Water Level Fluctuation Requirements of Emergent Macrophyte Typha angustifolia L.

Abstract
:1. Introduction
2. Materials and Methods
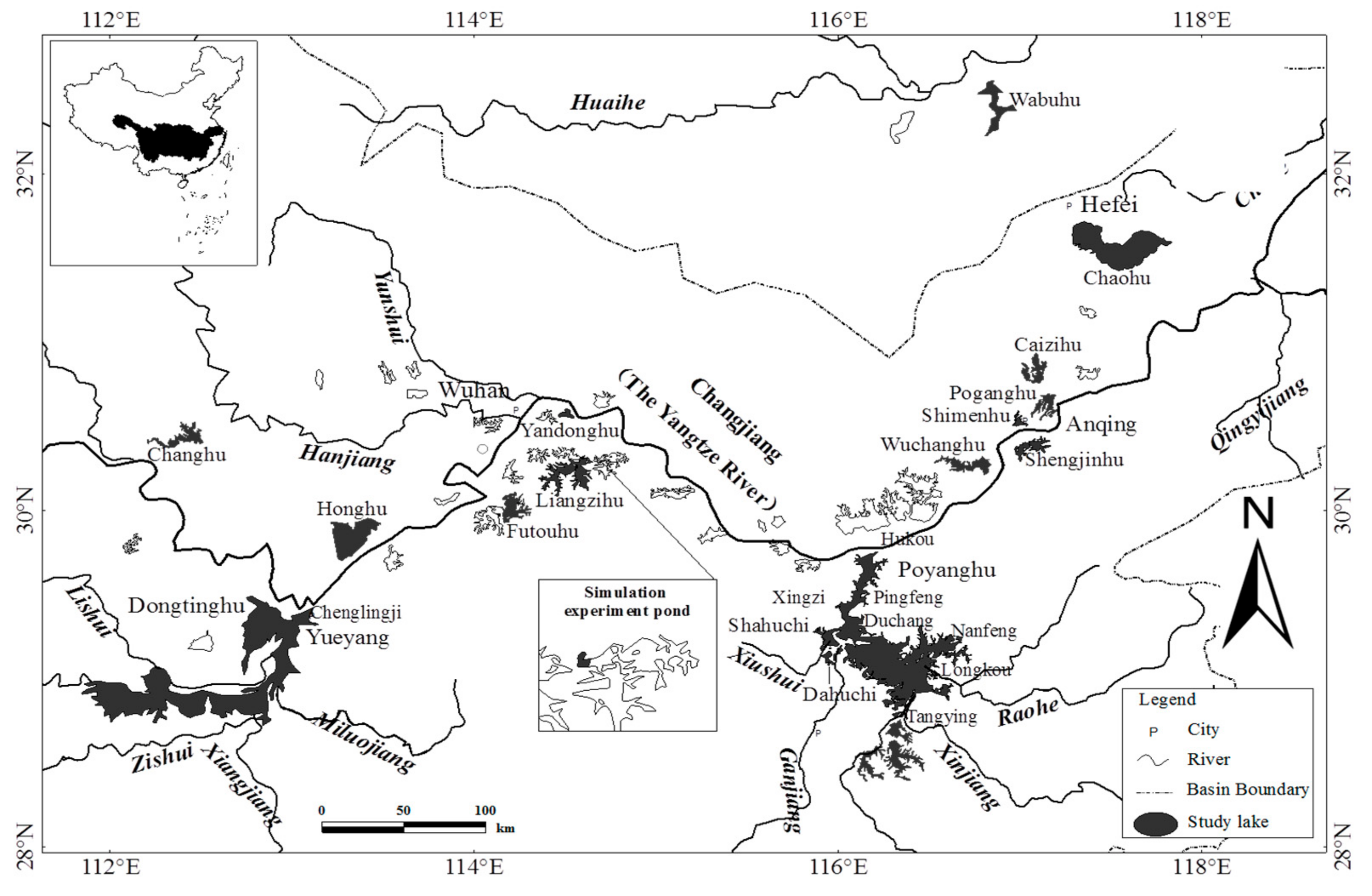
2.1. Study in Lakes
2.2. Field Investigation Method
2.3. WLFs and Meteorological Parameters
2.4. Pond Simulation Experiment
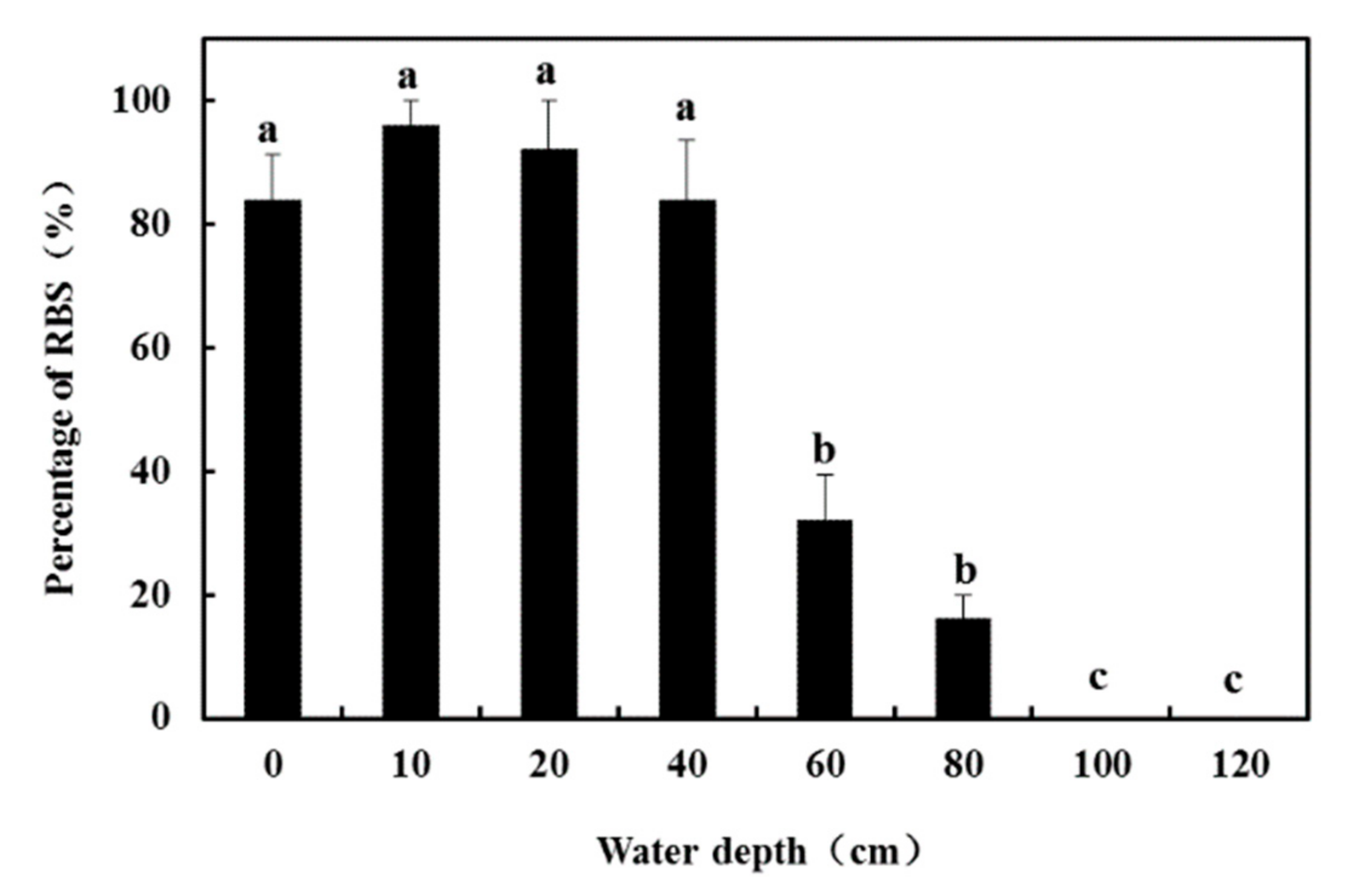
2.4.1. Experiment 1: Responses of RBS to Submergence Depth
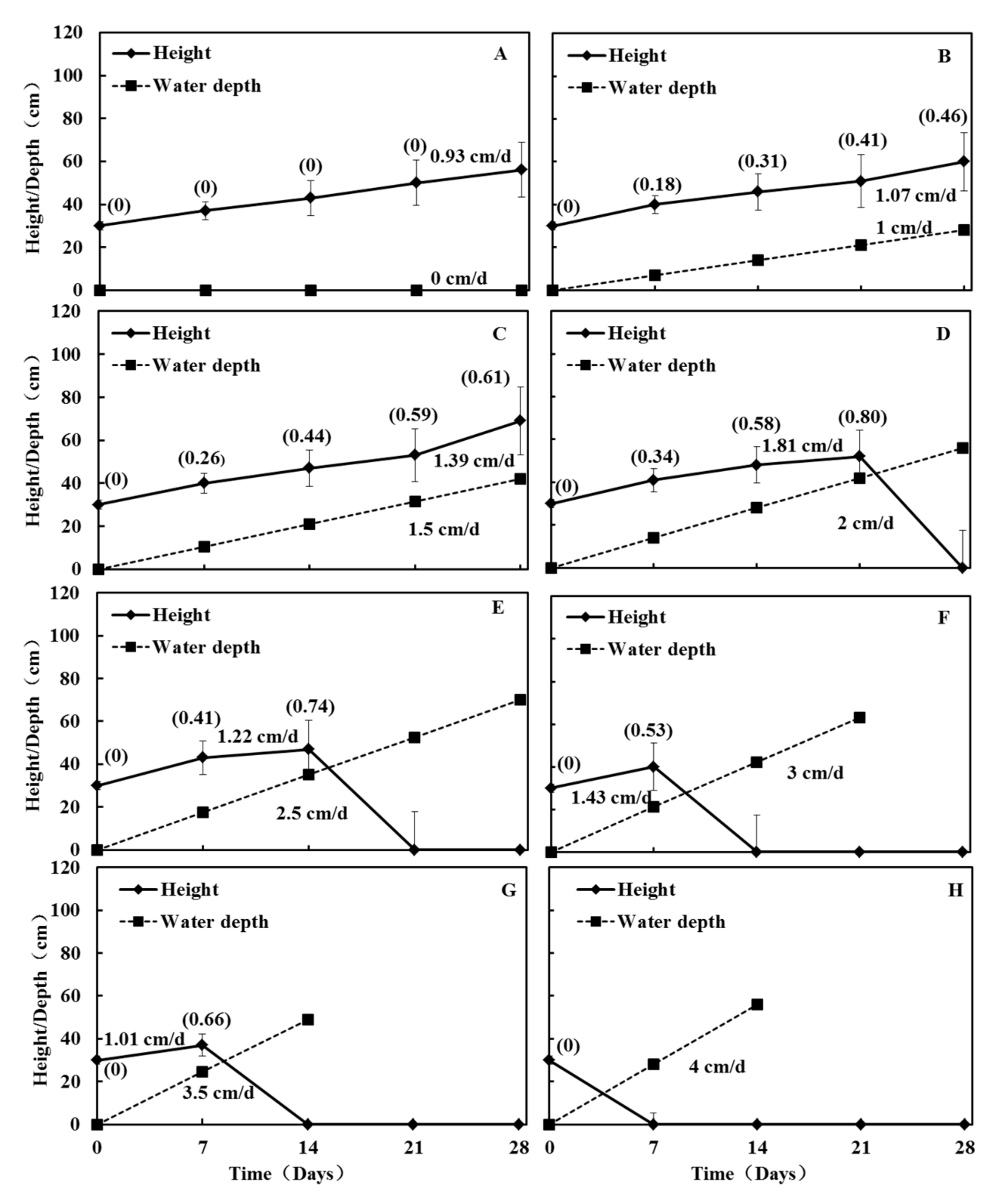
2.4.2. Experiment 2: Responses of Seedling Growth to Submergence Depth and Submergence Rate
2.5. Data Analysis
3. Results
3.1. Phenology of Growth in T. angustifolia
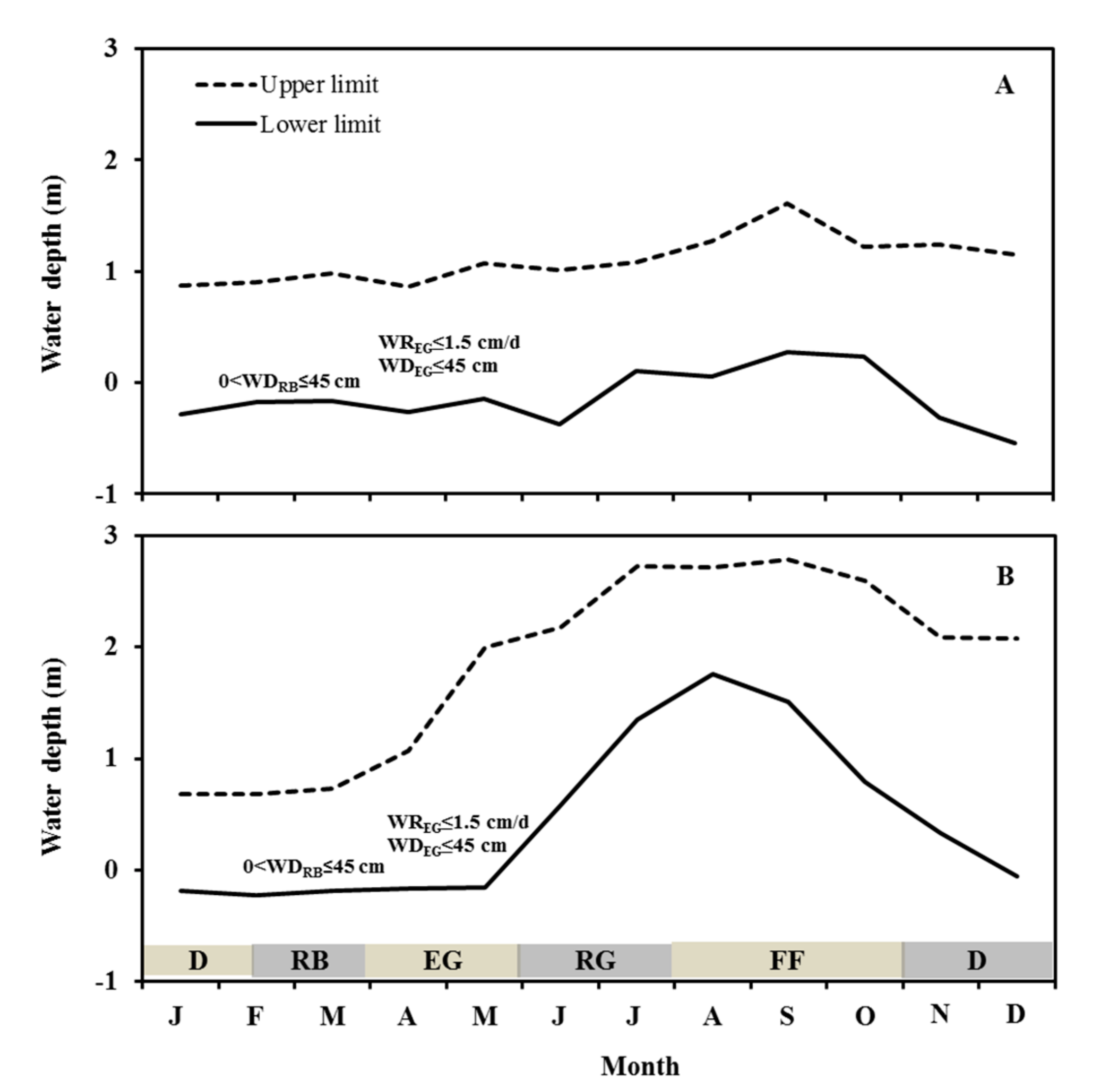
3.2. WLFs, Temperature and T. angustifolia Performance in Lakes
3.3. Responses of RBS to Submergence Depth (Experiment 1)
3.4. Responses of Seedling Growth to Submergence Depth and Submergence Rate (Experiment 2)
4. Discussion
4.1. WLFRs of Typha angustifolia
4.2. Comparison with Competitive Hygrophytes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Maberly, S.C. Ecology of Shallow Lakes. Aquat. Bot. 2000, 68, 93–94. [Google Scholar] [CrossRef]
- Poff, N.L.; Allan, J.D.; Bain, M.B.; Karr, J.R.; Prestegaard, K.L.; Richter, B.D.; Sparks, R.E.; Stromberg, J.C. The natural flow regime. BioScience 1997, 47, 769–784. [Google Scholar] [CrossRef]
- Lytle, D.A.; Poff, N.L. Adaption to natural flow regimes. Trends Ecol. Evol. 2004, 19, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Manzur, M.E.; Grimoldi, A.A.; Insausti, P.; Striker, G.G. Escape from water or remain quiescent? Lotus tenuis changes its strategy depending on depth of submergence. Ann. Bot. 2009, 104, 1163–1169. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.Z.; Wang, H.J. Ecological effects of river-lake disconnection and restoration strategies in the mid-lower Yangtze River. In Ecological Management on Water and Sediment in the Yangtze River Basin; Science Press: Beijing, China, 2009. (In Chinese) [Google Scholar]
