Effects of Drying and Re-Wetting on Litter Decomposition and Nutrient Recycling: A Manipulative Experiment

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
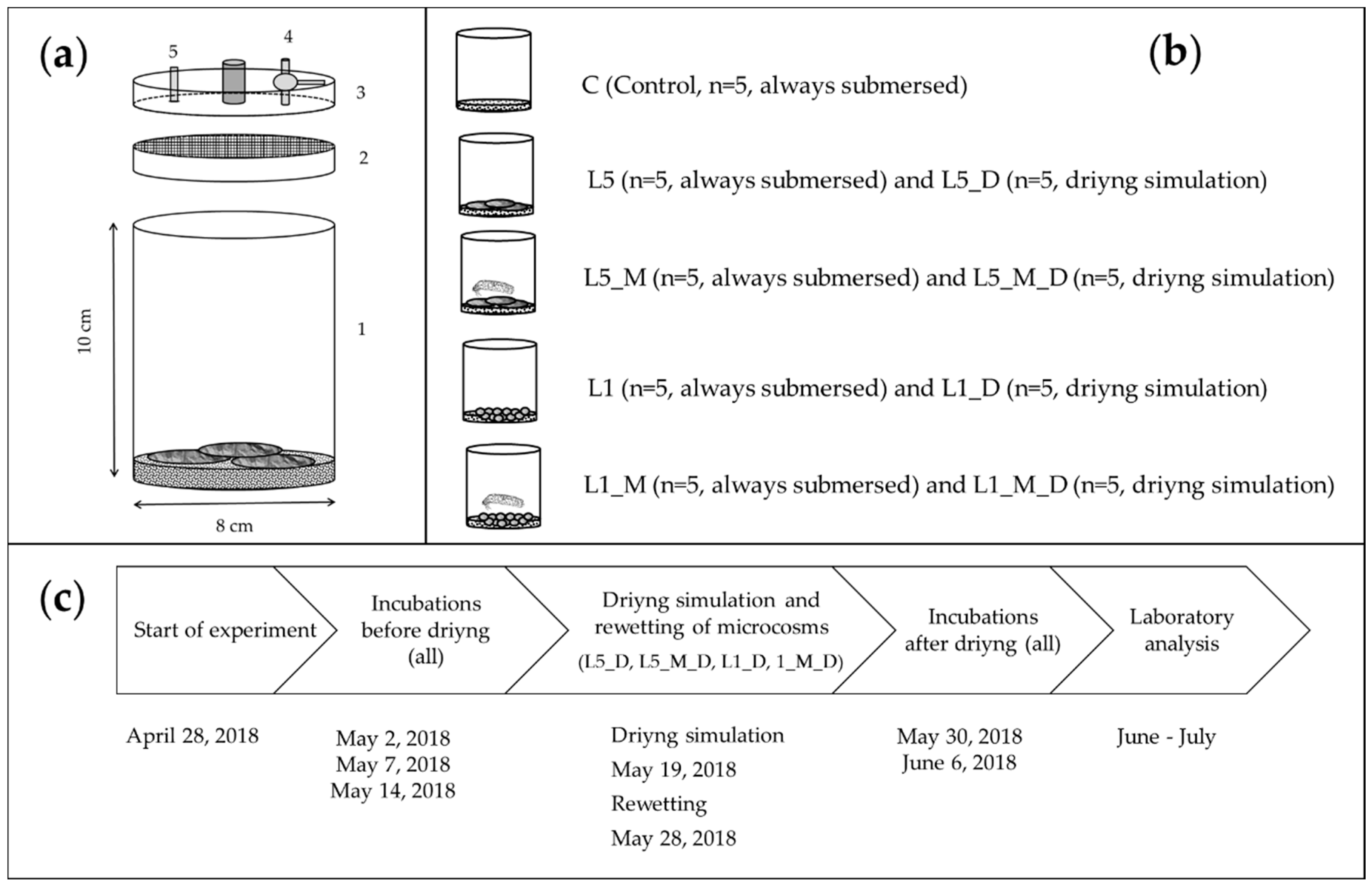
2.1. Experimental Set-Up
2.2. Leaves and Shredders
2.3. Measurement of Benthic Fluxes and Shredders Metabolism
2.4. Elemental Analysis
2.5. Data Analyses
3. Results
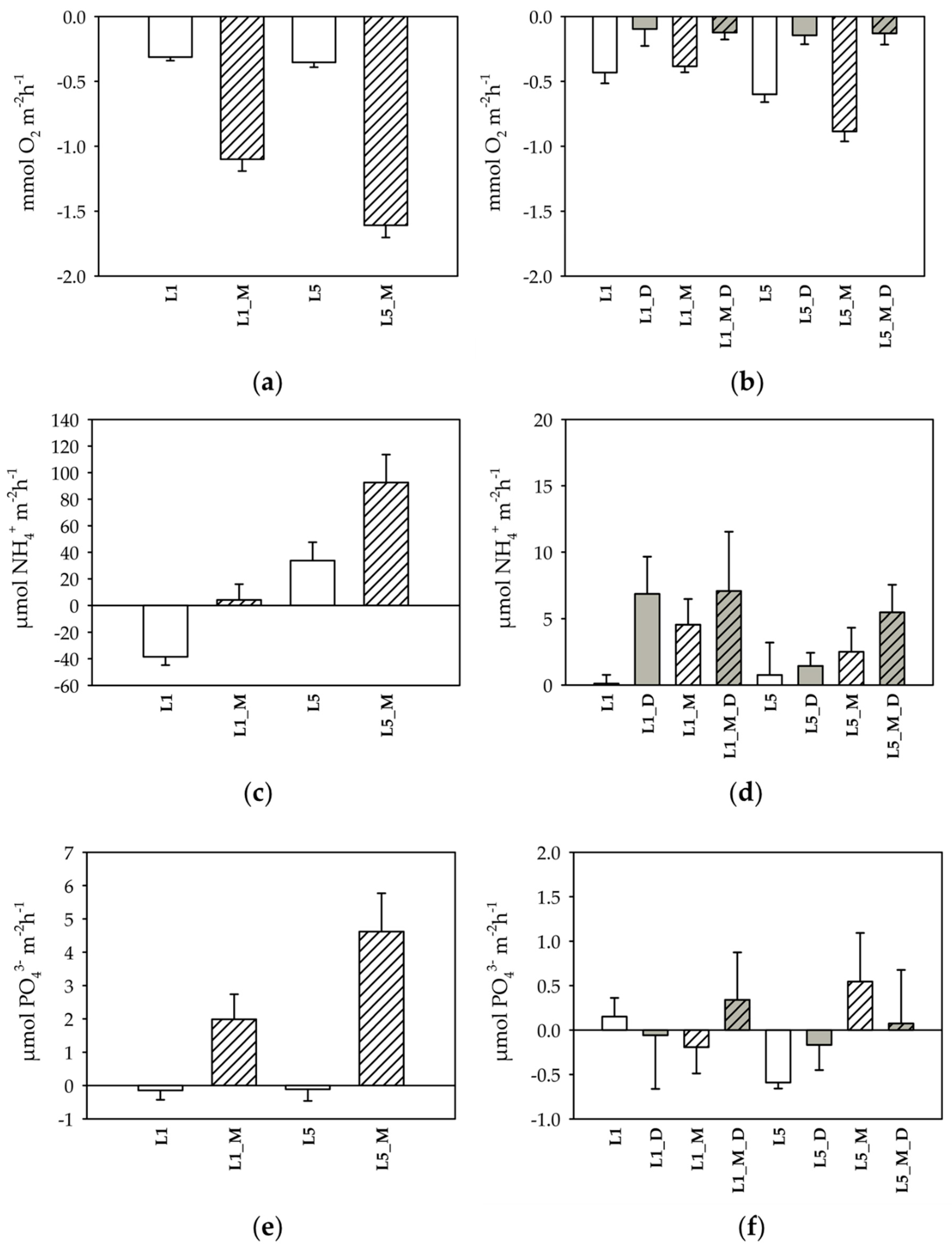
3.1. Measurements of Shredders Respiration and Excretion Rates
3.2. Benthic Respiration and Nutrient Fluxes
3.3. Residual Biomass and Its Elemental Composition
4. Discussion
4.1. Submersion, Macrofauna and Large Areas Promote Leaf Litter Decomposition
4.2. What Fluxes Tell Us About Decomposition
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Poff, N.L.; Zimmerman, J.K.H. Ecological responses to altered flow regimes: A literature review to inform the science and management of environmental flows. Freshw. Biol. 2010, 55, 194–205. [Google Scholar] [CrossRef]
- Milliman, J.D.; Farnsworth, K.L.; Jones, P.D.; Xu, K.H.; Smith, L.C. Climatic and anthropogenic factors affecting river discharge to the global ocean, 1951–2000. Glob. Planet. Change 2008, 62, 187–194. [Google Scholar] [CrossRef]
- Dahm, C.N.; Baker, M.A.; Moore, D.I.; Thibault, J.R. Coupled biogeochemical and hydrological responses of streams and rivers to drought. Freshw. Biol. 2003, 48, 1219–1231. [Google Scholar] [CrossRef]
- Von Schiller, D.; Acuña, V.; Graeber, D.; Martí, E.; Ribot, M.; Sabater, S.; Timoner, X.; Tockner, K. Contraction, fragmentation and expansion dynamics determine nutrient availability in a Mediterranean forest stream. Aquat. Sci. 2011, 73, 485–497. [Google Scholar] [CrossRef]
- Power, M.E.; Holomuzki, J.R.; Lowe, R.L. Food webs in Mediterranean rivers. Hydrobiologia 2013, 719, 119–136. [Google Scholar] [CrossRef]
- Sabater, S.; Tockner, K. Effects of Hydrologic Alterations on the Ecological Quality of River Ecosystems. In Water Scarcity in the Mediterranean. The Handbook of Environmental Chemistry; Sabater, S., Barceló, D., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; Volume 8, pp. 15–39. ISBN 978-3-642-03970-6. [Google Scholar]
