Assessing the Functional Response to Streamside Fencing of Pastoral Waikato Streams, New Zealand


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites and Design
2.2. Physical Habitat
2.3. Functional Indicators
2.3.1. Organic Matter Retention
2.3.2. Ecosystem Metabolism
2.3.3. Organic Matter Processing
2.3.4. Nutrient Transformation
2.4. Data Analysis
3. Results
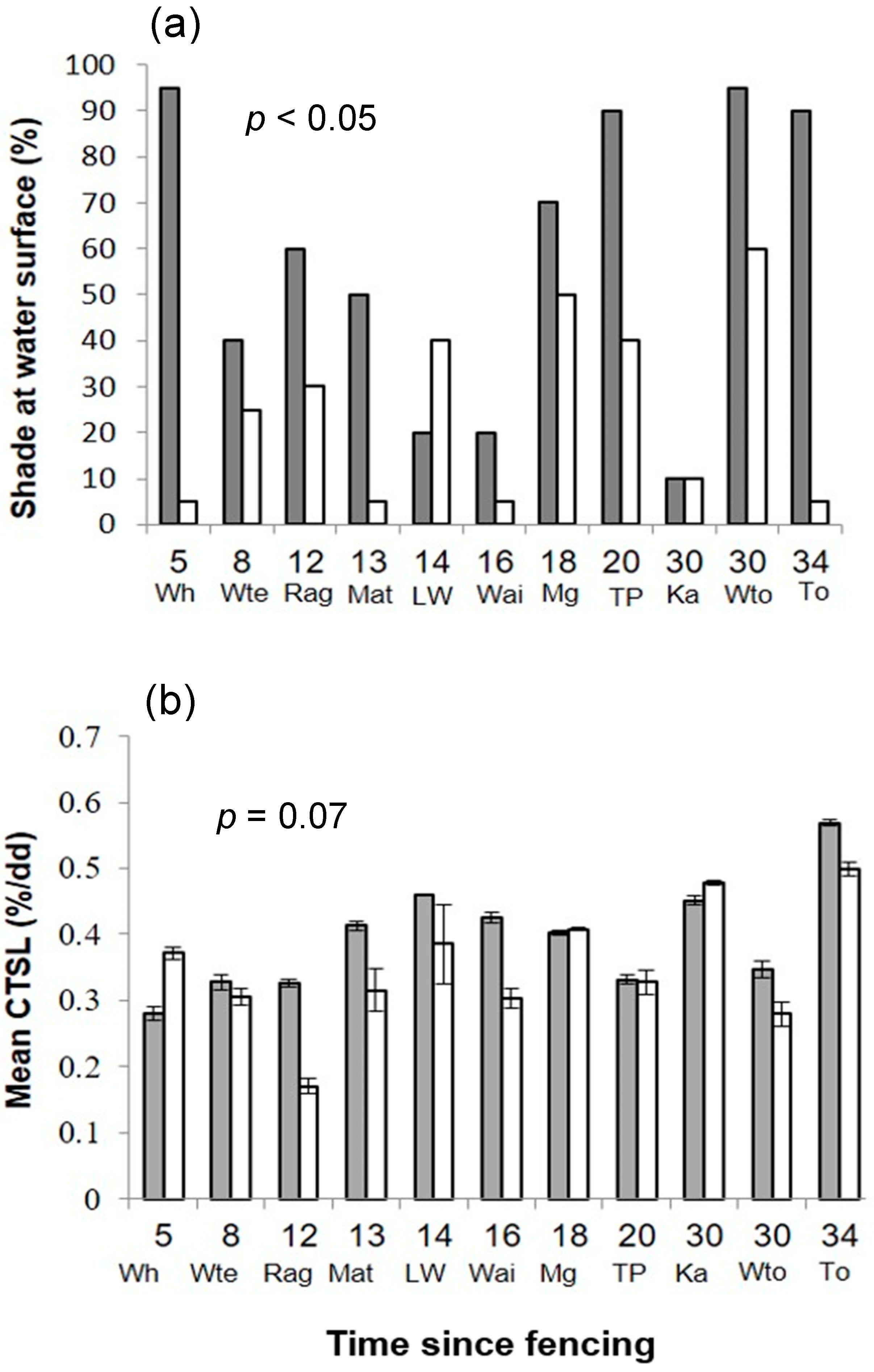
3.1. Physical Habitat
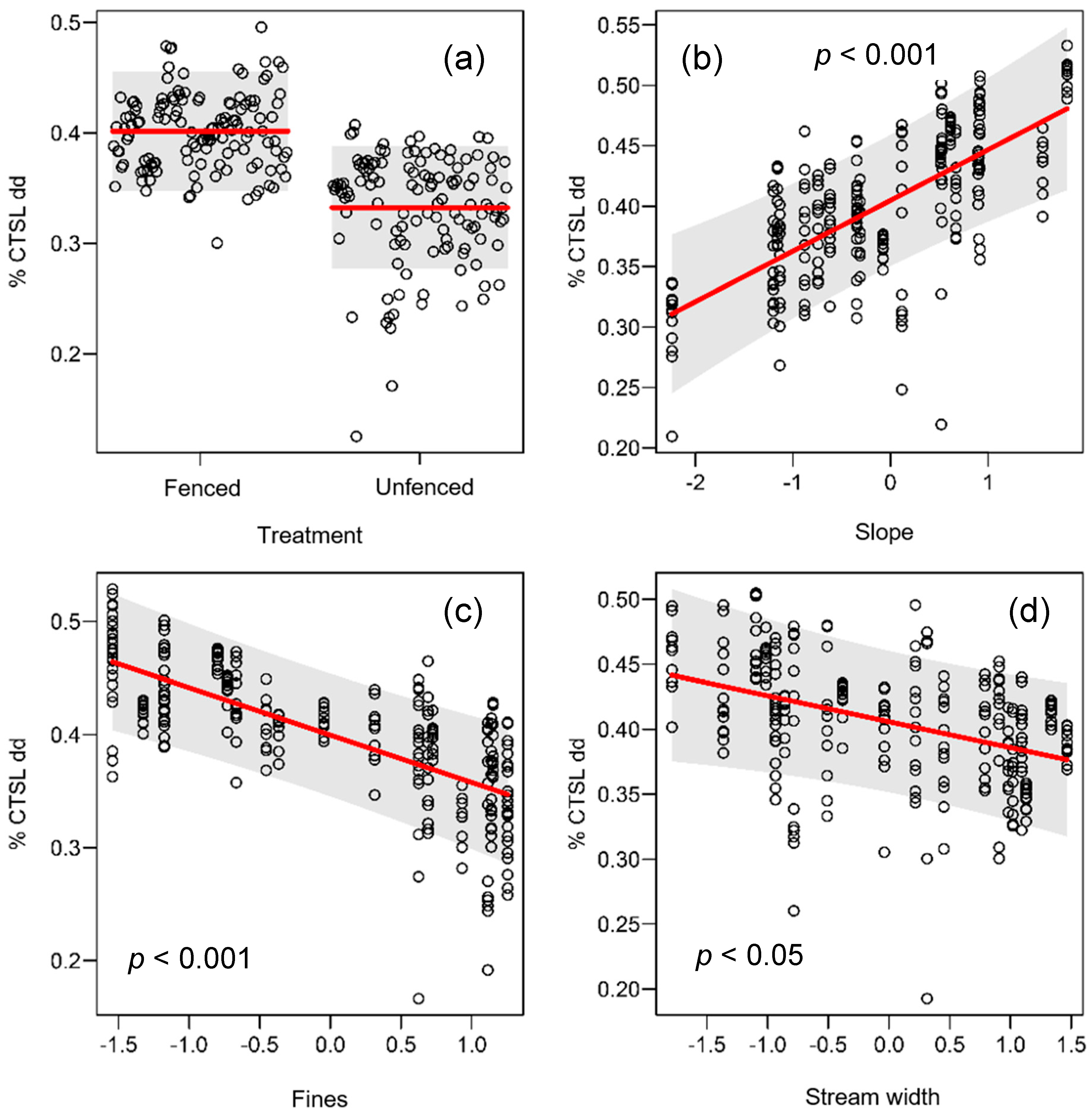
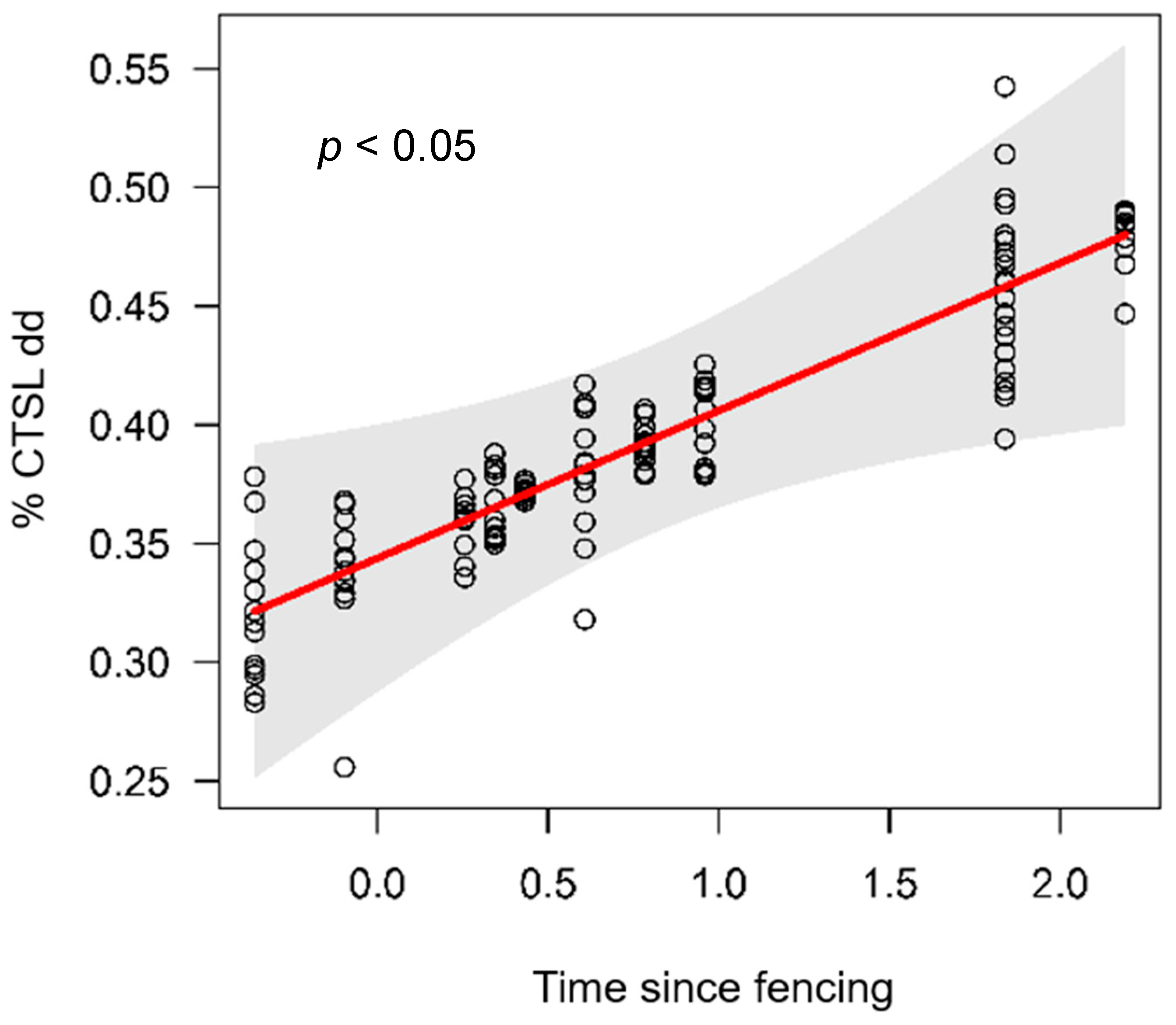
3.2. Functional Variables
3.2.1. Organic Matter Retention
3.2.2. Ecosystem Metabolism
3.2.3. Organic matter processing
3.2.4. Nutrient transformation
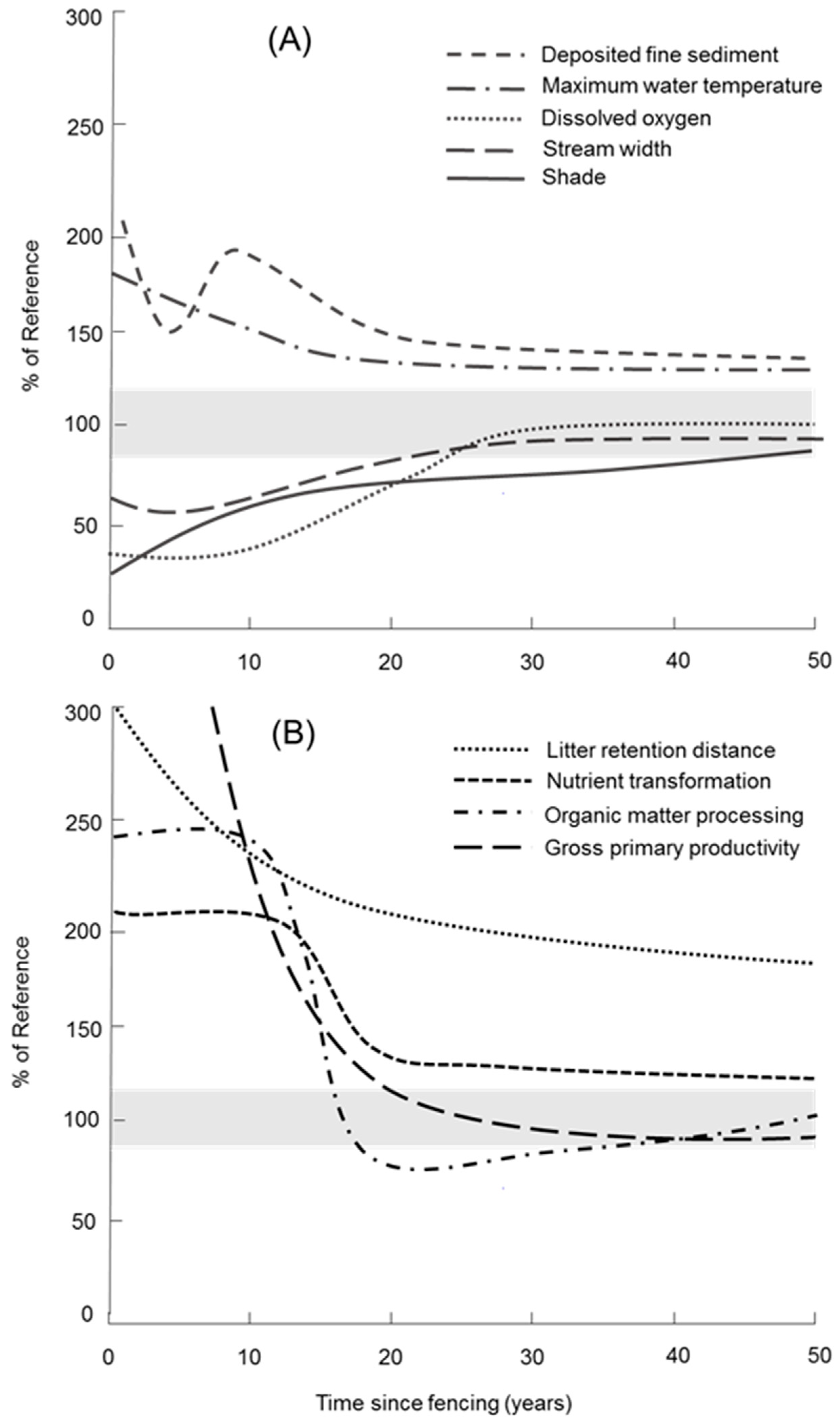
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bernhardt, E.S.; Palmer, M.A.; Allan, J.D.; Alexander, G.; Barnas, K.; Brooks, S.; Carr, J.; Clayton, S.; Dahm, C.; Follstad-Shah, J.; et al. Synthesizing U.S. River restoration efforts. Science 2005, 308, 636–637. [Google Scholar] [CrossRef] [PubMed]
- Roni, P.; Hanson, K.; Beechie, T. Global Review of the Physical and Biological Effectiveness of Stream Habitat Rehabilitation Techniques. N. Am. J. Fish. Manag. 2008, 28, 856–890. [Google Scholar] [CrossRef]
- Pedersen, M.L.; Andersen, J.M.; Nielsen, K.; Linnemann, M. Restoration of Skjern River and its valley: Project description and general ecological changes in the project area. Ecol. Eng. 2007, 30, 131–144. [Google Scholar] [CrossRef]
- ICPR. Action Plan on Floods 1995–2005—Action Targets, Implementation and Results; Brochure, Abridged Version of Technical Report No. 156; ICPR: Koblenz, Germany, 2007; p. 16. ISBN 3-935324-63-4. [Google Scholar]
- Schwarz, U. Assessment of the Restoration Potential along the Danube and MAIN Tributaries; Final Draft; WWF International, Danube-Carpathian Programme: Vienna, Austria, May 2010. [Google Scholar]
- McKergow, L.A.; Matheson, F.E.; Quinn, J.M. Riparian management: A restoration tool for New Zealand streams. Ecol. Manag. Restor. 2016, 17, 218–227. [Google Scholar] [CrossRef]
- Parkyn, S.M.; Davies-Colley, R.J.; Halliday, N.J.; Costley, K.J.; Croker, G.F. Planted riparian buffer zones in New Zealand: Do they live up to expectations? Restor. Ecol. 2003, 11, 436–447. [Google Scholar] [CrossRef]
- Craig, L.S.; Palmer, M.A.; Richardson, D.C.; Filoso, S.; Bernhardt, E.S.; Bledsoe, B.P.; Doyle, M.W.; Groffman, P.M.; Hassett, B.A.; Kaushal, S.S.; et al. Stream restoration strategies for reducing river nitrogen loads. Front. Ecol. Environ. 2008, 6, 529–538. [Google Scholar] [CrossRef] [Green Version]
- Bragina, L.; Sherlock, O.; van Rossum, A.; Jennings, E. Cattle exclusion using fencing reduces Escherichia coli (E. coli) level in stream sediment reservoirs in northeast Ireland. Agric. Ecosyst. Environ. 2017, 239, 349–358. [Google Scholar] [CrossRef]
- Palmer, M.; Hondula, K.; Koch, B. Ecological Restoration of Streams and Rivers: Shifting Strategies and Shifting Goals. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 247. [Google Scholar] [CrossRef]
- Follstad Shah, J.J.; Dahm, C.N.; Gloss, S.P.; Bernhardt, E.S. River and Riparian Restoration in the Southwest: Results of the National River Restoration Science Synthesis Project. Restor. Ecol. 2007, 15, 550–562. [Google Scholar] [CrossRef]
- Collins, K.E.; Doscher, C.; Rennie, H.G.; Ross, J.G. The Effectiveness of Riparian ‘Restoration’ on Water Quality—A Case Study of Lowland Streams in Canterbury, New Zealand. Restor. Ecol. 2013, 21, 40–48. [Google Scholar] [CrossRef]