- Zhang, X.K.; Liu, X.Q.; Wang, H.Z. Developing water level regulation strategies for macrophytes restoration of a large river-disconnected lake. Ecol. Eng. 2014, 68, 25–31. [Google Scholar] [CrossRef]
- Mahoney, J.M.; Rood, S.B. Stream flow requirements for cottonwood seedling recruitment in integrative model. Wetlands 1998, 18, 634–645. [Google Scholar] [CrossRef]
- Rood, S.B.; Samuelson, G.M.; Braatne, J.H.; Gourley, C.R.; Hughes, F.M.R.; Mahoney, J.M. Managing river flows to restore floodplain forests. Front. Ecol. Environ. 2005, 3, 193–201. [Google Scholar] [CrossRef]
- Merritt, D.M.; Scott, M.L.; Poff, N.L.; Auble, G.T.; Lytle, D.A. Theory, methods and tools for determining environmental flows for riparian vegetation: Riparian vegetation-flow response guilds. Freshw. Biol. 2010, 55, 206–225. [Google Scholar] [CrossRef]
- Scott, M.L.; Shafroth, P.B.; Auble, G.T. Responses of riparian cottonwoods to alluvial water table declines. Environ. Manag. 1999, 23, 347–358. [Google Scholar] [CrossRef]
- Cooper, D.J.; D’Amico, D.R.; Scott, M.L. Physiological and morphological response patterns of Populus deltoides to alluvial groundwater. J. Environ. Manag. 2003, 31, 215–226. [Google Scholar] [CrossRef]
- Kennedy, M.P.; Milne, J.M.; Murphy, K.J. Experimental growth responses to groundwater level variation and competition in five British wetland plant species. Wetl. Ecol. Manag. 2003, 11, 383–396. [Google Scholar] [CrossRef]