- Vannote, R.L.; Water, S.; Corporation, W.; Laboratories, B.N.; Cummins, K.W.; Sedell, J.R. The River Continuum Concept. Can. J. Fish. Aquat. Sci. 1980, 37, 130–137. [Google Scholar] [CrossRef]
- Webster, J.R.; Benfield, E.F.; Ehrman, T.P.; Schaeffer, M.A.; Tank, J.E.; Hutchens, J.J.; D’Angelo, D.J. What happens to allochthonous material that falls into streams? A synthesis of new and published information from Coweeta. Freshw. Biol. 1999, 41, 687–705. [Google Scholar] [CrossRef]
- Acuña, V.; Muñoz, I.; Giorgi, A.; Omella, M.; Sabater, F.; Sabater, S. Drought and postdrought recovery cycles in an intermittent Mediterranean stream: structural and functional aspects. J. North Am. Benthol. Soc. 2005, 24, 919–933. [Google Scholar] [CrossRef]
- Corti, R.; Datry, T.; Drummond, L.; Larned, S.T. Natural variation in immersion and emersion affects breakdown and invertebrate colonization of leaf litter in a temporary river. Aquat. Sci. 2011, 73, 537–550. [Google Scholar] [CrossRef]
- Mora-Gómez, J.; Duarte, S.; Cássio, F.; Pascoal, C.; Romaní, A.M. Microbial decomposition is highly sensitive to leaf litter emersion in a permanent temperate stream. Sci. Total Environ. 2018, 621, 486–496. [Google Scholar] [CrossRef] [PubMed]
- Riedl, H.L.; Marczak, L.B.; McLenaghan, N.A.; Hoover, T.M. The role of stranding and inundation on leaf litter decomposition in headwater streams. Riparian Ecol. Conserv. 2013, 1, 3–10. [Google Scholar] [CrossRef]
- Kaushik, N.K.; Hynes, H.B.N. The fate of the dead leaves that fall into streams. Arch. für Hydrobiol. 1971, 68, 465–515. [Google Scholar]
- Datry, T. Benthic and hyporheic invertebrate assemblages along a flow intermittence gradient: Effects of duration of dry events. Freshw. Biol. 2012, 57, 563–574. [Google Scholar] [CrossRef]
- Vanni, M.J. Nutrient Cycling by Animals in Freshwater Ecosystems. Annu. Rev. Ecol. Syst. 2002, 33, 341–370. [Google Scholar] [CrossRef]
- Villanueva, V.D.; Albariño, R.; Canhoto, C. Positive effect of shredders on microbial biomass and decomposition in stream microcosms. Freshw. Biol. 2012, 57, 2504–2513. [Google Scholar] [CrossRef]
- De Souza Rezende, R.; Leite, G.F.M.; Ramos, K.; Torres, I.; Tonin, A.M.; Gonçalves Júnior, J.F. Effects of litter size and quality on processing by decomposers in a tropical savannah stream. Biotropica 2018, 50, 578–585. [Google Scholar] [CrossRef]