- Lake, P.S.; Bond, N.; Reich, P. Linking ecological theory with stream restoration. Freshw. Biol. 2007, 52, 597–615. [Google Scholar] [CrossRef]
- Suding, K.N.; Hobbs, R.J. Threshold models in restoration and conservation: A developing framework. Trends Ecol. Evol. 2009, 24, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Suding, K.N. Toward an Era of Restoration in Ecology: Successes, Failures, and Opportunities Ahead. Annu. Rev. Ecol. Evol. Syst. 2011, 42, 465–487. [Google Scholar] [CrossRef] [Green Version]
- Brooks, A.; Howell, T.; Abbe, T.B.; Arthington, A. Confronting Hysteresis: Wood Based River Rehabilitation in Highly Altered Riverine Landscapes in South-Eastern Australia. Geomorphology 2006, 79, 395–422. [Google Scholar] [CrossRef]
- Rapport, D.J.; Costanza, R.; McMichael, A.J. Assessing ecosystem health. Trends Ecol. Evol. 1998, 13, 397–402. [Google Scholar] [CrossRef]
- Young, R.G.; Matthaei, C.D.; Townsend, C.R. Organic matter breakdown and ecosystem metabolism: Functional indicators for assessing river ecosystem health. J. N. Am. Benthol. Soc. 2008, 27, 605–625. [Google Scholar] [CrossRef]
- Quinn, J. Effects of Rural Land Use (Especially Forestry) and Riparian Management on Stream Habitat. N. Z. J. For. 2005, 49, 16. [Google Scholar]
- Holmes, R.; Hayes, J.; Matthaei, C.; Closs, G.; Williams, M.; Goodwin, E. Riparian management affects instream habitat condition in a dairy stream catchment. N. Z. J. Mar. Freshw. Res. 2016, 50, 581–599. [Google Scholar] [CrossRef]
- Guzha, A.C.; Rufino, M.C.; Okoth, S.; Jacobs, S.; Nóbrega, R.L.B. Impacts of land use and land cover change on surface runoff, discharge and low flows: Evidence from East Africa. J. Hydrol. Reg. Stud. 2018, 15, 49–67. [Google Scholar] [CrossRef]
- Nugroho, P.; Marsono, D.; Sudira, P.; Suryatmojo, H. Impact of Land-use Changes on Water Balance. Procedia Environ. Sci. 2013, 17, 256–262. [Google Scholar] [CrossRef] [Green Version]
- Castillo, M.M.; Morales, H.; Valencia, E.; Morales, J.J.; Cruz-Motta, J.J. The effects of human land use on flow regime and water chemistry of headwater streams in the highlands of Chiapas. Knowl. Manag. Aquat. Ecosyst. 2013. [Google Scholar] [CrossRef]
- Quinn, J.M.; Stroud, M.J. Water quality and sediment and nutrient export from New Zealand hill-land catchments of contrasting land use. N. Z. J. Mar. Freshw. Res. 2002, 36, 409–429. [Google Scholar] [CrossRef]
- Niyogi, D.K.; Simon, K.; Townsend, C.R. Breakdown of tussock grass in streams along a gradient of agricultural development in New Zealand. Freshw. Biol. 2003, 48, 1698–1708. [Google Scholar] [CrossRef]
- Von Schiller, D.; Marti, E.; Riera, J.L.; Sabater, F. Effects of nutrients and light on periphyton biomass and nitrogen uptake in Mediterranean streams with contrasting land uses. Freshw. Biol. 2007, 52, 891–906. [Google Scholar] [CrossRef]
- Violin, C.R.; Cada, P.; Sudduth, E.B.; Hassett, B.A.; Penrose, D.L.; Bernhardt, E.S. Effects of urbanization and urban stream restoration on the physical and biological structure of stream ecosystems. Ecol. Appl. A Publ. Ecol. Soc. Am. 2011, 21, 1932. [Google Scholar] [CrossRef]
- Shilla, D.J.; Shilla, D.A. The effects of catchment land use on water quality and macroinvertebrate assemblages in Otara Creek, New Zealand. Chem. Ecol. 2011, 27, 445–460. [Google Scholar] [CrossRef]