- Driver, P.D.; Barbour, E.J.; Michener, K. An integrated surface water, groundwater and wetland plant model of drought response and recovery for environmental water management. In Proceedings of the MODSIM 2011 International Congress on Modelling and Simulation 2011, Perth, Australia, 12–16 December 2011; Volume 26, pp. 2444–2450. [Google Scholar]
- Merritt, L.D.M. Hydrologic regimes and riparian forests: A structured population model for cottonwood. Ecology 2004, 85, 2493–2503. [Google Scholar]
- Smith, M.; Caswell, H.; Mettler-Cherry, P. Stochastic flood and precipitation regimes and the population dynamics of a threatened floodplain plant. Ecol. Appl. 2005, 15, 1036–1052. [Google Scholar] [CrossRef] [Green Version]
- Hebb, A.J.; Mortsch, L.D.; Deadman, P.J.; Cabrera, A.R. Modeling wetland vegetation community response to water-level change at Long Point, Ontario. J. Great Lakes Res. 2013, 39, 191–200. [Google Scholar] [CrossRef]
- Palanisamy, B.; Chui, T.F.M. Understanding wetland plant dynamics in response to water table changes through ecohydrological modelling. Ecohydrology 2013, 6, 287–296. [Google Scholar] [CrossRef]
- Edwards, A.L.; Lee, D.W.; Richards, J.H. Responses to a fluctuating environment: Effects of water depth on growth and biomass allocation in Eleocharis cellulosa Torr. (Cyperaceae). Can. J. Bot. 2003, 81, 964–975. [Google Scholar] [CrossRef]
- Deegan, B.M.; White, S.D.; Ganf, G.G. The influence of water level fluctuations on the growth of four emergent macrophyte species. Aquat. Bot. 2007, 86, 309–315. [Google Scholar] [CrossRef]
- Wu, X.D.; Wang, G.X.; Wei, H.N.; Li, Z.G.; Hang, Z.Q. Growth responses of Hydrilla verticillata to increasing water levels. J. Lake Sci. 2012, 24, 384–390. (In Chinese) [Google Scholar]