- Gessner, M.O.; Chauvet, E.; Dobson, M.; Oikos, S.; May, F.; Gessner, M. A perspective on leaf litter breakdown in streams. Oikos 1999, 85, 377–384. [Google Scholar] [CrossRef]
- Arsuffi, T.L.; Suberkropp, K. Selective Feeding by Stream Caddisfly (Trichoptera) Detritivores on Leaves with Fungal- Colonized Patches. Nord. Soc. Oikos 1985, 45, 50–58. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Linkins, A.E. Enzymic and chemical analysis of particulate organic matter from a boreal river. Freshw. Biol. 1990, 23, 301–309. [Google Scholar] [CrossRef]
- Rinkes, Z.L.; DeForest, J.L.; Grandy, A.S.; Moorhead, D.L.; Weintraub, M.N. Interactions between leaf litter quality, particle size, and microbial community during the earliest stage of decay. Biogeochemistry 2014, 117, 153–168. [Google Scholar] [CrossRef]
- Datry, T.; Arscott, D.B.; Sabater, S. Recent perspectives on temporary river ecology. Aquat. Sci. 2011, 73, 453–457. [Google Scholar] [CrossRef]
- Pinna, M.; Basset, A. Summer drought disturbance on plant detritus decomposition processes in three River Tirso (Sardinia, Italy) sub-basins. Hydrobiologia 2004, 522, 311–319. [Google Scholar] [CrossRef]
- Schlief, J.; Mutz, M. Leaf decay processes during and after a supra-seasonal hydrological drought in a temperate lowland stream. Int. Rev. Hydrobiol. 2011, 96, 633–655. [Google Scholar] [CrossRef]
- Abril, M.; Muñoz, I.; Menéndez, M. Heterogeneity in leaf litter decomposition in a temporary Mediterranean stream during flow fragmentation. Sci. Total Environ. 2016, 553, 330–339. [Google Scholar] [CrossRef] [PubMed]
- Schlief, J.; Mutz, M. Effect of sudden flow reduction on the decomposition of alder leaves (Alnus glutinosa [L.] Gaertn.) in a temperate lowland stream: A mesocosm study. Hydrobiologia 2009, 624, 205–217. [Google Scholar] [CrossRef]
- Arroita, M.; Flores, L.; Larrañaga, A.; Chauvet, E.; Elosegi, A. Hydrological contingency: Drying history affects aquatic microbial decomposition. Aquat. Sci. 2018, 80, 1–12. [Google Scholar] [CrossRef]
- Amalfitano, S.; Fazi, S.; Zoppini, A.; Barra Caracciolo, A.; Grenni, P.; Puddu, A. Responses of benthic bacteria to experimental drying in sediments from Mediterranean temporary rivers. Microb. Ecol. 2008, 55, 270–279. [Google Scholar] [CrossRef] [PubMed]