- Tank, J.L.; Rosi-Marshall, E.J.; Griffiths, N.A.; Entrekin, S.A.; Stephen, M.L. A review of allochthonous organic matter dynamics and metabolism in streams. J. N. Am. Benthol. Soc. 2010, 29, 118–146. [Google Scholar] [CrossRef] [Green Version]
- Giling, D.P.; Grace, M.R.; Mac Nally, R.; Thompson, R.M. The influence of native replanting on stream ecosystem metabolism in a degraded landscape: Can a little vegetation go a long way? Freshw. Biol. 2013, 58, 2601–2613. [Google Scholar] [CrossRef]
- Clapcott, J.E.; Young, R.G.; Goodwin, E.O.; Leathwick, J.R. Exploring the response of functional indicators of stream health to land-use gradients. Freshw. Biol. 2010, 55, 2181–2199. [Google Scholar] [CrossRef]
- Quinn, J.M.; Phillips, N.R.; Parkyn, S.M. Factors influencing retention of coarse particulate organic matter in streams. Earth Surf. Process. Landf. 2007, 32, 1186–1203. [Google Scholar] [CrossRef]
- McTammany, M.E.; Benfield, E.F.; Webster, J.R. Recovery of stream ecosystem metabolism from historical agriculture. J. N. Am. Benthol. Soc. 2007, 26, 532–545. [Google Scholar] [CrossRef]
- Webster, J.R.; Benfield, E.F. Vascular plant breakdown in freshwater ecosystems. Annu. Rev. Ecol. Syst. 1986, 17, 567–594. [Google Scholar] [CrossRef]
- Tiegs, S.D.; Clapcott, J.; Griffiths, N.; Boulton, A.J. A standardized cotton strip assay for measuring organic-matter decomposition in streams. Ecol. Indic. 2013, 32, 131–139. [Google Scholar] [CrossRef]
- Boulton, A.; Quinn, J. A simple and versatile technique for assessing cellulose decomposition potential in floodplain and riverine sediments. Arch. Fur Hydrobiol. 2000, 150, 133–151. [Google Scholar] [CrossRef]
- Griffiths, N.A.; Tiegs, S.D. Organic-matter decomposition along a temperature gradient in a forested headwater stream. Freshw. Sci. 2016, 35, 518–533. [Google Scholar] [CrossRef]
- Vyšná, V.; Dyer, F.; Maher, W.; Norris, R. Cotton strip decomposition rate as a river condition indicator–Diel temperature range and deployment season and length also matter. Ecol. Indic. 2014, 45, 508–521. [Google Scholar] [CrossRef]
- Bierschenk, A.; Savage, C.; Townsend, C.; Matthaei, C. Intensity of land use in the catchment influences ecosystem functioning along a freshwater-marine continuum. Ecosystems 2012, 15, 637–651. [Google Scholar] [CrossRef]
- Udy, J.W.; Fellows, C.S.; Bartkow, M.E.; Bunn, S.E.; Clapcott, J.E.; Harch, B.D. Measures of nutrient processes as indicators of stream ecosystem health. Hydrobiologia 2006, 572, 89–102. [Google Scholar] [CrossRef]
- Mulholland, P.J.; Webster, J. Nutrient dynamics in streams and the role of J-NABS. J. N. Am. Benthol. Soc. 2010, 29, 100–117. [Google Scholar] [CrossRef]
- Von Schiller, D.; Acuña, V.; Aristi, I.; Arroita, M.; Basaguren, A.; Bellin, A.; Boyero, L.; Butturini, A.; Ginebreda, A.; Kalogianni, E.; et al. River ecosystem processes: A synthesis of approaches, criteria of use and sensitivity to environmental stressors. Sci. Total Environ. 2017, 596–597, 465–480. [Google Scholar] [CrossRef]
- Diebel, M.W.; Zanden, M.J.V. Nitrogen stable isotopes in streams: Effects of agricultural sources and transformations. Ecol. Appl. 2009, 19, 1127–1134. [Google Scholar] [CrossRef] [PubMed]
- Udy, J.W.; Bunn, S.E. Elevated δ15N values in aquatic plants from cleared catchments: Why? Mar. Freshw. Res. 2001, 52, 347–351. [Google Scholar] [CrossRef]