- Bai, X.; Chen, K.; Chen, X. Short-time response in growth and sediment properties of Zizania latifolia to water depth. Environ. Earth Sci. 2013, 70, 2847–2854. [Google Scholar] [CrossRef]
- Wang, Q.; Chen, J.; Liu, F.; Li, W. Morphological changes and resource allocation of Zizania latifolia (Griseb.) Stapf in response to different submergence depth and duration. Flora 2014, 209, 279–284. [Google Scholar] [CrossRef]
- Wang, H.L.; Zhang, X.K.; Wan, A. Morphological responses of Zizania latifolia seedlings at different ages to short-term submergence. J. Lake Sci. 2018, 30, 192–198. (In Chinese) [Google Scholar]
- Wang, H.Z.; Wang, H.J.; Liang, X.M.; Ni, L.Y.; Liu, X.Q.; Cui, Y.D. Empirical modelling of submerged macrophytes in Yangtze lakes. Ecol. Model. 2005, 188, 483–491. [Google Scholar] [CrossRef]
- Yuan, S.B.; Yang, Z.D.; Liu, X.Q.; Wang, H.Z. Key parameters of water level fluctuations determining the distribution of Carex in shallow lakes. Wetlands 2017, 6, 1005–1014. [Google Scholar] [CrossRef]
- Liu, X.; Yang, Z.D.; Yuan, S.B.; Wang, H.Z. A novel methodology for the assessment of water level requirements in shallow lakes. Ecol. Eng. 2017, 102, 31–38. [Google Scholar] [CrossRef]
- Yuan, S.B.; Yang, Z.D.; Liu, X.Q.; Wang, H.Z. Water level requirements of a Carex hygrophyte in Yangtze floodplain lakes. Ecol. Eng. 2019, 129, 29–37. [Google Scholar] [CrossRef]
- Yuan, S.B. Environmental Flow Requirements of Hygrophytic Vegetation in the Yangtze Floodplain. Ph.D. Thesis, University of Chinese Academy of Sciences, Beijing, China, 2019. (In Chinese). [Google Scholar]
- Weisner, S.E.B. Within lake patterns in depth penetration of emergent vegetation. Freshw. Biol. 1991, 26, 133–142. [Google Scholar] [CrossRef]