- Bonada, N.; Rieradevall, M.; Prat, N.; Resh, V.H. Benthic macroinvertebrate assemblages and macrohabitat connectivity in Mediterranean-climate streams of northern California. J. North Am. Benthol. Soc. 2006, 25, 32–43. [Google Scholar] [CrossRef]
- Mora-Gómez, J. Leaf Litter Decomposition in Mediterranean Streams: Microbial Porcesses and Responses to Drought under Current Global Change Scenario. Ph.D. Thesis, University of Girona, Girona, Spain, 2014. [Google Scholar]
- Mariluan, G.D.; Villanueva, V.D.; Albariño, R.J. Leaf litter breakdown and benthic invertebrate colonization affected by seasonal drought in headwater lotic systems of Andean Patagonia. Hydrobiologia 2015, 760, 171–187. [Google Scholar] [CrossRef]
- Fritz, K.M.; Feminella, J.W.; Colson, C.; Lockaby, B.G.; Governo, R.; Rummer, R.B. Biomass and decay rates of roots and detritus in sediments of intermittent Coastal Plain streams. Hydrobiologia 2006, 556, 265–277. [Google Scholar] [CrossRef]
- Otto, C.; Svensson, B.S. The Significance of Case Material Selection for the Survival of Caddis Larvae. J. Anim. Ecol. 1980, 49, 855–865. [Google Scholar] [CrossRef]
- Dalsgaard, T.; Nielsen, L.P.; Brotas, V.; Viaroli, P.; Underwood, G.J.C.; Nedwell, D.; Sundbäck, K.; Rysgaard, S.; Miles, A.; Bartoli, M.; et al. Protocol Handbook for NICE—Nitrogen Cycling in Estuaries: A Project under the EU Research Programme: Marine Science and Technology (MAST III); Dalsgaard, T., Ed.; National Environmental Research Institute: Silkeborg, Denmark, 2000; ISBN 8777725352. [Google Scholar]
- Golterman, H.L.; Clymo, R.S.; Ohnstadt, M.A.M. Methods for Physical and Chemical Analysis of Freshwaters, 2nd ed.; IBP Handbook; Blackwell Scientific: Oxford, UK, 1978; Volume 8. [Google Scholar]
- Bower, C.E.; Holm-Hansen, T. A salicylate-hypochlorite method for determining ammonia in seawater. Can. J. Fish. Aquat. Sci. 1980, 37, 794–798. [Google Scholar] [CrossRef]