- Hamilton, S.; Tank, J.; Raikow, D.; Wollheim, W.; Peterson, B.; Webster, J. Nitrogen uptake and transformation in a midwestern U.S. stream: A stable isotope enrichment study. Biogeochemistry 2001, 54, 297–340. [Google Scholar] [CrossRef]
- Rubin, Z.; Kondolf, G.M.; Rios-Touma, B. Evaluating Stream Restoration Projects: What Do We Learn from Monitoring? Water 2017, 9, 174. [Google Scholar] [CrossRef]
- Sudduth, E.B.; Hassett, B.A.; Cada, P.; Bernhardt, E.S. Testing the Field of Dreams Hypothesis: Functional responses to urbanization and restoration in stream ecosystems. Ecol. Appl. 2011, 21, 1972–1988. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, F.; Peterson, E.E.; Boone, E.L.; Sippel, S.; Bunn, S.E.; Harch, B.D. Identifying the spatial scale of land use that most strongly influences overall river ecosystem health score. Ecol. Appl. 2012, 22, 2188–2203. [Google Scholar] [CrossRef] [PubMed]
- Parkyn, S.; Collier, K.; Clapcott, J.; David, B.; Davies-Colley, R.; Matheson, F.; Quinn, J.; Shaw, W.; Storey, R. The Restoration Indicators Toolkit: Indicators for Monitoring the Ecological Success of Stream Restoration; National Institute of Water and Atmospheric Research: Hamilton, New Zealand, 2010; p. 134. Available online: http://www.envirolink.govt.nz/assets/Envirolink/RestorationIndicatorToolkit-stream.pdf (accessed on 20 June 2019).
- Leathwick, J.R.; West, D.; Gerbeaux, P.; Kelly, D.; Robertson, H.; Brown, D.; Chadderton, W.L.; Ausseil, A.-G. Freshwater Ecosystems of New Zealand (FENZ) Database; Department of Conservation: Wellington, New Zealand, 2010.
- Ministry for the Environment. The New Zealand Land Cover Database (LCDB) 4; Ministry for the Environment: Wellington, New Zealand, 2014.
- Wolman, M.G. A method of sampling coarse river-bed material. Trans. Am. Geophys. Union 1954, 35, 951–956. [Google Scholar] [CrossRef]
- Speaker, R.; Moore, K.; Gregory, S. Analysis of the process of retention of organic matter in stream ecosystems. Verh. Int. Ver. Fuer Theor. Angew. Limnol. 1984, 22, 1835–1841. [Google Scholar] [CrossRef]
- Webster, J.R.; Covich, A.P.; Tank, J.L.; Crockett, T.V. Retention of coarse organic particles in streams in the southern Appalachian Mountains. J. N. Am. Benthol. Soc. 1994, 13, 140–150. [Google Scholar] [CrossRef]
- James, A.B.W.; Henderson, I.M. Comparison of coarse particulate organic matter retention in meandering and straightened sections of a third-order New Zealand stream. River Res. Appl. 2005, 21, 641–650. [Google Scholar] [CrossRef]
- Brookshire, E.N.J.; Dwire, K.A. Controls on patterns of coarse organic particle retention in headwater streams. J. N. Am. Benthol. Soc. 2003, 22, 17–34. [Google Scholar]
- Odum, H.T. Primary production in flowing waters. Limnol. Oceanogr. 1956, 1, 102–117. [Google Scholar] [CrossRef]
- Marzolf, E.R.; Mulholland, P.J.; Steinman, A.D. Improvements to the diurnal upstream-downstream dissolved oxygen change technique for determining whole-stream metabolism in small streams. Can. J. Fish. Aquat. Sci. 1994, 51, 1591–1599. [Google Scholar] [CrossRef]
- Young, R.G.; Huryn, A.D. Effects of land use on stream metabolism and organic matter turnover. Ecol. Appl. 1999, 9, 1359–1376. [Google Scholar] [CrossRef]
- Owens, M. Measurements on non-isolated natural communities in running waters. In A Manual on Methods for Measuring Primary Production in Aquatic Environments; Vollenweider, R.A., Ed.; Blackwell Scientific Publications: Oxford, UK, 1974; pp. 111–119. [Google Scholar]