- Sharma, P.; Asaeda, T.; Fujino, T. Effect of water depth on the rhizome dynamics of Typha angustifolia. Wetl. Ecol. Manag. 2008, 16, 43–49. [Google Scholar] [CrossRef]
- Sharma, P.; Asaeda, T.; Kalibbala, M.; Fujino, T. Morphology, growth and carbohydrate storage of the plant Typha angustifolia at different water depths. Chem. Ecol. 2008, 24, 133–145. [Google Scholar] [CrossRef]
- Asaeda, T.; Fujino, T.; Manatunge, J. Morphological adaptations of emergent plants to water flow: A case study with Typha angustifolia, Zizania latifolia and Phragmites australis. Freshw. Biol. 2010, 50, 1991–2001. [Google Scholar] [CrossRef]
- Wang, Q.L.; Chen, J.R.; Liu, H. The growth responses of two emergent plants to the water depth. Acta Hydrobiol. Sin. 2012, 36, 583–587. (In Chinese) [Google Scholar]
- Grace, J.B.; Wetzel, R.G. Habitat partitioning and competitive displacement in Typha: Experimental field studies. Am. Nat. 1981, 118, 463–474. [Google Scholar] [CrossRef]
- Zhang, X.K. Water Level Fluctuation Requirements of Plants in the Yangtze Floodplain Lakes. Ph.D. Thesis, University of Chinese Academy of Sciences, Beijing, China, 2013. (In Chinese). [Google Scholar]
- Inoue, T.; Tsuchiya, T. Depth distribution of three Typha species, Typha orientalis Presl, Typha angustifolia L. and Typha latifolia L. in an artificial pond. Plant. Species Biol. 2010, 24, 47–52. [Google Scholar] [CrossRef]
- Squires, L. Water-depth tolerances of the dominant emergent macrophytes of the Delta Marsh, Manitoba. Can. J. Bot. 1992, 70, 1860–1867. [Google Scholar] [CrossRef]
- Boers, A.M.; Zedler, J.B. Stabilized water levels and Typha invasiveness. Wetlands 2008, 28, 676–685. [Google Scholar] [CrossRef]