- Aspila, K.I.; Agemian, H.; Chau, A.S.Y. A semi-automated method for the determination of inorganic, organic and total phosphate in sediments. Analyst 1976, 101, 187–197. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Hall, R.O.; Koch, B.J.; Marshall, M.C.; Taylor, B.W.; Tronstad, L.M. How body size mediates the role of animals in nutrient cycling in aquatic ecosystems. In Body Size: The Structure and Function of Aquatic Ecosystems; Hildrew, A.G., Edmonds-Brown, R., Raffaelli, D.G., Eds.; Cambridge University Press: New York, NY, USA, 2007; pp. 286–305. ISBN 9780511611223. [Google Scholar]
- Leberfinger, K.; Bohman, I.; Herrmann, J. Drought impact on stream detritivores: Experimental effects on leaf litter breakdown and life cycles. Hydrobiologia 2010, 652, 247–254. [Google Scholar] [CrossRef]
- Dehedin, A.; Maazouzi, C.; Puijalon, S.; Marmonier, P.; Piscart, C. The combined effects of water level reduction and an increase in ammonia concentration on organic matter processing by key freshwater shredders in alluvial wetlands. Glob. Chang. Biol. 2013, 19, 763–774. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, V.; Chauvet, E. Synergistic effects of water temperature and dissolved nutrients on litter decomposition and associated fungi. Glob. Chang. Biol. 2011, 17, 551–564. [Google Scholar] [CrossRef]
- Mas-Martí, E.; Romaní, A.M.; Muñoz, I. Consequences of warming and resource quality on the stoichiometry and nutrient cycling of a stream shredder. PLoS One 2015, 10, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Lake, P.S. Ecological effects of perturbation by drought in flowing waters. Freshw. Biol. 2003, 48, 1161–1172. [Google Scholar] [CrossRef]
- Datry, T.; Larned, S.T.; Tockner, K. Intermittent rivers: A challenge for freshwater ecology. Bioscience 2014, 64, 229–235. [Google Scholar] [CrossRef]
- Tzoraki, O.; Nikolaidis, N.P.; Amaxidis, Y.; Skoulikidis, N.T. In-stream biogeochemical processes of a temporary river. Environ. Sci. Technol. 2007, 41, 1225–1231. [Google Scholar] [CrossRef] [PubMed]
- Austin, A.T.; Vivanco, L. Plant litter decomposition in a semi-arid ecosystem controlled by photodegradation. Nature 2006, 442, 555–558. [Google Scholar] [CrossRef]
- Foulquier, A.; Artigas, J.; Pesce, S.; Datry, T. Drying responses of microbial litter decomposition and associated fungal and bacterial communities are not affected by emersion frequency. Freshw. Sci. 2015, 34, 1233–1244. [Google Scholar] [CrossRef]