- Petersen, R.C.; Cummins, K.W. Leaf processing in a woodland stream. Freshw. Biol. 1974, 4, 343–368. [Google Scholar] [CrossRef]
- Anderson, M.J. Permutational test for univariate or multivariate analysis of variance and regression. Can. J. Fish. Aquat. Sci. 2001, 58, 626–639. [Google Scholar] [CrossRef]
- Anderson, M.J. Permutational Multivariate Analysis of Variance (PERMANOVA). In Wiley StatsRef: Statistics Reference Online; Balakrishnan, N., Colton, T., Everitt, B., Piegorsch, W., Ruggeri, F., Teugels, J.L., Eds.; Department of Statistics, University of Auckland: Auckland, New Zealand, 2017. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. PRIMER v7: User Manual/Tutorial; Primer-E: Plymouth, UK, 2015; p. 296. [Google Scholar]
- Anderson, M.J.; Gorley, R.N.; Clarke, R.K. PERMANOVA+ for Primer: Guide to Software and Statisticl Methods; Primer-E Limited: Auckland, New Zealand, 2008. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Elphick, C.S. A protocol for data exploration to avoid common statistical problems. Methods Ecol. Evol. 2010, 1, 3–14. [Google Scholar] [CrossRef]
- Nakagawa, S.; Schielzeth, H. A general and simple method for obtaining R² from Generalized Linear Mixed-effects Models. Methods Ecol. Evol. 2013, 4, 133–142. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67. [Google Scholar] [CrossRef]
- R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Clary, W.P. Stream Channel and Vegetation Responses to Late Spring Cattle Grazing. J. Range Manag. 1999, 52, 218–227. [Google Scholar] [CrossRef]
- Bernhardt, E.S.; Palmer, M.A. River restoration: The fuzzy logic of repairing reaches to reverse catchment scale degradation. Ecol. Appl. 2011, 21, 1926–1931. [Google Scholar] [CrossRef] [PubMed]
- Lepori, F.; Palm, D.; Malmqvist, B. Effects of stream restoration on ecosystem functioning: Detritus retentiveness and decomposition. J. Appl. Ecol. 2005, 42, 228–238. [Google Scholar] [CrossRef]
- Loades, K.W.; Bengough, A.G.; Bransby, M.F.; Hallett, P.D. Planting density influence on fibrous root reinforcement of soils. Ecol. Eng. 2010, 36, 276–284. [Google Scholar] [CrossRef]
- Boothroyd, I.K.G.; Quinn, J.M.; Langer, E.R.; Costley, K.J.; Steward, G. Riparian buffers mitigate effects of pine plantation logging on New Zealand streams: 1. Riparian vegetation structure, stream geomorphology and periphyton. For. Ecol. Manag. 2004, 194, 199–213. [Google Scholar] [CrossRef]
- Wagenhoff, A.; Townsend, C.R.; Matthaei, C.D. Macroinvertebrate responses along broad stressor gradients of deposited fine sediment and dissolved nutrients: A stream mesocosm experiment. J. Appl. Ecol. 2012, 49, 892–902. [Google Scholar] [CrossRef]
- Sponseller, R.A.; Benfield, E.F. Influences of land use on leaf breakdown in southern Appalachian headwater streams: A multiple-scale analysis. J. N. Am. Benthol. Soc. 2001, 20, 44–59. [Google Scholar] [CrossRef]
- Risse-Buhl, U.; Mendoza-Lera, C.; Norf, H.; Pérez, J.; Pozo, J.; Schlief, J. Contrasting habitats but comparable microbial decomposition in the benthic and hyporheic zone. Sci. Total Environ. 2017, 605–606, 683–691. [Google Scholar] [CrossRef]
- Piggott, J.J.; Niyogi, D.K.; Townsend, C.R.; Matthaei, C.D. Multiple stressors and stream ecosystem functioning: Climate warming and agricultural stressors interact to affect processing of organic matter. J. Appl. Ecol. 2015, 52, 1126–1134. [Google Scholar] [CrossRef]