- Wang, H.Z.; Liu, X.Q.; Wang, H.J. The Yangtze River Floodplain: Threats and Rehabilitation. Am. Fish. Soc. Symp. 2016, 84, 263–291. [Google Scholar]
- Li, C.M.; Ye, X.Q.; Wu, M.; Shao, X.X. Effect of water depth and co-existence on the growth characteristics of Phragmites communis and Typha angustifolia. Wetl. Sci. 2015, 13, 609–615. (In Chinese) [Google Scholar]
- Tang, S.W.; Cao, J.; Xu, L.M. Simulation Experiment on Adaptation of Typha orientalis to Flooded Habitat. Wetl. Sci. 2019, 17, 582–592. (In Chinese) [Google Scholar]
- Chen, H.; Zamorano, M.F.; Ivanoff, D. Effect of flooding depth on growth, biomass, photosynthesis, and chlorophyll fluorescence of Typha domingensis. Wetlands 2010, 30, 957–965. [Google Scholar] [CrossRef]
- Li, W. Studies on Aquatic Vegetation and its Succession in Honghu Lake. Ph.D. Thesis, Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan, China, 1995. (In Chinese). [Google Scholar]
- Maricle, B.R.; Lee, R.W. Aerenchyma development and oxygen transport in the estuarine cordgrasses Spartina alterniflora, and S. anglica. Aquat. Bot. 2002, 74, 109–120. [Google Scholar] [CrossRef]
- Voesenek, L.A.C.J.; Benschop, J.J.; Bou, J.; Cox, M.C.H.; Groeneveld, H.W.; Millenaar, F.F.; Vreeburg, R.A.M.; Peeters, A.J.M. Interactions between plant hormones regulate submergence-induced shoot elongation in the flooding-tolerant dicot Rumex palustris. Ann. Bot. 2003, 91, 205–211. [Google Scholar] [CrossRef] [Green Version]
- Colmer, T.D.; Voesenek, L.A.C.J. Flooding tolerance: Suites of plant traits in variable environments. Funct. Plant Biol. 2009, 36, 665–681. [Google Scholar] [CrossRef]
