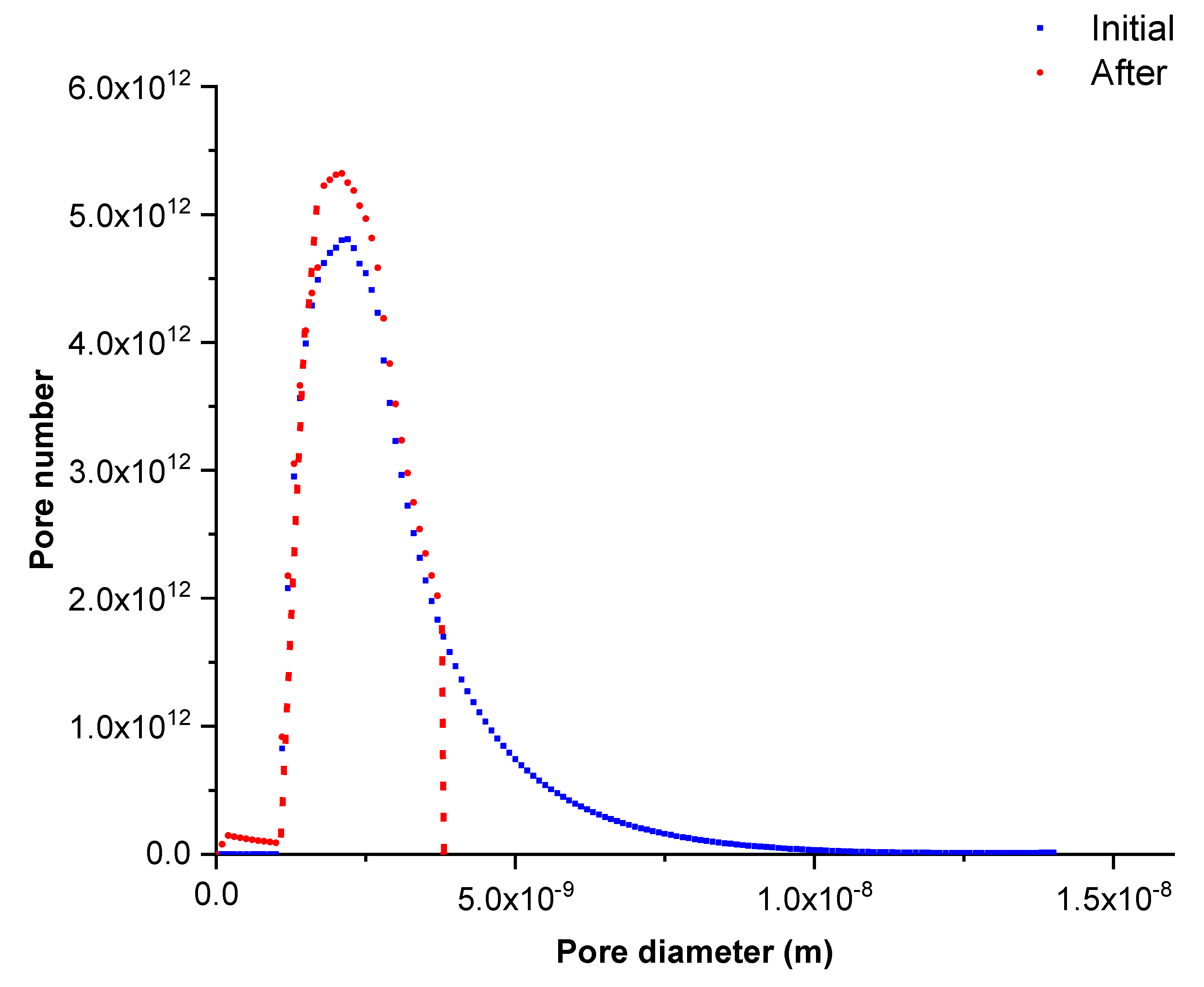
Figure 5 shows the pore size distribution of the membrane after several filtration tests of lysozyme solution (series 1) compared to the initial one. This pore size distribution was obtained by equaling the experimental and calculated permeation fluxes. The results show that adsorption phenomena took place in the larger pores, partially clogging them and reducing their apparent diameters. Consequently, the pores with an initial size higher than 3.9 nm (corresponding to pores larger than adsorbed solute) completely disappeared and the pore quantity with a diameter in between the 1.8–2.8 nm range significantly increased.
To validate this modeling, rejection rates of the four filtrated molecules were calculated before and after lysozyme adsorption.
Experimental and calculated rejection rates of the studied molecules are given in
Table 5 for the membrane before (initial properties) and after several filtration tests of lysozyme solutions. Taking into account that a pore size distribution provides a good description of membrane selectivity performances and its modification over time, from this new size distribution, the rejection rates of
L-phenylalanine,
L-Tyrosine, and Lysozyme increased and are in good agreement with the experimental results. Nevertheless, the rejection rate of vitamin B12 increased up to 60%, which is rather different regarding the experimental results (between 85% and 90%). These results are basically prior image and need to be further investigated and confirmed by other membranes and molecules.







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}