- Parkyn, S.M.; Davies-Colley, R.J.; Cooper, A.B.; Stroud, M.J. Predictions of stream nutrient and sediment yield changes following restoration of forested riparian buffers. Ecol. Eng. 2005, 24, 551–558. [Google Scholar] [CrossRef]
- Simon, K.S.; Townsend, C.R.; Biggs, B.J.F.; Bowden, W.B. Temporal variation of N and P uptake in 2 New Zealand streams. J. N. Am. Benthol. Soc. 2005, 24, 1–18. [Google Scholar] [CrossRef]
- Clapcott, J.E.; Young, R.G.; Neale, M.W.; Doehring, K.A.M.; Barmuta, L.A. Land use affects temporal variation in stream metabolism. Freshw. Sci. 2016, 35, 1164–1175. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, S.K. Biogeochemical time lags may delay responses of streams to ecological restoration: Time lags in stream restoration. Freshw. Biol. 2012, 57, 43–57. [Google Scholar] [CrossRef]
- Benfield, E.; Fritz, K.; Tiegs, S. Leaf litter Breakdown. In Methods in Stream Ecology; Elsevier: Amsterdam, The Netherlands, 2017; pp. 71–82. [Google Scholar]
- Giling, D.P.; Mac Nally, R.; Thompson, R.M. How sensitive are invertebrates to riparian-zone replanting in stream ecosystems? Mar. Freshw. Res. 2016, 67, 1500–1511. [Google Scholar] [CrossRef]
- Allan, J.D.; Erickson, D.L.; Fay, J. The influence of catchment land use on stream integrity across multiple spatial scales. Freshw. Biol. 1997, 37, 149–161. [Google Scholar] [CrossRef] [Green Version]
- Ranganath, S.C.; Hession, W.; Wynn Thompson, T. Livestock Exclusion Influences on Riparian Vegetation, Channel Morphology, and Benthic Macroinvertebrate Assemblages. J. Soil Water Conserv. 2009, 64, 33–42. [Google Scholar] [CrossRef]
- Lowrance, R. Riparian forest ecosystems as filters for non-point source pollution. In Success, Limitations, and Frontiers in Ecosystem Science; Pace, M.L., Groffman, P.M., Eds.; Springer: New York, NY, USA, 1998; pp. 113–141. [Google Scholar]
- England, L.E.; Rosemond, A.D. Small reductions in forest cover weaken terrestrial-aquatic linkages in headwater streams. Freshw. Biol. 2004, 49, 721–734. [Google Scholar] [CrossRef]
- Gore, J.A.; Shields, F.D. Can large rivers be restored? Bioscience 1995, 45, 142–152. [Google Scholar] [CrossRef]
- Storey, R.G.; Cowley, D.R. Recovery of three New Zealand rural streams as they pass through native forest remnants. Hydrobiologia 1997, 353, 63–76. [Google Scholar] [CrossRef]
- Scarsbrook, M.R.; Halliday, J. Transition from pasture to native forest land-use along stream continua: Effects on stream ecosystems and implications for restoration. N. Z. J. Mar. Freshw. Res. 1999, 33, 293–310. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site (Site Code) | Treatment Fenced (F)/Unfenced (U) | Years since Fencing | SR Length (m) + SR Proportion (%) of Total Upstream River Length | SR Mean Wetted Width (m) | SR Mean Depth (m) | SR Mean Riparian Width (m) | SR Mean Discharge (L/s) | TCA (ha) + (SR Riparian Area Proportion (%) of Total Catchment Area) | Dominant Bed Substrate | Slope (%) |
|---|---|---|---|---|---|---|---|---|---|---|
| Whatawhata (Wh) | F | 5 | 560 (29.3) | 1.5 | 0.21 | 53.5 | 22.6 | 301.6 (1.73) | Silt | 0.1 |
| U | 0 | 500 (20.7) | 2.0 | 0.20 | 7.4 | 52.0 | 301.6 (0.24) | Gravel | 1.15 | |
| Waitete (Wte) | F | 8 | 900 (21.5) | 3.4 | 0.34 | 5.0 | 126.9 | 636.1 (0.03) | Cobble | 0.03 |
| U | 0 | 640 (19.5) | 5.2 | 0.33 | 0.0 | 130.3 | 636.1 (< 0.01) | Cobble | 0.17 | |