- Bruder, A.; Chauvet, E.; Gessner, M.O. Litter diversity, fungal decomposers and litter decomposition under simulated stream intermittency. Funct. Ecol. 2011, 25, 1269–1277. [Google Scholar] [CrossRef]
- Langhans, S.D.; Tockner, K. The role of timing, duration, and frequency of inundation in controlling leaf litter decomposition in a river-floodplain ecosystem (Tagliamento, northeastern Italy). Oecologia 2006, 147, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Graça, M.A.S. The Role of Invertebrates on Leaf Litter Decomposition in Streams—A Review. Int. Rev. Hydrobiol. 2001, 86, 383–393. [Google Scholar] [CrossRef]
- Cummins, K.W. Structure and Function of Stream Ecosystems. Bioscience 1974, 24, 631–641. [Google Scholar] [CrossRef]
- Bärlocher, F.; Schweizer, M.; Barlocher, F. Effects of Leaf Size and Decay Rate on Colonization by Aquatic Hyphomycetes. Oikos 1983, 41, 205. [Google Scholar] [CrossRef]
- Ruginis, T.; Bartoli, M.; Petkuviene, J.; Zilius, M.; Lubiene, I.; Laini, A.; Razinkovas-Baziukas, A. Benthic respiration and stoichiometry of regenerated nutrients in lake sediments with Dreissena polymorpha. Aquat. Sci. 2014, 76, 405–417. [Google Scholar] [CrossRef]
- Melillo, J.M.; Naiman, R.J.; Aber, J.D.; Linkins, A.E. Factors Controlling Mass Loss and Nitrogen Dynamics of Plant Litter Decaying in Northern Streams. Bull. Mar. Sci. 1984, 35, 341–356. [Google Scholar]
- Chauvet, E. Changes in the chemical composition of alder, poplar and willow leaves during decomposition in a river. Hydrobiologia 1987, 148, 35–44. [Google Scholar] [CrossRef]
- Webster, J.R.; Newbold, J.D.; Thomas, S.A.; Valett, H.M.; Mulholland, P.J. Nutrient Uptake and Mineralization during Leaf Decay in Streams—A Model Simulation. Int. Rev. Hydrobiol. 2009, 94, 372–390. [Google Scholar] [CrossRef]
- Martínez, A.; Pérez, J.; Molinero, J.; Sagarduy, M.; Pozo, J. Effects of flow scarcity on leaf-litter processing under oceanic climate conditions in calcareous streams. Sci. Total Environ. 2015, 503–504, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Wilson, J.S.; Baldwin, D.S. Exploring the ‘Birch effect’ in reservoir sediments: influence of inundation history on aerobic nutrient release. Chem. Ecol. 2008, 24, 379–386. [Google Scholar] [CrossRef]