- Chen, X.S.; Deng, Z.M.; Xie, Y.H.; Li, F.; Li, X. Differential growth and vegetative reproduction by two co-occurring emergent macrophytes along a water table gradient. Pak. J. Bot. 2014, 46, 881–886. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | January | February | March | April | May | June | July | |
| Height | r | −0.324 ** | −0.322 ** | −0.302 ** | −0.328 ** | −0.345 ** | 0.763 ** | 0.654 ** |
| P | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | |
| Biomass | r | −0.395 ** | −0.398 ** | −0.461 ** | −0.444 ** | −0.548 ** | 0.226 * | 0.124 |
| P | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | 0.017 | 0.184 | |
| August | September | October | November | December | Amplitude | |||
| Height | r | 0.541 ** | 0.465 ** | 0.462 ** | 0.298 ** | −0.350 ** | 0.765 ** | |
| P | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | ||
| Biomass | r | 0.044 | 0.057 | 0.043 | 0.018 | −0.07 | −0.389 ** | |
| P | 0.653 | 0.542 | 0.634 | 0.76 | 0.48 | <0.01 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Z.-D.; Yuan, S.-B.; Liu, X.-Q.; Wang, H.-Z. Water Level Fluctuation Requirements of Emergent Macrophyte Typha angustifolia L. Water 2020, 12, 127. https://doi.org/10.3390/w12010127
Yang Z-D, Yuan S-B, Liu X-Q, Wang H-Z. Water Level Fluctuation Requirements of Emergent Macrophyte Typha angustifolia L. Water. 2020; 12(1):127. https://doi.org/10.3390/w12010127
Chicago/Turabian StyleYang, Zhen-Dong, Sai-Bo Yuan, Xue-Qin Liu, and Hong-Zhu Wang. 2020. "Water Level Fluctuation Requirements of Emergent Macrophyte Typha angustifolia L." Water 12, no. 1: 127. https://doi.org/10.3390/w12010127
APA StyleYang, Z. -D., Yuan, S. -B., Liu, X. -Q., & Wang, H. -Z. (2020). Water Level Fluctuation Requirements of Emergent Macrophyte Typha angustifolia L. Water, 12(1), 127. https://doi.org/10.3390/w12010127