| Raglan (Rag) | F | 12 | 480 (8.5) | 1.9 | 0.15 | 36.9 | 17.9 | 619.9 (0.12) | Silt | 3.3 |
| U | 0 | 470 (7.7) | 2.0 | 0.24 | 4.9 | 14.7 | 619.9 (0.02) | Silt | 2.5 | |
| Matarawa (Mat) | F | 13 | 1100 (22.5) | 2.2 | 0.35 | 7.7 | 90.0 | 1025.3 (0.01) | Pumice | 0.10 |
| U | 0 | 1250 (33.1) | 1.8 | 0.23 | 3.7 | 69.1 | 850.9 (0.03) | Pumice | 0.46 | |
| Little Waipa (LW) | F | 14 | 670 (3.9) | 7.1 | 0.34 | 7.8 | 248.0 | 8612.5 (0.01) | Cobble | 0.27 |
| U | 0 | 994 (6.1) | 4.1 | 0.31 | 20.0 | 391.5 | 3467.2 (0.04) | Pumice | 0.73 | |
| Waitetuna (Wai) | F | 16 | 1600 (13.5) | 6.0 | 0.67 | 8.2 | 783.1 | 5568.1 (0.02) | Silt | 0.19 |
| U | 0 | 920 (8.4) | 4.4 | 0.47 | 1.8 | 604.3 | 5427.5 (0.01) | Silt | 0.11 | |
| Mangawhara (Mg) | F | 18 | 1200 (9.9) | 6.1 | 0.28 | 14.8 | 188.2 | 3151.3 (0.01) | Gravel | 0.37 |
| U | 0 | 1260 (11.5) | 6.7 | 0.27 | 0.6 | 245.0 | 3077.8 (<0.01) | Gravel | 0.82 | |
| Tapapakanga (TP) | F | 20 | 650 (0.1) | 5.8 | 0.13 | 11.4 | 140.7 | 997.4 (0.23) | Cobble | 0.87 |
| U | 0 | 1000 (18.4) | 5.5 | 0.15 | 9.9 | 173.6 | 997.4 (0.20) | Gravel | 0.12 | |
| Kakahu (Ka) | F | 30 | 1500 (11.1) | 5.7 | 0.31 | 15.3 | 604.5 | 3715.6 (0.15) | Gravel | 0.28 |
| U | 0 | 715 (0.1) | 2.8 | 0.27 | 0.0 | 151.8 | 1005.3 (< 0.01) | Gravel | 0.73 | |
| Waitomo (Wto) | F | 30 | 1200 (16.0) | 3.9 | 0.49 | 22.7 | 265.7 | 1949.6 (0.25) | Pumice | 0.15 |
| U | 0 | 1200 (17.6) | 2.6 | 0.56 | 3.6 | 287.6 | 1835.7 (0.04) | Pumice | 0.27 | |
| Taupo (To) | F | 34 | 1700 (16.0) | 2.1 | 0.18 | 75.0 | 131.5 | 3101.9 (1.02) | Gravel | 1.14 |
| U | 0 | 830 (0.1) | 1.1 | 0.21 | 0.0 | 59.6 | 387.7 (0.54) | Gravel | 0.74 |
| Metric | Variables | Fenced Sites | (Min–Max) | Unfenced Sites | (Min–Max) | T-value | p-value |
|---|---|---|---|---|---|---|---|
| Physical | Stream depth (m) | 0.31 | (0.13–0.67) | 0.29 | (0.15–0.56) | 0.77 | 0.46 |
| Physical | Stream width (m) | 4.11 | (1.50–7.10) | 3.50 | (1.10–6.70) | 1.34 | 0.21 |
| Physical | Channel width (m) | 10.10 | (3.10–28.70) | 9.00 | (4.8–15.91) | 0.47 | 0.64 |
| Physical | Discharge (m3/s) | 0.25 | (0.02–0.78) | 0.19 | (0.02–0.60) | 1.47 | 0.17 |
| Physical | Slope (%) | 0.62 | (0.03–3.28) | 0.80 | (0.11–2.45) | 1.23 | 0.25 |
| Physical | Silt (%) | 17.45 | (0.00-89.00) | 16.80 | (0.00–100.00) | 0.07 | 0.95 |
| Physical | Pumice/Sand (%) | 18.36 | (0.00–77.00) | 29.70 | (0.00–99.00) | 2.63 | 0.06 |
| Physical | Fines (%) | 37.36 | (1.00–91.00) | 43.80 | (1.00–100.00) | 0.63 | 0.55 |
| Physical | Shade (%) | 58.10 | (10.00–95.00) | 25.00 | (5.00–60.00) | 3.30 | <0.05 |
| Physical | Dissolved oxygen (% saturation) | 95.80 | (81.60–128.20) | 93.90 | (76.90–120.90) | 1.20 | 0.25 |
| Physical | Max daily water temperature (°C) | 14.60 | (10.60–20.20) | 14.80 | (10.70–21.40) | 0.76 | 0.46 |
| Functional | Retention dowel (Vdep; mm/s) | 3.50 | (0.30–14.90) | 1.60 | (0.60–4.10) | 1.40 | 0.18 |
| Functional | Retention gingko (Vdep; mm/s) | 2.10 | (0.27–5.90) | 2.10 | (0.50–6.00) | 0.08 | 0.93 |
| Functional | Retention triangle (Vdep; mm/s) | 5.90 | (0.80–18.1) | 6.50 | (1.70–14.4) | 0.43 | 0.67 |
| Functional | GPP (g O2/m2/day) | 3.10 | (0.10–10.9) | 2.10 | (0.03–5.20) | 1.02 | 0.33 |
| Functional | ER (g O2/m2/day) | 6.00 | (1.40–17.8) | 6.20 | (0.70–11.2) | 0.13 | 0.89 |
| Functional | Wood mass loss (%/degree day) | 0.07 | (0.04–0.10) | 0.08 | (0.03–0.13) | 1.40 | 0.19 |