- Datry, T.; Foulquier, A.; Corti, R.; von Schiller, D.; Tockner, K.; Mendoza-Lera, C.; Clément, J.C.; Gessner, M.O.; Moleón, M.; Stubbington, R.; et al. A global analysis of terrestrial plant litter dynamics in non-perennial waterways. Nat. Geosci. 2018, 11, 497–503. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Df | O2 | NH4+ | PO43− | ||||
|---|---|---|---|---|---|---|---|
| p-Value | F Value | p-Value | F Value | p-Value | F Value | ||
| Macrofauna | 1 | <0.001 | 216.5 | <0.001 | 13.2 | <0.001 | 22.6 |
| Leaf size | 1 | <0.001 | 15.7 | <0.001 | 33.1 | 0.07 | 3.4 |
| Macrofauna: leaf size | 1 | <0.01 | 11.4 | 0.6 | 0.3 | 0.08 | 3.2 |
| Residuals | 36 | ||||||
| Df | O2 | NH4+ | PO43− | ||||
|---|---|---|---|---|---|---|---|
| p-Value | F Value | p-Value | F Value | p-Value | F Value | ||
| Macrofauna | 1 | 0.2 | 1.4 | 0.3 | 1.1 | 0.3 | 1.3 |
| Leaf size | 1 | <0.01 | 11.9 | 0.4 | 0.6 | 0.8 | 0.09 |
| Drying | 1 | <0.001 | 73.9 | 0.02 | 5.8 | 0.8 | 0.05 |
| Macrofauna: leaf size | 1 | 0.2 | 1.9 | 0.8 | 0.06 | 0.3 | 1.1 |
| Macrofauna: drying | 1 | 0.3 | 1.1 | 0.6 | 0.3 | 0.9 | 0.02 |
| Drying: leaf size | 1 | <0.01 | 8.6 | 0.2 | 1.8 | 0.8 | 0.09 |
| Macrofauna: drying: leaf size | 1 | 0.08 | 3.2 | 0.6 | 0.3 | 0.2 | 1.8 |
| Residuals | 32 | ||||||
| Leaves | Leaves Biomass | C (%) | C (g m−2) | N (%) | N (g m−2) | P (%) | P (g m−2) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Df | p-Value | F Value | p-Value | F Value | p-Value | F Value | p-Value | F Value | p-Value | F Value | p-Value | F Value | p-Value | F Value | |
| Macrofauna | 1 | <0.001 | 32 | 0.5 | 0.4 | <0.001 | 48 | 0.4 | 0.6 | <0.001 | 103 | 0.9 | 0.01 | <0.001 | 24 |
| Leaf size | 1 | 0.07 | 3.4 | <0.001 | 24 | <0.001 | 18.1 | 0.06 | 3.6 | 0.2 | 2 | 0.2 | 2.2 | 0.06 | 3.6 |
| Drying | 1 | <0.001 | 13.1 | 0.3 | 1.2 | <0.001 | 13.6 | <0.001 | 53.4 | 0.4 | 0.8 | <0.001 | 30 | 0.2 | 1.8 |
| Macrofauna : leaf size | 1 | 0.3 | 1.2 | 0.4 | 0.6 | 0.1 | 2.2 | 0.9 | 0.007 | 0.02 | 6 | 0.3 | 1.2 | 0.6 | 0.3 |
| Macrofauna: drying | 1 | 0.9 | 0.005 | 0.7 | 0.2 | 0.6 | 0.3 | 1 | 0.001 | 0.2 | 2.1 | 0.004 | 9.4 | 0.07 | 3.5 |
| Drying: leaf size | 1 | 0.4 | 0.7 | 0.8 | 0.1 | 0.1 | 2.2 | 0.2 | 2 | 0.8 | 0.1 | 0.5 | 0.5 | 0.9 | 0.02 |