| Functional | Cotton tensile strength loss (%/degree day) | 0.40 | (0.30–0.60) | 0.30 | (0.20–0.50) | 2.05 | 0.07 |
| Functional | Nutrient transformation (δ15N; ‰) | 5.60 | (3.40–9.10) | 5.80 | (3.20–9.90) | 0.28 | 0.78 |
| Physical | Stream Depth | Stream Width | Channel Width | Discharge | Slope | Shade | |||||||||||||
| df | MS | F | p | MS | F | p | MS | F | p | MS | F | p | MS | F | p | MS | F | p | |
| Time since fencing | 1 | 0.00 | 0.11 | 0.74 | 0.00 | 0.00 | 0.99 | 0.35 | 1.32 | 0.29 | 0.03 | 0.66 | 0.43 | 0.00 | 0.00 | 0.97 | 0.00 | 0.00 | 0.96 |
| Treatment | 1 | 0.00 | 0.48 | 0.51 | 0.07 | 1.63 | 0.23 | 0.00 | 0.00 | 1.00 | 0.01 | 2.01 | 0.18 | 0.14 | 4.04 | 0.08 | 5.43 | 8.73 | 0.01 |
| Site | 9 | 0.02 | 12.92 | 0.00 | 0.35 | 7.92 | 0.00 | 0.26 | 1.62 | 0.22 | 0.05 | 7.69 | 0.01 | 0.28 | 8.33 | 0.00 | 0.97 | 1.57 | 0.26 |
| Res | 10 | 0.00 | 0.04 | 0.16 | 0.01 | 0.03 | 0.62 | ||||||||||||
| Temperature | Max. Temperature | Silt | Pumice/Sand | Fines | |||||||||||||||
| df | MS | F | p | MS | F | p | MS | F | p | MS | F | p | MS | F | p | ||||
| Time since fencing | 1 | 25.41 | 1.49 | 0.23 | 0.01 | 1.43 | 0.28 | 14.80 | 3.52 | 0.09 | 8.88 | 2.14 | 0.16 | 0.02 | 0.01 | 0.95 | |||
| Treatment | 1 | 9.06 | 2.46 | 0.07 | 0.00 | 0.69 | 0.43 | 0.04 | 0.02 | 0.85 | 0.89 | 3.19 | 0.11 | 0.08 | 0.18 | 0.65 | |||
| Site | 9 | 17.06 | 4.62 | 0.00 | 0.01 | 4.89 | 0.01 | 4.21 | 2.89 | 0.06 | 4.15 | 14.80 | 0.00 | 3.96 | 8.85 | 0.00 | |||
| Res | 10 | 3.69 | 0.00 | 1.46 | 0.28 | 0.45 | |||||||||||||
| Functional | GPP | ER | Wood Mass Loss/dd | CTSL/dd | Stable Isotope | Dowel | |||||||||||||
| df | MS | F | p | MS | F | p | MS | F | p | MS | F | p | MS | F | p | MS | F | p | |
| Time since fencing | 1 | 0.02 | 0.02 | 0.88 | 0.17 | 0.30 | 0.61 | 0.00 | 1.01 | 0.33 | 0.02 | 5.18 | 0.04 | 0.06 | 0.88 | 0.37 | 0.19 | 0.31 | 0.62 |
| Treatment | 1 | 0.00 | 0.01 | 0.94 | 0.09 | 0.44 | 0.52 | 0.00 | 3.13 | 0.11 | 0.01 | 4.05 | 0.06 | 0.00 | 0.04 | 0.83 | 0.41 | 1.46 | 0.29 |
| Site | 9 | 0.74 | 2.88 | 0.06 | 0.56 | 2.72 | 0.06 | 0.00 | 1.30 | 0.11 | 0.00 | 3.20 | 0.04 | 0.07 | 0.82 | 0.61 | 0.61 | 2.15 | 0.13 |
| Res | 10 | 0.26 | 0.21 | 0.00 | 0.00 | 0.08 | 0.28 | ||||||||||||
| Gingko | Triangle | ||||||||||||||||||
| df | MS | F | p | MS | F | p | |||||||||||||
| Time since fencing | 1 | 0.26 | 0.45 | 0.52 | 0.56 | 1.53 | 0.25 | ||||||||||||
| Treatment | 1 | 0.00 | 0.01 | 0.94 | 0.12 | 0.93 | 0.35 | ||||||||||||
| Site | 9 | 0.57 | 5.41 | 0.01 | 0.37 | 2.87 | 0.08 | ||||||||||||
| Res | 10 | 0.11 | 0.13 | ||||||||||||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Doehring, K.; Clapcott, J.E.; Young, R.G. Assessing the Functional Response to Streamside Fencing of Pastoral Waikato Streams, New Zealand. Water 2019, 11, 1347. https://doi.org/10.3390/w11071347
Doehring K, Clapcott JE, Young RG. Assessing the Functional Response to Streamside Fencing of Pastoral Waikato Streams, New Zealand. Water. 2019; 11(7):1347. https://doi.org/10.3390/w11071347
Chicago/Turabian StyleDoehring, Katharina, Joanne E. Clapcott, and Roger G. Young. 2019. "Assessing the Functional Response to Streamside Fencing of Pastoral Waikato Streams, New Zealand" Water 11, no. 7: 1347. https://doi.org/10.3390/w11071347
APA StyleDoehring, K., Clapcott, J. E., & Young, R. G. (2019). Assessing the Functional Response to Streamside Fencing of Pastoral Waikato Streams, New Zealand. Water, 11(7), 1347. https://doi.org/10.3390/w11071347