| Macrofauna: drying: leaf size | 1 | 0.4 | 0.7 | 0.8 | 0.1 | 0.4 | 0.7 | 0.5 | 0.6 | 0.7 | 0.1 | 0.8 | 0.09 | 0.9 | 0.02 |
| Residuals | 32 | ||||||||||||||
| C:N | C:P | N:P | |
|---|---|---|---|
| Initial litter | 104 ± 5 | 3933 ± 450 | 38 ± 4 |
| L1 | 38 ± 2 | 1721 ± 183 | 46 ± 6 |
| L1_D | 43 ± 1 | 1729 ± 114 | 40 ± 2 |
| L1_M | 36 ± 2 | 1497 ± 71 | 42 ± 2 |
| L1_M_D | 43 ± 1 | 2291 ± 157 | 54 ± 5 |
| L5 | 39 ± 2 | 2124 ± 144 | 55 ± 3 |
| L5_D | 50 ± 3 | 2566 ± 284 | 51 ± 3 |
| L5_M | 39 ± 2 | 1826 ± 190 | 46 ± 4 |
| L5_M_D | 50 ± 2 | 3185 ± 64 | 64 ± 11 |
| Debris | C (g m−2) | N (g m−2) | P (g m−2) | ||||
|---|---|---|---|---|---|---|---|
| Df | p-Value | F Value | p-Value | F Value | p-Value | F Value | |
| Macrofauna | 1 | <0.001 | 49.3 | <0.001 | 26.6 | 0.5 | 0.4 |
| Leaf size | 1 | 0.02 | 5.6 | 0.5 | 0.4 | <0.001 | 39 |
| Drying | 1 | <0.001 | 17.2 | <0.001 | 35.2 | 0.09 | 3 |
| Macrofauna: leaf size | 1 | 0.09 | 3 | 0.4 | 0.9 | 0.6 | 0.2 |
| Macrofauna: drying | 1 | 0.3 | 1.2 | 0.4 | 0.8 | 0.04 | 4.6 |
| Drying: leaf size | 1 | 0.09 | 3 | 0.8 | 0.1 | 0.6 | 0.3 |
| Macrofauna: drying: leaf size | 1 | 0.2 | 1.6 | 0.03 | 5.5 | 0.4 | 0.8 |
| Residuals | 31 | ||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palmia, B.; Bartoli, M.; Laini, A.; Bolpagni, R.; Ferrari, C.; Viaroli, P. Effects of Drying and Re-Wetting on Litter Decomposition and Nutrient Recycling: A Manipulative Experiment. Water 2019, 11, 708. https://doi.org/10.3390/w11040708
Palmia B, Bartoli M, Laini A, Bolpagni R, Ferrari C, Viaroli P. Effects of Drying and Re-Wetting on Litter Decomposition and Nutrient Recycling: A Manipulative Experiment. Water. 2019; 11(4):708. https://doi.org/10.3390/w11040708
Chicago/Turabian StylePalmia, Beatrice, Marco Bartoli, Alex Laini, Rossano Bolpagni, Claudio Ferrari, and Pierluigi Viaroli. 2019. "Effects of Drying and Re-Wetting on Litter Decomposition and Nutrient Recycling: A Manipulative Experiment" Water 11, no. 4: 708. https://doi.org/10.3390/w11040708
APA StylePalmia, B., Bartoli, M., Laini, A., Bolpagni, R., Ferrari, C., & Viaroli, P. (2019). Effects of Drying and Re-Wetting on Litter Decomposition and Nutrient Recycling: A Manipulative Experiment. Water, 11(4), 708. https://doi.org/10.3390/w11040708